
Diet and Temperature Effects on the Survival of Larval Red Deep-Sea Crabs, Chaceon quinquedens (Smith, 1879), under Laboratory Conditions

Abstract
:1. Introduction
2. Materials and Methods
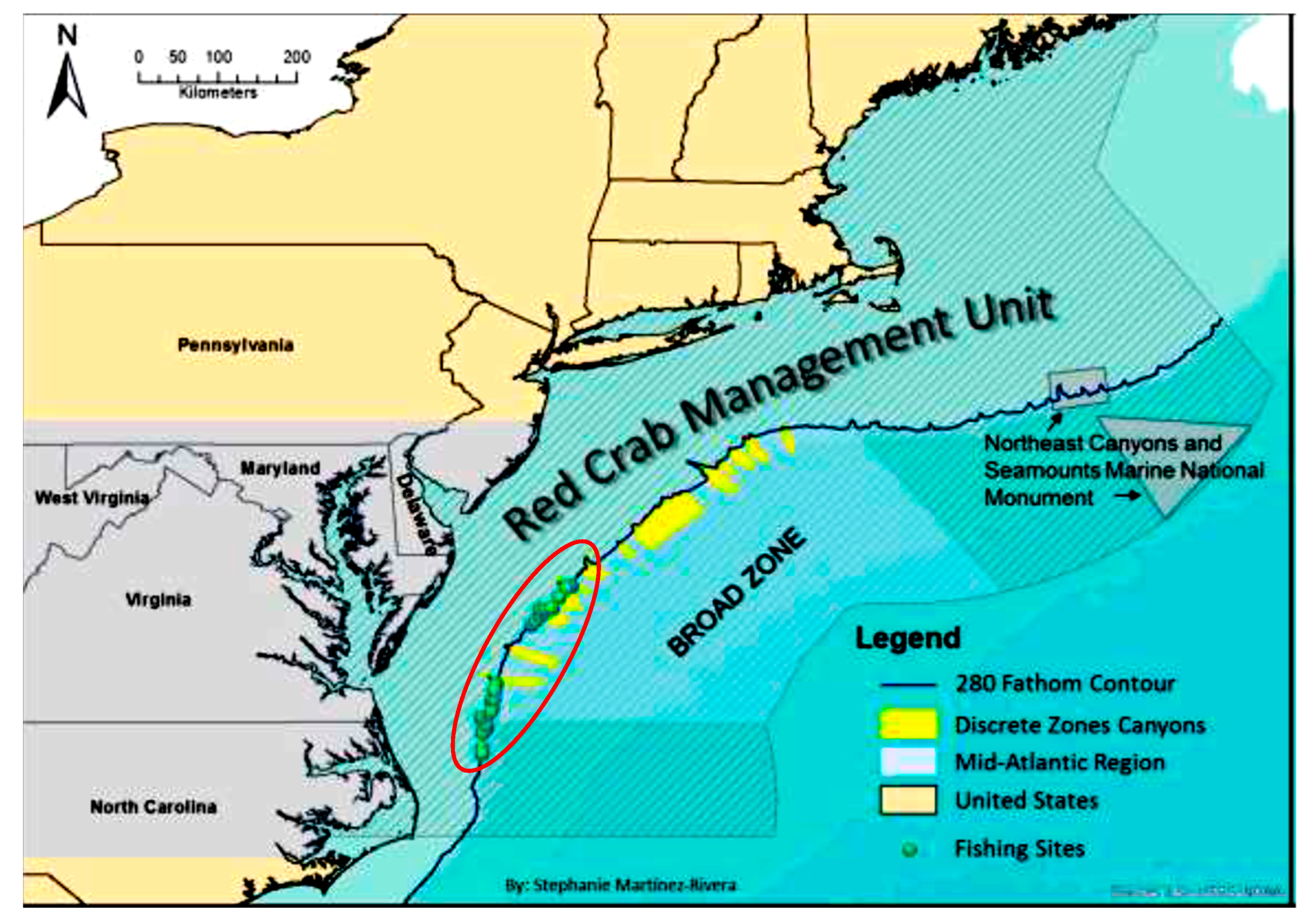
2.1. Crab Collection and Husbandry
2.2. Experimental Rearing System Setups
2.3. Larval Crab Feeding Experiments
2.4. Statistical Analysis
3. Results
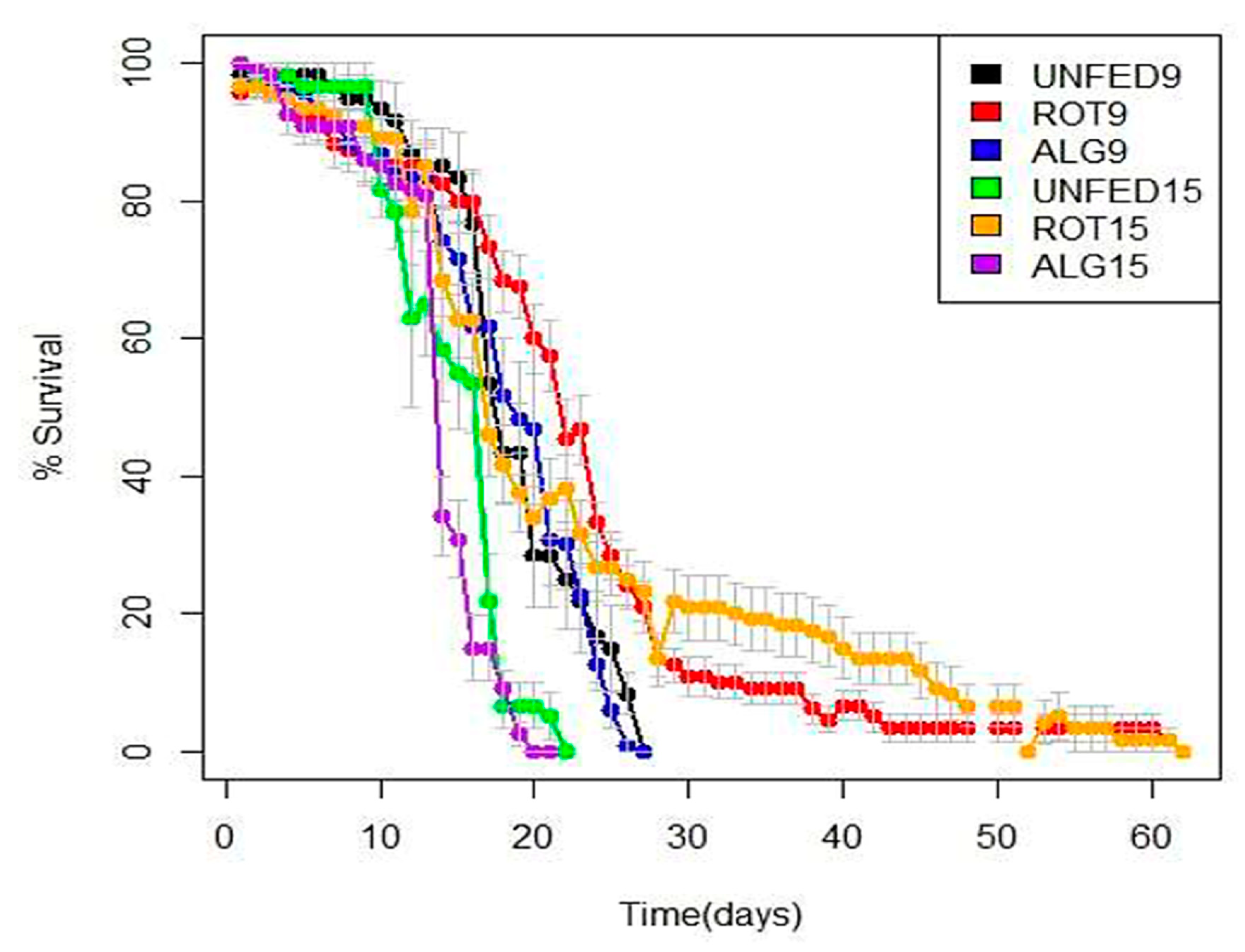
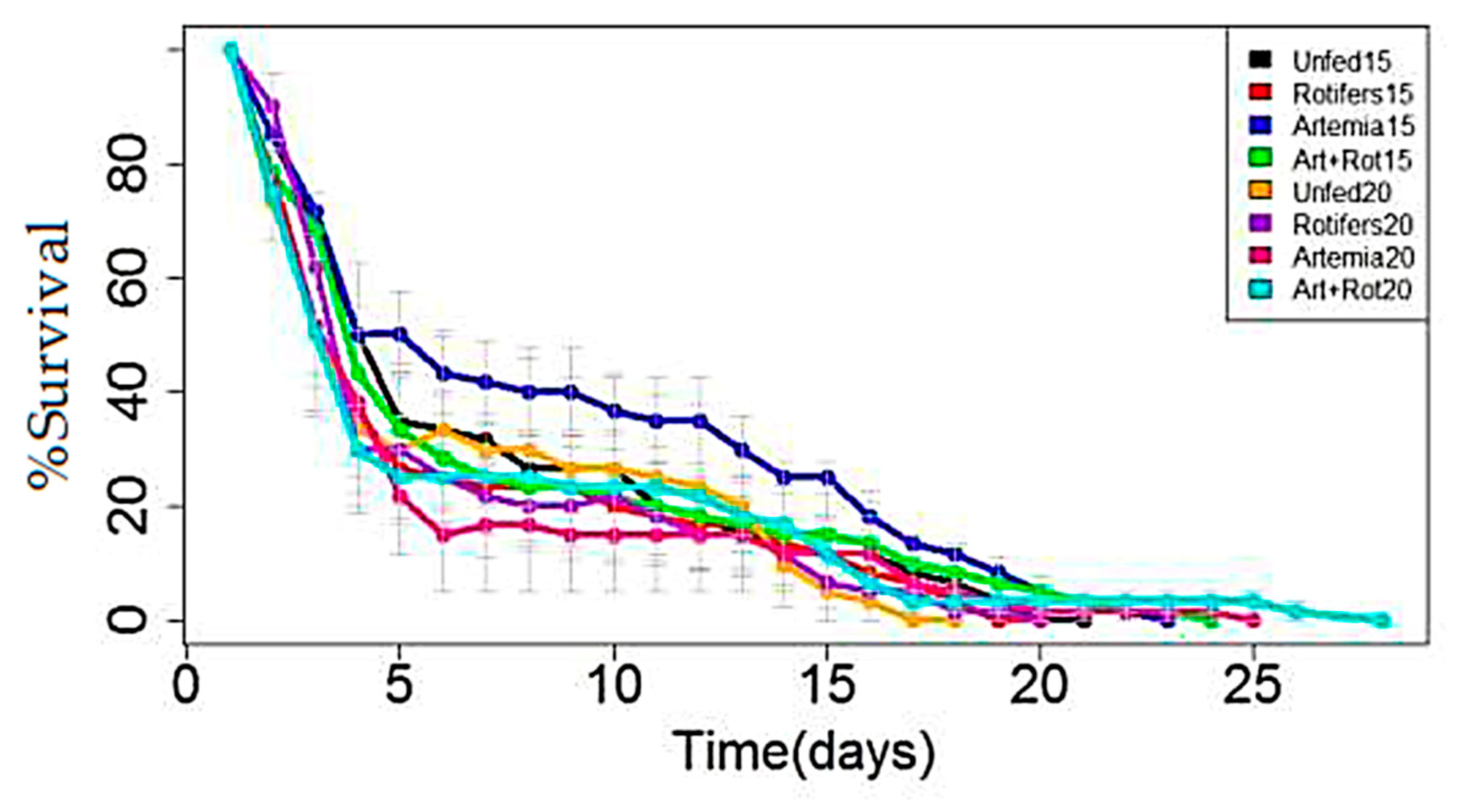
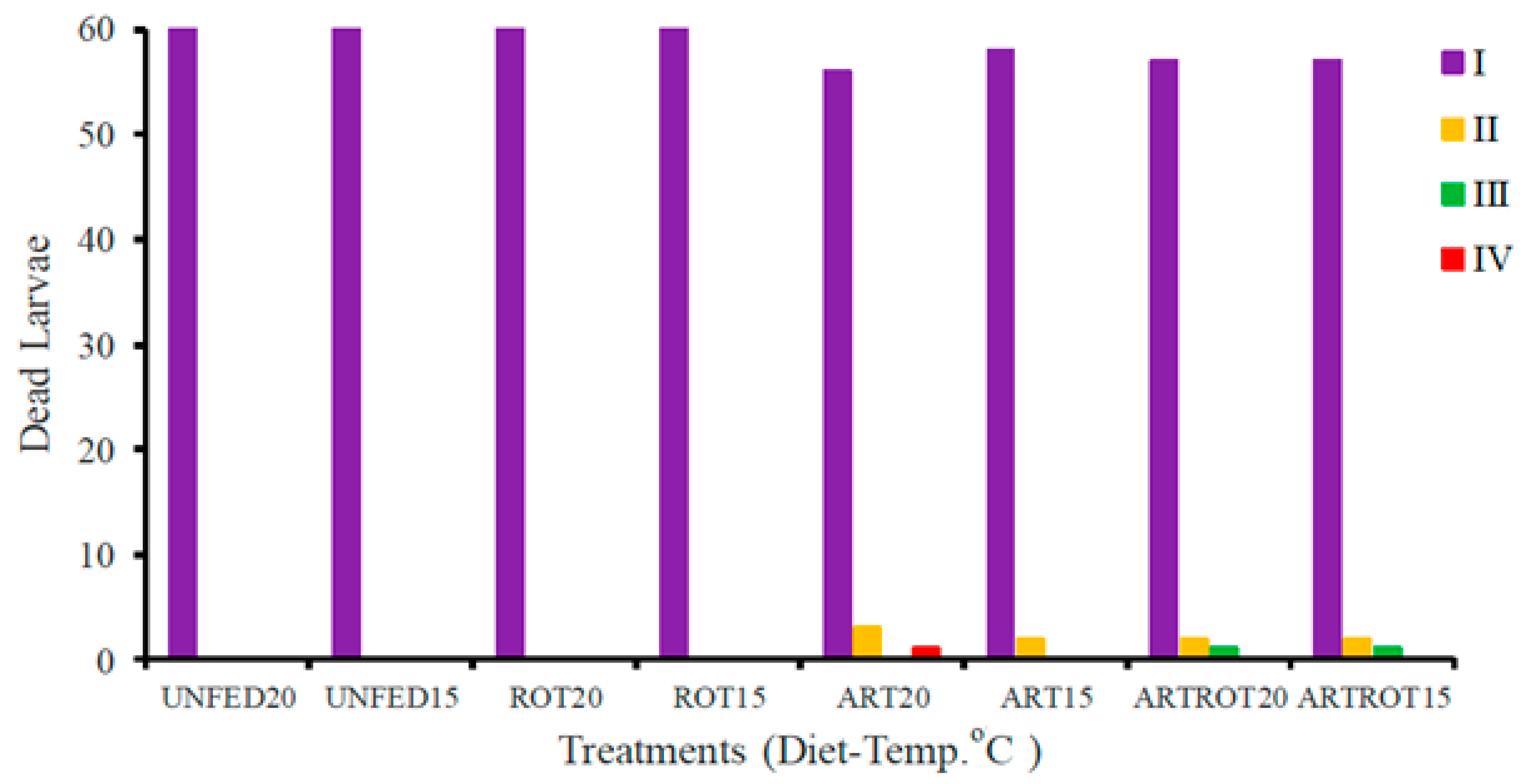
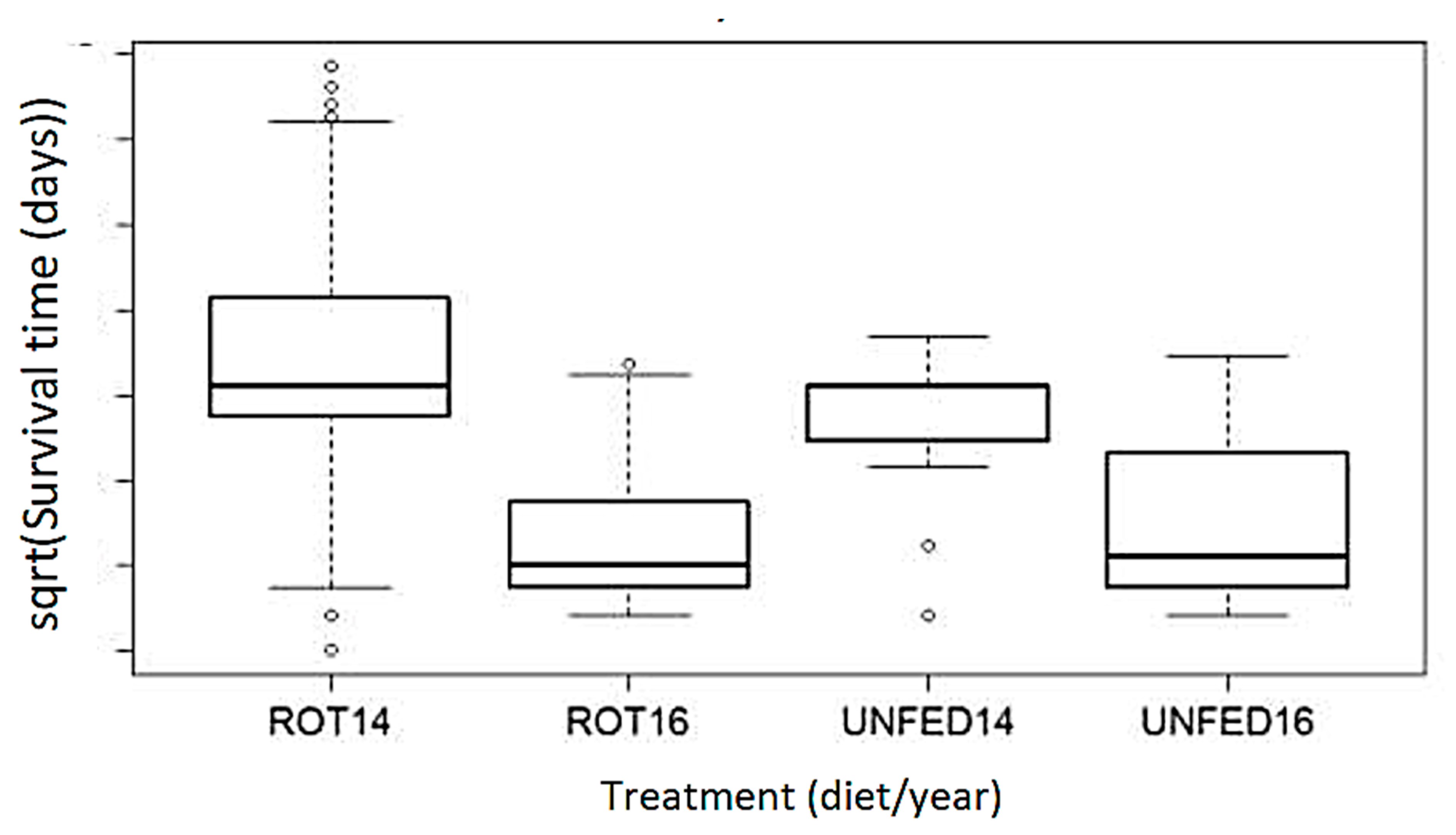
3.1. RDSC Larval Survival
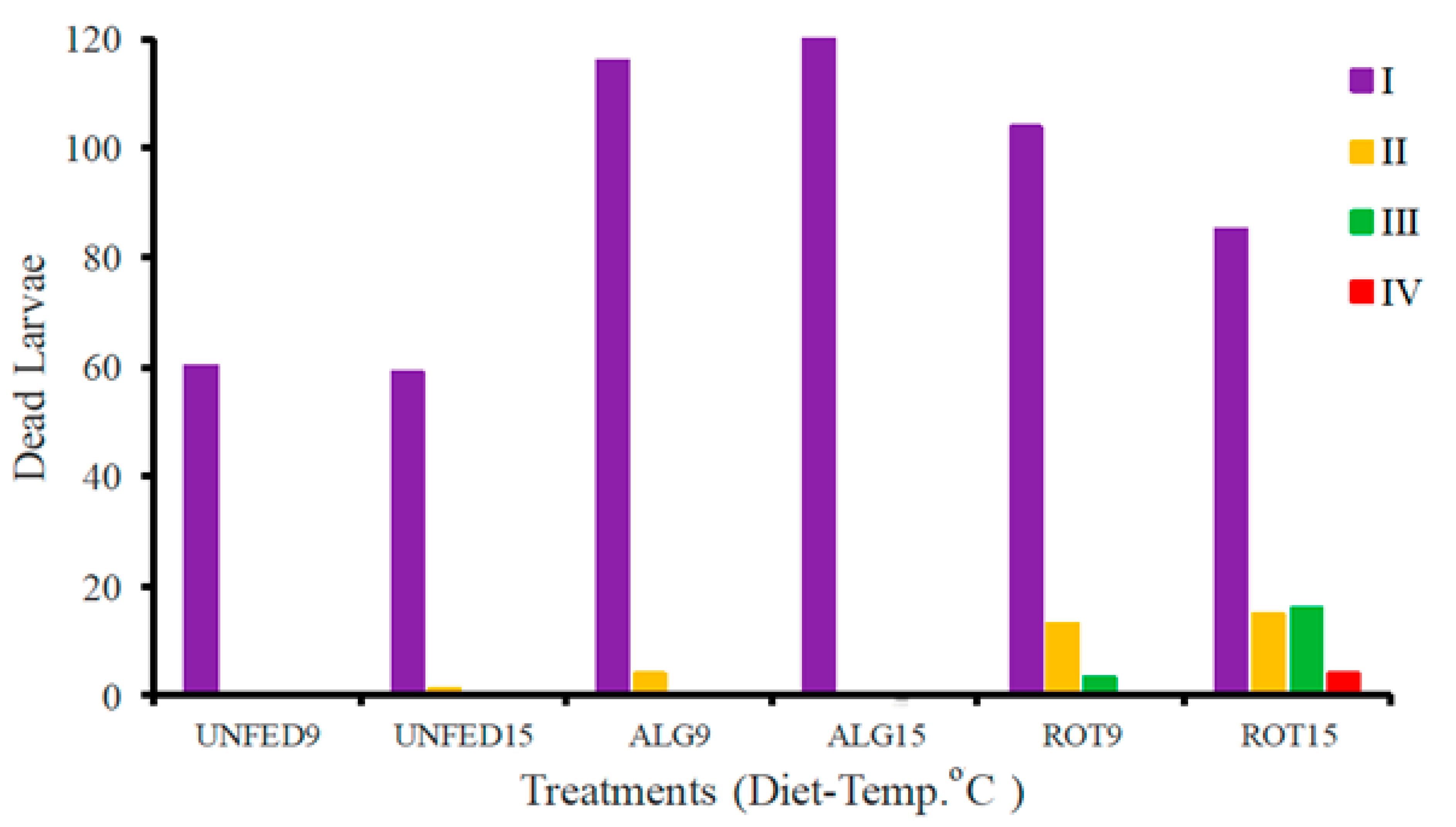
3.2. Larval Development
3.3. Comparison of Survival between the LFSI and LFSII Systems
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.I. The stalk-eyed crustaceans of the Atlantic coast of North America north of 508 Cape Cod. Trans. Conn. Acad. Arts Sci. 1879, 5, 27–136. [Google Scholar]
- Haefner, P.A., Jr.; Musick, J.A. Observations on distribution and abundance of red crabs in Norfolk Canyon and adjacent continental slope. Mar. Fish. Rev. 1974, 36, 31–34. [Google Scholar]
- Wigley, R.L.; Theroux, R.B.; Murray, H.E. Deep sea red crab, Geryon quinquedens, Survey off northeastern United States. Mar. Fish. Rev. 1975, 37, 1–21. [Google Scholar]
- Steimle, F.W.; Zetlin, C.; Chang, S. Essential Fish Habitat Source Document: Red Deepsea Crab, Chaceon (Geryon) quinquedens, Life History and Habitat Characteristics. In NOAA Technical Memorandum NMFS-NE-163; NOAA: Wood Hole, MA, USA, 2001. [Google Scholar]
- Hastie, L.C. Deep-water Geryonid crabs: A continental slope resource. Oceanogr. Mar. Biol. Annu. Rev. 1995, 33, 561–584. [Google Scholar]
- Chute, A.; Jacobson, L.; Redding, B. Clarification of Methods Used to Determine Deep Sea Red Crab (Chaceon Quinquedens) Abundance from Data Collected during the 1974 Photographic Survey. In USA Department of Commerce, Northeast Fisheries Science Center Reference Document 13–22; NOAA: Washington, DC, USA, 2013; 19p. [Google Scholar]
