
Projected Changes in Spawning Ground Distribution of Mature Albacore Tuna in the Indian Ocean under Various Global Climate Change Scenarios


Abstract
:1. Introduction
2. Materials and Methods
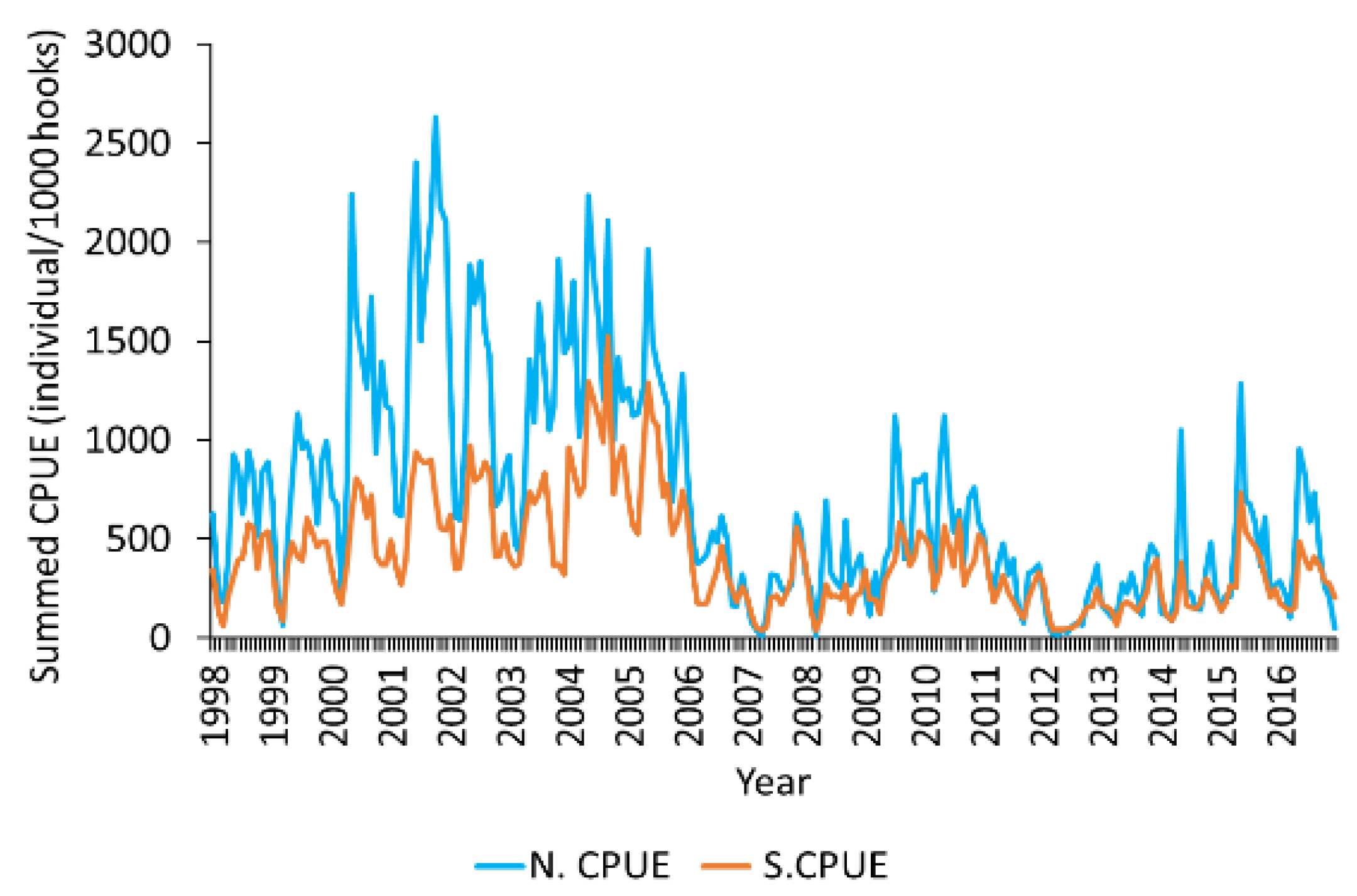
2.1. Albacore Tuna Fishery Data
2.2. Oceanographic Data
2.3. Projected Oceanographic Data
2.4. Standardisation of Nominal Catch per Unit Effort
2.5. Species Distribution Modelling
2.5.1. Construction of Suitability Index Curves
2.5.2. Model Construction
2.6. Model Selection and Prediction
3. Results
3.1. Standardisation of N.CPUE Data
3.2. Construction of SI Curves
3.3. Analysis of Habitat Models, Model Selection, and Validation
3.4. Validation of Selected Models
3.5. HSI Prediction
3.6. Projected Habitat Changes
4. Discussion
4.1. Projected Habitat Changes under Various Climate Change Scenarios
4.2. Increased Water Temperature and Albacore Spawning
4.3. Temperature and Other Oceanographic Parameters
4.4. Sustainable Development Goals
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nikolic, N.; Morandeau, G.; Hoarau, L.; West, W.; Arrizabalaga, H.; Hoyle, S.; Nicol, S.J.; Bourjea, J.; Puech, A.; Farley, J.H.; et al. Review of albacore tuna, Thunnus alalunga, biology, fisheries and management. Rev. Fish Biol. Fish. 2017, 27, 775–810. [Google Scholar] [CrossRef]
- Guillotreau, P.; Squires, D.; Sun, J.; Compeán, G.A. Local, regional and global markets: What drives the tuna fisheries? Rev. Fish Biol. Fish. 2017, 27, 909–929. [Google Scholar] [CrossRef]
- Mondal, S.; Vayghan, A.H.; Lee, M.A.; Wang, Y.C.; Semedi, B. Habitat suitability modeling for the feeding ground of immature albacore in the southern Indian Ocean using satellite-derived sea surface temperature and chlorophyll data. Remote Sens. 2021, 13, 2669. [Google Scholar] [CrossRef]
- Chen, I.C.; Lee, P.F.; Tzeng, W.N. Distribution of albacore (Thunnus alalunga) in the Indian Ocean and its relation to environmental factors. Fish. Oceanogr. 2005, 14, 71–80. [Google Scholar] [CrossRef]
- Hereher, M.E. Assessment of climate change impacts on sea surface temperatures and sea level rise—The Arabian Gulf. Climate 2020, 8, 50. [Google Scholar] [CrossRef]
- Ruela, R.; Sousa, M.C.; de Castro, M.; Dias, J.M. Global and regional evolution of sea surface temperature under climate change. Glob. Planet. Chang. 2020, 190, 103190. [Google Scholar] [CrossRef]
- Wiryawan, B.; Loneragan, N.; Mardhiah, U.; Kleinertz, S.; Wahyuningrum, P.I.; Pingkan, J.; Wildan; Timur, P.S.; Duggan, D.; Yulianto, I. Catch per unit effort dynamic of yellowfin tuna related to sea surface temperature and chlorophyll in Southern Indonesia. Fishes 2020, 5, 28. [Google Scholar] [CrossRef]