- Wahle, R.A.; Bergeron, C.E.; Chute, A.S.; Jacobson, L.D.; Chen, Y. The Northwest Atlantic deep-sea red crab (Chaceon quinquedens) population before and after the onset of harvesting. ICES J. Mar. Sci. 2008, 65, 862–872. [Google Scholar] [CrossRef]
- NEFMC (New England Fishery Management Council). Atlantic Deep-Sea Red Crab Fishing Years 2014–2016 Specifications, Including A Regulatory Flexibility Analysis; New England Fisheries Management Council, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Northeast Region: Newburyport, MA, USA, 2016. [Google Scholar]
- NEFMC (New England Fishery Management Council). Amendment 3 Fishery Management Plan for Deep-Sea Red Crab (Chaceon Quinquedens) (FMP) Incorporating Specifications for Fishing Years 2011–2013 (March 1, 2011–February 28, 2014) and Including an Environmental Assessment (EA), Regulatory Impact Review (RIR), and Initial Regulatory Flexibility Analysis (IRFA); New England Fishery Management Council, National Marine Fisheries Service: Newburyport, MA, USA, 2014. [Google Scholar]
- Martínez-Rivera, S.; Stevens, B.G. Embryonic development and fecundity of the red deep-sea crab Chaceon quinquedens (Smith, 1879) (Decapoda: Brachyura: Geryonidae) in the Mid-Atlantic Bight determined by image analysis. J. Crustac. Biol. 2020, 40, 230–236. [Google Scholar] [CrossRef]
- Martínez-Rivera, S.; Long, W.C.; Stevens, B.G. Physiological and behavioral sexual maturity of female red deep-sea crabs Chaceon quinquedens (Smith, 1879) (Decapoda: Brachyura: Geryonidae) in the Mid-Atlantic Bight. J. Crustac. Biol. 2020, 40, 330–340. [Google Scholar] [CrossRef]
- Hines, A.H. Larval Patterns in the Life Histories of Brachyuran Crabs (Crustacean, Decapoda, Brachyura). Bull. Mar. Sci. 1986, 39, 444–466. [Google Scholar]
- Perkins, H.C. The larval stages of the deep sea red crab, Geryon quinquedens Smith, reared under laboratory conditions (Decapoda: Brachyrhyncha). Fish. Bull. 1973, 71, 69–82. [Google Scholar]
- Okamoto, K.; Atsumi, S.; Matsuyama, H. Complete larval development of the deep sea red crab Chaceon granulatus under laboratory conditions. Aquac. Sci. 2005, 53, 93–94. [Google Scholar]
- Onishi, K. Larval breeding of a deep-sea crab, Geryon trispinosus (Herbst). Bull. Shizuoka Pref. Fish. Exp. Stn. 1982, 16, 87–95. [Google Scholar]
- Sulkin, S.D. The significance of diet in the growth and development of larvae of the blue crab, Callinectes sapidus rathbun, under laboratory conditions. J. Exp. Mar. Biol. Ecol. 1975, 20, 119–135. [Google Scholar] [CrossRef]
- Sulkin, S.D. Nutritional requirements during larval development of the portunid crab, Callinectes sapidus Rathbun. J. Exp. Mar. Biol. Ecol. 1978, 34, 29–41. [Google Scholar] [CrossRef]
- Rosowski, M.C. The Effect of Temperature on Growth and Dispersal of Larvae of Geryon quinquedens Smith (Brachyura; Geryonidae). Master’s Thesis, University of Delaware, Newark, DE, USA, 1979. [Google Scholar]
- Sulkin, S.D.; Van Heukelem, W.F. Ecological and evolutionary significance of nutritional flexibility in planktotrophic larvae of the Deep Sea red crab Geryon quinquedens and the Stone Crab Menippe mercenaria*. Mar. Ecol. Prog. Ser. 1980, 2, 91–95. [Google Scholar] [CrossRef]
- Van Heukelem, W.; Christman, M.C.; Epifanio, C.E.; Sulkin, S.D. Growth of Geryon quinquedens (Brachyura: Geryonidae) juveniles in the laboratory. Fish. Bull. 1983, 81, 903–905. [Google Scholar]
- Farrelly, C.; Sulkin, S.D. Effect of starvation on megalopal development in the brachyuran crab Hemigrapsus oregonensis. J. Crustacean Biol. 1988, 8, 614–617. [Google Scholar] [CrossRef]
- Staton, J.L.; Sulkin, S.D. Nutritional requirements and starvation resistance in larvae of the brachyuran crabs Sesarma cinereum (Bosc) and S. reticulatum (Say). J. Exp. Mar. Biol. Ecol. 1991, 152, 271–284. [Google Scholar] [CrossRef]
- Anger, K.; Dawirs, R.R.; Anger, V.; Costlow, J.D. Effects of early starvation periods on zoeal development of brachyuran crabs. Biol. Bull. 1981, 161, 199–212. [Google Scholar] [CrossRef]