- Jansen, T.; Gislason, H. Temperature affects the timing of spawning and migration of North Sea mackerel. Cont. Shelf Res. 2011, 31, 64–72. [Google Scholar] [CrossRef]
- Kramer, D.L. Dissolved oxygen and fish behavior. Environ. Biol. Fishes 1987, 18, 81–92. [Google Scholar] [CrossRef]
- Sear, D.A.; Pattison, I.; Collins, A.L.; Newson, M.D.; Jones, J.I.; Naden, P.S.; Carling, P.A. Factors controlling the temporal variability in dissolved oxygen regime of salmon spawning gravels. Hydrol. Process. 2014, 28, 86–103. [Google Scholar] [CrossRef]
- Urbina, M.A.; Glover, C.N. Effect of salinity on osmoregulation, metabolism and nitrogen excretion in the amphidromous fish, inanga (Galaxias maculatus). J. Exp. Mar. Biol. Ecol. 2015, 473, 7–15. [Google Scholar] [CrossRef]
- Eddy, F.B.; Chang, Y.J. Effects of salinity in relation to migration and development in fish. In The Vertebrate Gas Transport Cascade: Adaptations to Environment and Mode of Life; CRC Press, Inc.: Boca Raton, FL, USA, 1993; pp. 35–42. [Google Scholar]
- Ferreira-Martins, D.; Coimbra, J.; Antunes, C.; Wilson, J.M. Effects of salinity on upstream-migrating, spawning sea lamprey, Petromyzon marinus. Conserv. Physiol. 2016, 4, cov064. [Google Scholar] [CrossRef] [PubMed]
- Reinoso, S.; Mora-Pinargote, J.; Bohórquez-Cruz, M.; Sonneholzner, S.; Argüello-Guevara, W. Effect of water salinity on embryonic development of longfin yellowtail Seriola rivoliana larvae. Aquac. Res. 2020, 51, 1317–1321. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.; Liu, L. Synchronous Variations in Abundance and Distribution of Ommastrephes bartramii and Dosidicus gigas in the Pacific Ocean. J. Ocean. Univ. China 2021, 20, 695–705. [Google Scholar] [CrossRef]
- Fournier, S.; Lee, T.; Wang, X.; Armitage, T.W.; Wang, O.; Fukumori, I.; Kwok, R. Sea surface salinity as a proxy for Arctic Ocean freshwater changes. J. Geophys. Res. Ocean. 2020, 125, e2020JC016110. [Google Scholar] [CrossRef]
- Asch, R.G.; Checkley, D.M., Jr. Dynamic height: A key variable for identifying the spawning habitat of small pelagic fishes. Deep Sea Res. Part I Oceanogr. Res. Pap. 2013, 71, 79–91. [Google Scholar] [CrossRef]
- Ma, T.; Wu, G.; Liu, Y.; Mao, J. Abnormal warm sea-surface temperature in the Indian Ocean, active potential vorticity over the Tibetan Plateau, and severe flooding along the Yangtze River in summer 2020. Q. J. R. Meteorol. Soc. 2022, 148, 1001–1019. [Google Scholar] [CrossRef]
- Garcia-Soto, C.; Cheng, L.; Caesar, L.; Schmidtko, S.; Jewett, E.B.; Cheripka, A.; Rigor, I.; Caballero, A.; Chiba, S.; Báez, J.C.; et al. An overview of ocean climate change indicators: Sea surface temperature, ocean heat content, ocean pH, dissolved oxygen concentration, Arctic Sea ice extent, thickness and volume, sea level and strength of the AMOC (Atlantic Meridional Overturning Circulation). Front. Mar. Sci. 2021, 8, 642372. [Google Scholar]