- Epelbaum, A.B.; Kovatcheva, N.P. Daily food intakes and optimal food concentrations for red king crab (Paralithodes camtschaticus) larvae fed Artemia nauplii under laboratory conditions. Aquac. Nutr. 2005, 11, 455–461. [Google Scholar] [CrossRef]
- Stevens, B.G.; Swiney, K.M. Hatch Timing, Incubation Period, and Reproductive Cycle for Captive Primiparous and Multiparous Red King Crab, Paralithodes Camtschaticus. J. Crustac. Biol. 2007, 27, 37–48. [Google Scholar] [CrossRef]
- Perez-Perez, N.M. Hatching Periodicity, Survival and Development of the Early Life Stages of Red Deepsea Crab (Chaceon Quinquedens) in Laboratory Conditions: Effects of Diet and Temperature; Natural Resources Graduate Program, Delaware State University: Dover, DE, USA, 2017; p. 116. [Google Scholar]
- van Heuven, S.; Pierrot, S.D.; Rae, J.W.B.; Lewis, E.; Wallace, D.W.R. MATLAB Program Developed for CO2 System Calculations. ORNL/CDIAC-105b. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy: Oak Ridge, TN, USA, 2011. [Google Scholar] [CrossRef]
- Lewis, E.; Wallace, D. Program Developed for CO2 System Calculations; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory: Oak Ridge, TN, USA, 1998; p. 38. [Google Scholar]
- Stevens, B.G.; Guida, V. Depth and temperature distribution, morphometrics, and sex ratios of red deepsea crab (Chaceon quinquedens) at 4 sampling sites in the Mid-Atlantic Bight. Fish. Bull. 2016, 114, 343–359. [Google Scholar] [CrossRef]
- Brown, M.B.; Forsythe, A.B. Robust tests for the equality of variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- Hinz, S.; Sulkin, S.; Strom, S.; Testermann, J. Discrimination in ingestion of protistan prey by larval crabs. Mar. Ecol. Prog. Ser. 2001, 222, 155–162. [Google Scholar] [CrossRef]
- Waiho, K.; Fazhan, H.; Quinitio, E.T.; Baylon, J.C.; Fujaya, Y.; Azmie, G.; Wu, Q.; Shi, X.; Ikhwanuddin, M.; Ma, H. Larval rearing of mud crab (Scylla): What lies ahead. Aquaculture 2018, 493, 37–50. [Google Scholar] [CrossRef]
- Suprayudi, M.A.; Takeuchi, T.; Hamasaki, K. Effects of Artemia enriched with eicosapentaenoic and docosahexaenoic acid on survival and occurrence of molting failure in megalop larvae of the mud crab Scylla serrata. Fish. Sci. 2004, 70, 650–658. [Google Scholar] [CrossRef]
- Perez, M.; Sulkin, S. Palatability of autotrophic dinoflagellates to newly hatched larval crabs. Mar. Biol. 2005, 146, 771–780. [Google Scholar] [CrossRef]
- Sulkin, S.; Blanco, A.; Chan, J.; Bryant, M. Effects of limiting access to prey on development of first zoeal stage of the brachyuran crabs Cancer magister and Hemigrapsus oregonensis. Mar. Biol. 1998, 131, 515–521. [Google Scholar] [CrossRef]
- Sulkin, S.; Lehto, J.; Strom, S.; Hutchinson, D. Nutritional role of protists in the diet of first stage larvae of the Dungeness crab Cancer magister. Mar. Ecol. Prog. Ser. 1998, 169, 237–242. [Google Scholar] [CrossRef]
- Sulkin, S.D.; McKeen, G.L. Laboratory study of survival and duration of individual zoeal stages as a function of temperature in the brachyuran crab Cancer magister. Mar. Biol. 1989, 103, 31–37. [Google Scholar] [CrossRef]
- Lehto, J.; Sulkin, S.; Strom, S.; Johnson, D. Protists and detrital particles as prey for the first larval stage of the brachyuran crab, Hemigrapsus oregonensis. J. Exp. Mar. Biol. Ecol. 1998, 230, 213–224. [Google Scholar] [CrossRef]
- Gimenez, L.; Anger, K. Effects of temporary food limitation on survival and development of brachyuran crab larvae. J. Plankton Res. 2005, 27, 485–494. [Google Scholar] [CrossRef]
- Rhyne, A.L.; Penha-Lopes, G.; Lin, J. Growth, development, and survival of larval Mithraculus sculptus (Lamark) and Mithraculus forceps (A. Milne Edwards) (Decapoda: Brachyura: Majidae): Economically important marine ornamental crabs. Aquaculture 2005, 245, 183–191. [Google Scholar] [CrossRef]