- Dinnat, E.P.; Le Vine, D.M.; Boutin, J.; Meissner, T.; Lagerloef, G. Remote sensing of sea surface salinity: Comparison of satellite and in situ observations and impact of retrieval parameters. Remote Sens. 2019, 11, 750. [Google Scholar] [CrossRef]
- Wells, B.K.; Grimes, C.B.; Waldvogel, J.B. Quantifying the effects of wind, upwelling, curl, sea surface temperature and sea level height on growth and maturation of a California Chinook salmon (Oncorhynchus tshawytscha) population. Fish. Oceanogr. 2007, 16, 363–382. [Google Scholar] [CrossRef]
- Makwinja, R.; Mengistou, S.; Kaunda, E.; Alamirew, T. Lake Malombe fish stock fluctuation: Ecosystem and fisherfolks. Egypt. J. Aquat. Res. 2021, 47, 321–327. [Google Scholar] [CrossRef]
- Michael-Bitton, G.; Gal, G.; Corrales, X.; Ofir, E.; Shechter, M.; Zemah-Shamir, S. Economic aspects of fish stock accounting as a renewable marine natural capital: The Eastern Mediterranean continental shelf ecosystem as a case study. Ecol. Econ. 2022, 200, 107539. [Google Scholar] [CrossRef]
- Alqattan, M.E.; Gray, T.S. Marine Pollution in Kuwait and Its Impacts on Fish-Stock Decline in Kuwaiti Waters: Reviewing the Kuwaiti Government’s Policies and Practices. Front. Sustain. 2021, 2, 667822. [Google Scholar] [CrossRef]
- Khoshnood, Z. Effects of environmental pollution on fish: A short review. Transylv. Rev. Syst. Ecol. Res. 2017, 19, 49–60. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Lønborg, C.; Carreira, C.; Jickells, T.; Álvarez-Salgado, X.A. Impacts of global change on ocean dissolved organic carbon (DOC) cycling. Front. Mar. Sci. 2020, 7, 466. [Google Scholar] [CrossRef]
- Arneth, A.; Shin, Y.J.; Leadley, P.; Rondinini, C.; Bukvareva, E.; Kolb, M.; Midgley, G.F.; Oberdorff, T.; Palomo, I.; Saito, O. Post-2020 biodiversity targets need to embrace climate change. Proc. Natl. Acad. Sci. USA 2020, 117, 30882–30891. [Google Scholar] [CrossRef]
- Suh, S.; Johnson, J.A.; Tambjerg, L.; Sim, S.; Broeckx-Smith, S.; Reyes, W.; Chaplin-Kramer, R. Closing yield gap is crucial to avoid potential surge in global carbon emissions. Glob. Environ. Chang. 2020, 63, 102100. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pörtner, H.O.; Roberts, D.; Skea, J.; Shukla, P.R. Global Warming of 1.5 C: IPCC Special Report on Impacts of Global Warming of 1.5 C above Pre-Industrial Levels in Context of Strengthening Response to Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Kroeker, K.J.; Sanford, E. Ecological leverage points: Species interactions amplify the physiological effects of global environmental change in the ocean. Annu. Rev. Mar. Sci. 2022, 14, 75–103. [Google Scholar] [CrossRef]
- Hu, W.; Du, J.; Su, S.; Tan, H.; Yang, W.; Ding, L.; Dong, P.; Yu, W.; Zheng, X.; Chen, B. Effects of climate change in the seas of China: Predicted changes in the distribution of fish species and diversity. Ecol. Indic. 2022, 134, 108489. [Google Scholar] [CrossRef]
- Nataniel, A.; Pennino, M.G.; Lopez, J.; Soto, M. Modelling the impacts of climate change on skipjack tuna (Katsuwonus pelamis) in the Mozambique Channel. Fish. Oceanogr. 2022, 31, 149–163. [Google Scholar] [CrossRef]
- Nicol, S.; Lehodey, P.; Senina, I.; Bromhead, D.; Frommel, A.Y.; Hampton, J.; Havenhand, J.; Margulies, D.; Munday, P.L.; Scholey, V.; et al. Ocean futures for the world’s largest yellowfin tuna population under the combined effects of ocean warming and acidification. Front. Mar. Sci. 2022, 9, 816772. [Google Scholar] [CrossRef]
- Olafsdottir, A.H.; Utne, K.R.; Jacobsen, J.A.; Jansen, T.; Óskarsson, G.J.; Nøttestad, L.; Elvarsson, B.; Broms, C.; Slotte, A. Geographical expansion of Northeast Atlantic mackerel (Scomber scombrus) in the Nordic Seas from 2007 to 2016 was primarily driven by stock size and constrained by low temperatures. Deep Sea Res. Part II Top. Stud. Oceanogr. 2019, 159, 152–168. [Google Scholar] [CrossRef]
- Rutterford, L.A.; Simpson, S.D.; Bogstad, B.; Devine, J.A.; Genner, M.J. Sea temperature is the primary driver of recent and predicted fish community structure across Northeast Atlantic shelf seas. Glob. Chang. Biol. 2023, 29, 2510–2521. [Google Scholar] [CrossRef] [PubMed]
- Issifu, I.; Alava, J.J.; Lam, V.W.; Sumaila, U.R. Impact of ocean warming, overfishing and mercury on European fisheries: A risk assessment and policy solution framework. Front. Mar. Sci. 2022, 8, 770805. [Google Scholar] [CrossRef]
- Zhu, B.R.; Verhoeven, M.A.; Velasco, N.; Sanchez-Aguilar, L.; Zhang, Z.; Piersma, T. Current breeding distributions and predicted range shifts under climate change in two subspecies of Black-tailed Godwits in Asia. Glob. Chang. Biol. 2022, 28, 5416–5426. [Google Scholar] [CrossRef]
- Cuilleret, M.; Doyen, L.; Gomes, H.; Blanchard, F. Resilience management for coastal fisheries facing with global changes and uncertainties. Econ. Anal. Policy 2022, 74, 634–656. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, J.; Krebs, P. Low impact development practices mitigate urban flooding and non-point pollution under climate change. J. Clean. Prod. 2022, 347, 131320. [Google Scholar] [CrossRef]
- Pilcher, D.J.; Cross, J.N.; Hermann, A.J.; Kearney, K.A.; Cheng, W.; Mathis, J.T. Dynamically downscaled projections of ocean acidification for the Bering Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2022, 198, 105055. [Google Scholar] [CrossRef]