- Guarizo, M.; Costa, T.M.; Marochi, M.Z. Effect of diet during larval development of Menippe nodifrons Stimpson, 1859 and Callinectes danae Smith, 1869. Aquac. Int. 2020, 28, 1969–1980. [Google Scholar] [CrossRef]
- Nghia, T.T.; Wille, M.; Vandendriessche, S.; Vinh, Q.T.; Sorgeloos, P. Influence of highly unsaturated fatty acids in live food on larviculture of mud crab Scylla paramamosain (Estampador 1949). Aquac. Res. 2007, 38, 1512–1528. [Google Scholar] [CrossRef]
- Andrés, M.; Estévez, A.; Rotllant, G. Growth, Survival and biochemical composition of spider crab Maja brachydactyla (Balss, 1922) (Decapoda: Majidae) larvae reared under different stocking densities, prey: Larva ratios and diets. Aquaculture 2007, 273, 494–502. [Google Scholar] [CrossRef]
- Suprayudi, M.A.; Takeuchi, T.; Hamasaki, K. Cholesterol Effect on Survival and Development of Larval Mud Crab Scylla serrata. HAYATI J. Biosci. 2012, 19, 1–5. [Google Scholar] [CrossRef]
- Baylon, J.C.; Bravo, M.E.A.; Maningo, N.C. Ingestion of Brachionus plicatilis and Artemia salina nauplii by mud crab Scylla serrata larvae. Aquac. Res. 2004, 35, 62–70. [Google Scholar] [CrossRef]
- Genodepa, J.; Southgate, P.C.; Zeng, C. Diet particle size preference and optimal ration for mud crab, Scylla serrata, larvae fed microbound diets. Aquaculture 2004, 230, 493–505. [Google Scholar] [CrossRef]
- Hamasaki, K.; Suprayudi, M.A.; Takeuchi, T. Effect of dietary n-3HUFA on larval morphogenesis and metamorphosis to megalops in the seed production of mud crab, Scylla serrata (Brachyura: Portunidae). Aquac. Sci. 2002, 50, 333–340. [Google Scholar]
- Baylon, J.C. Effects of salinity and temperature on survival and development of larvae and juveniles of the mud crab, Scylla serrata (Crustacea: Decapoda: Portunidae). J. World Aquac. Soc. 2010, 41, 858–873. [Google Scholar] [CrossRef]
- Gong, J.; Yu, K.; Shu, L.; Ye, H.; Li, S.; Zeng, C. Evaluating the effects of temperature, salinity, starvation and autotomy on molting success, molting interval and expression of ecdysone receptor in early juvenile mud crabs, Scylla paramamosain. J. Exp. Mar. Biol. Ecol. 2015, 464, 11–17. [Google Scholar] [CrossRef]