- Vayghan, A.H.; Poorbagher, H.; Shahraiyni, H.T.; Fazli, H.; Saravi, H.N. Suitability indices and habitat suitability index model of Caspian kutum (Rutilus frisii kutum) in the southern Caspian Sea. Aquat. Ecol. 2013, 47, 441–451. [Google Scholar] [CrossRef]
- Lee, M.A.; Weng, J.S.; Lan, K.W.; Vayghan, A.H.; Wang, Y.C.; Chan, J.W. Empirical habitat suitability model for immature albacore tuna in the North Pacific Ocean obtained using multisatellite remote sensing data. Int. J. Remote Sens. 2020, 41, 5819–5837. [Google Scholar] [CrossRef]
- Chen, X.; Li, G.; Feng, B.; Tian, S. Habitat suitability index of Chub mackerel (Scomber japonicus) from July to September in the East China Sea. J. Oceanogr. 2009, 65, 93–102. [Google Scholar] [CrossRef]
- Lee, D.; Son, S.; Kim, W.; Park, J.M.; Joo, H.; Lee, S.H. Spatio-temporal variability of the habitat suitability index for Chub Mackerel (Scomber japonicus) in the East/Japan Sea and the South sea of South Korea. Remote Sens. 2018, 10, 938. [Google Scholar] [CrossRef]
- Lee, D.; Son, S.H.; Lee, C.I.; Kang, C.K.; Lee, S.H. Spatio-temporal variability of the habitat suitability index for the Todarodes pacificus (Japanese common squid) around South Korea. Remote Sens. 2019, 11, 2720. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Brown, C.J.; Sydeman, W.J.; Kiessling, W.; Schoeman, D.S.; Moore, P.J.; Brander, K.; Bruno, J.F.; Buckley, L.B.; Burrows, M.T.; et al. Global imprint of climate change on marine life. Nat. Clim. Chang. 2013, 3, 919–925. [Google Scholar] [CrossRef]
- Chang, Y.J.; Hsu, J.; Lai, P.K.; Lan, K.W.; Tsai, W.P. Evaluation of the impacts of climate change on albacore distribution in the South Pacific Ocean by using ensemble forecast. Front. Mar. Sci. 2021, 8, 731950. [Google Scholar] [CrossRef]
- Christian, J.R.; Holmes, J. Changes in albacore tuna habitat in the northeast Pacific Ocean under anthropogenic warming. Fish. Oceanogr. 2016, 25, 544–554. [Google Scholar] [CrossRef]
- Dell’Apa, A.; Carney, K.; Davenport, T.M.; Carle, M.V. Potential medium-term impacts of climate change on tuna and billfish in the Gulf of Mexico: A qualitative framework for management and conservation. Mar. Environ. Res. 2018, 141, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.C.; Cheung, W.W. Using fuzzy logic to determine the vulnerability of marine species to climate change. Glob. Chang. Biol. 2018, 24, e719–e731. [Google Scholar] [CrossRef]
- Beaugrand, G.; Kirby, R.R. How do marine pelagic species respond to climate change? Theories and observations. Annu. Rev. Mar. Sci. 2018, 10, 169–197. [Google Scholar] [CrossRef]
- Oschlies, A. Ocean Deoxygenation from Climate Change; IUCN: Gland, Switzerland, 2019. [Google Scholar]
- Bromhead, D.; Scholey, V.; Nicol, S.; Margulies, D.; Wexler, J.; Stein, M.; Hoyle, S.; Lennert-Cody, C.; Williamson, J.; Havenhand, J.; et al. The potential impact of ocean acidification upon eggs and larvae of yellowfin tuna (Thunnus albacares). Deep. Sea Res. Part II Top. Stud. Oceanogr. 2015, 113, 268–279. [Google Scholar] [CrossRef]
- Olson, S.; Jansen, M.F.; Abbot, D.S.; Halevy, I.; Goldblatt, C. The effect of ocean salinity on climate and its implications for Earth’s habitability. Geophys. Res. Lett. 2022, 49, e2021GL095748. [Google Scholar] [CrossRef]
- Sagarminaga, Y.; Arrizabalaga, H. Relationship of Northeast Atlantic albacore juveniles with surface thermal and chlorophyll-a fronts. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2014, 107, 54–63. [Google Scholar] [CrossRef]
- Arrizabalaga, H.; Dufour, F.; Kell, L.; Merino, G.; Ibaibarriaga, L.; Chust, G.; Irigoien, X.; Santiago, J.; Murua, H.; Fraile, I.; et al. Global habitat preferences of commercially valuable tuna. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2015, 113, 102–112. [Google Scholar] [CrossRef]
- Rowden, A.A.; Anderson, O.F.; Georgian, S.E.; Bowden, D.A.; Clark, M.R.; Pallentin, A.; Miller, A. High-resolution habitat suitability models for the conservation and management of vulnerable marine ecosystems on the Louisville Seamount Chain, South Pacific Ocean. Front. Mar. Sci. 2017, 4, 335. [Google Scholar] [CrossRef]
- Mugagga, F.; Nabaasa, B.B. The centrality of water resources to the realization of Sustainable Development Goals (SDG). A review of potentials and constraints on the African continent. Int. Soil Water Conserv. Res. 2016, 4, 215–223. [Google Scholar] [CrossRef]
- Ntona, M.; Morgera, E. Connecting SDG 14 with the other Sustainable Development Goals through marine spatial planning. Mar. Policy 2018, 93, 214–222. [Google Scholar] [CrossRef]
- Virto, L.R. A preliminary assessment of the indicators for Sustainable Development Goal (SDG) 14 “Conserve and sustainably use the oceans, seas and marine resources for sustainable development”. Mar. Policy 2018, 98, 47–57. [Google Scholar] [CrossRef]
- Kenny, A.J.; Campbell, N.; Koen-Alonso, M.; Pepin, P.; Diz, D. Delivering sustainable fisheries through adoption of a risk-based framework as part of an ecosystem approach to fisheries management. Mar. Policy 2018, 93, 232–240. [Google Scholar] [CrossRef]
- Cormier, R.; Elliott, M. SMART marine goals, targets and management—Is SDG 14 operational or aspirational, is ‘Life Below Water’ sinking or swimming? Mar. Pollut. Bull. 2017, 123, 28–33. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Data | Abb. | Unit | Source | Time Period | Spatial Resolution | Temporal Resolution |