- Azra, M.N.; Aaqillah-Amr, M.A.; Ikhwanuddin, M.; Ma, H.; Waiho, K.; Ostrensky, A.; Tavares, C.P.D.S.; Abol-Munafi, A.B. Effects of climate-induced water temperature changes on the life history of brachyuran crabs. Rev. Aquac. 2020, 12, 1211–1216. [Google Scholar] [CrossRef]
- Ikhwanuddin, M.; Azra, M.N.; Talpur, M.A.; Abol-Munafi, A.B.; Shabdin, M.L. Optimal water temperature and salinity for production of blue swimming crab, Portunus pelagicus 1st day juvenile crab. Aquac. Aquar. Conserv. Legis. 2012, 5, 4–8. [Google Scholar]
- Bryars, S.R.; Havenhand, J.N. Effects of constant and varying temperatures on the development of blue swimmer crab (Portunus pelagicus) larvae: Laboratory observations and field predictions for temperate coastal waters. J. Exp. Mar. Biol. Ecol. 2006, 329, 218–229. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | * Diet | Temperature | Number of Larvae | Number of Broods |
|---|---|---|---|---|
| 2014 LFSI Flow-Through | U | 9 °C, 15 °C | 60 | 1 |
| R | 9 °C, 15 °C | 60 | 1 | |
| A | 9 °C, 15 °C | 60 | 1 | |
| A + R | 9 °C, 15 °C | 60 | 1 | |
| A + AL | 9 °C, 15 °C | 60 | 1 | |
| U | 9 °C, 15 °C | 60 | 1 | |
| R | 9 °C, 15 °C | 120 | 1 | |
| AL | 9 °C, 15 °C | 120 | 1 | |
| 2016 LFSII Recirculating | U | 15 °C, 20 °C | 60 | 7 |
| R | 15 °C, 20 °C | 60 | 7 | |
| A | 15 °C, 20 °C | 60 | 7 | |
| A + R | 15 °C, 20 °C | 60 | 7 |
| Experiment | * Diet | Concentration (Number of Diet/mL) | Amount Fed per Container (mL) |
|---|---|---|---|
| 2014 LFSI Flow-Through | U | 0 | 0 |
| R | 5–3 | 40–7 | |
| ** A | 5 | 20 | |
| ** A/R | 2.5/5 | 20/range from 40 to 25 | |
| ** A/AL | 2.5/10,000,000 | 20/range from 50 to 20 | |
| U | 0 | 0 | |
| R | range from 5 to 3 | range from 40 to 7 | |
| AL | 10,000,000 | range from 30 to 10 | |
| 2016 LFSII Recirculating | U | 0 | 0 |
| R | range from 7 to 4 | range from 18 to 2 | |
| A | range from 0.7 to 0.5 | range from 7 to 3 | |
| A/R | range from 0.4 to 0.3/from 3.5 to 2.5 | range from 4 to 1.5/from 9 to 1 |
| Treatment | F | df | p | |
|---|---|---|---|---|
| Unfed9 | Containers | 5.5949 | 2 | 0.06097 |
| Tanks | 1.8747 | 1 | 0.1709 | |
| Unfed15 | Containers | 3.3506 | 2 | 0.1873 |
| Tanks | 0.0047866 | 1 | 0.9448 | |
| Rotifers9 | Containers | 3.0138 | 2 | 0.2216 |
| Tanks | 6.9637 | 1 | * 0.008318 | |
| Rotifers15 | Containers | 25.956 | 2 | * 2.31 × 10−06 |
| Tanks | 3.8738 | 1 | * 0.04905 | |
| Algae9 | Containers | 5.5421 | 2 | 0.0626 |
| Tanks | 1.0977 | 1 | 0.2948 | |
| Algae15 | Containers | 6.5451 | 2 | * 0.03791 |
| Tanks | 0.061819 | 1 | 0.8036 |
| Treatment | F | df | p |
|---|---|---|---|
| Unfed15 | 3.2634 | 2 | 0.1956 |
| Unfed20 | 13.975 | 2 | * 0.00092 |
| Rotifers15 | 10.563 | 2 | * 0.00509 |
| Rotifers20 | 9.0347 | 2 | * 0.01092 |
| Artemia15 | 2.5972 | 2 | 0.2729 |
| Artemia20 | 16.506 | 2 | * 0.00026 |
| Art + Rot15 | 17.881 | 2 | * 0.00013 |
| Art + Rot20 | 1.5134 | 2 | 0.4692 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Pérez, N.M.; Poach, M.; Stevens, B.; Smith, S.L.; Ozbay, G. Diet and Temperature Effects on the Survival of Larval Red Deep-Sea Crabs, Chaceon quinquedens (Smith, 1879), under Laboratory Conditions. J. Mar. Sci. Eng. 2023, 11, 1064. https://doi.org/10.3390/jmse11051064
Pérez-Pérez NM, Poach M, Stevens B, Smith SL, Ozbay G. Diet and Temperature Effects on the Survival of Larval Red Deep-Sea Crabs, Chaceon quinquedens (Smith, 1879), under Laboratory Conditions. Journal of Marine Science and Engineering. 2023; 11(5):1064. https://doi.org/10.3390/jmse11051064
Chicago/Turabian StylePérez-Pérez, Nivette M., Matthew Poach, Bradley Stevens, Stacy L. Smith, and Gulnihal Ozbay. 2023. "Diet and Temperature Effects on the Survival of Larval Red Deep-Sea Crabs, Chaceon quinquedens (Smith, 1879), under Laboratory Conditions" Journal of Marine Science and Engineering 11, no. 5: 1064. https://doi.org/10.3390/jmse11051064