|---|---|---|---|---|---|---|
| Sea surface temperature | SST | °C | COP | 1998–2016 | 0.08° × 0.08° | Monthly |
| Dissolved oxygen | OXY | mL/L | ||||
| Sea surface salinity | SSS | psu | ||||
| Sea surface height | SSH | m |
| Institute | Code | Resolution |
|---|---|---|
| Institute Pierre Simon Laplace | IPSL | 1° × 1° |
| Geophysical Fluid Dynamics Laboratory | GFDL | 0.3–1° × 1° |
| Commonwealth Scientific and Industrial Research Organization | CSIRO | 1.5° × 1° |
| Hadley Center Global Environment Model | HadGEM | 0.3–1° × 1° |
| Max Plank Institute for Meteorology | MPI | 1° × 1° |
| Canadian Earth System Model | CanSEM | 1.5° × 1° |
| No. | Model | AMM | GMM | ||||||
|---|---|---|---|---|---|---|---|---|---|
| a | b | AIC | Adj. R2 | a | b | AIC | Adj. R2 | ||
| 1 | OXY, SST | 0.535 | 4.995 | 37.015 | 0.507 | 1.854 | 3.692 | 25.753 | 0.693 |
| 2 | OXY, SSS | −0.661 | 5.721 | 14.240 | 0.845 | 1.494 | 3.423 | 24.822 | 0.679 |
| 3 | OXY, SSH | −0.391 | 5.27 | 22.646 | 0.836 | 2.006 | 2.619 | 26.393 | 0.501 |
| 4 | SST, SSS | 2.595 | −2.129 | 26.758 | 0.375 | 1.144 | −0.77 | 26.186 | 0.115 |
| 5 | SST, SSH | 1.788 | −0.456 | 31.624 | 0.1 | 0.85 | 0.882 | 25.864 | 0.022 |
| 6 | SSS, SSH | −0.23 | 3.096 | 26.450 | 0.59 | −0.31 | 3.379 | 23.907 | 0.694 |
| 7 | OXY, SST, SSS | −0.896 | 5.623 | 33.347 | 0.71 | 1.258 | 4.459 | 18.943 | 0.87 |
| 8 | OXY, SST, SSH | 0.801 | 3.272 | 40.487 | 0.239 | 1.464 | 4.074 | 24.942 | 0.751 |
| 9 | SST, SSS, SSH | 2.117 | −0.361 | 38.593 | 0.117 | 0.339 | 2.273 | 29.604 | 0.333 |
| 10 | OXY, SST, SSS, SSH | 0.725 | 3.25 | 41.974 | 0.2 | 1.06 | 4.5 | 20.545 | 0.853 |
| Techniques | 70% | 30% | ||
|---|---|---|---|---|
| R2 | AUC | R2 | AUC | |
| Random split | 0.845 | 0.872 | 0.839 | 0.865 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mondal, S.; Ray, A.; Lee, M.-A.; Boas, M. Projected Changes in Spawning Ground Distribution of Mature Albacore Tuna in the Indian Ocean under Various Global Climate Change Scenarios. J. Mar. Sci. Eng. 2023, 11, 1565. https://doi.org/10.3390/jmse11081565
Mondal S, Ray A, Lee M-A, Boas M. Projected Changes in Spawning Ground Distribution of Mature Albacore Tuna in the Indian Ocean under Various Global Climate Change Scenarios. Journal of Marine Science and Engineering. 2023; 11(8):1565. https://doi.org/10.3390/jmse11081565
Chicago/Turabian StyleMondal, Sandipan, Aratrika Ray, Ming-An Lee, and Malagat Boas. 2023. "Projected Changes in Spawning Ground Distribution of Mature Albacore Tuna in the Indian Ocean under Various Global Climate Change Scenarios" Journal of Marine Science and Engineering 11, no. 8: 1565. https://doi.org/10.3390/jmse11081565