
A Snapshot of the Taxonomic Composition and Metabolic Activity of the Microbial Community in an Arctic Harbour (Ny-Ålesund, Kongsfjorden, Svalbard)
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
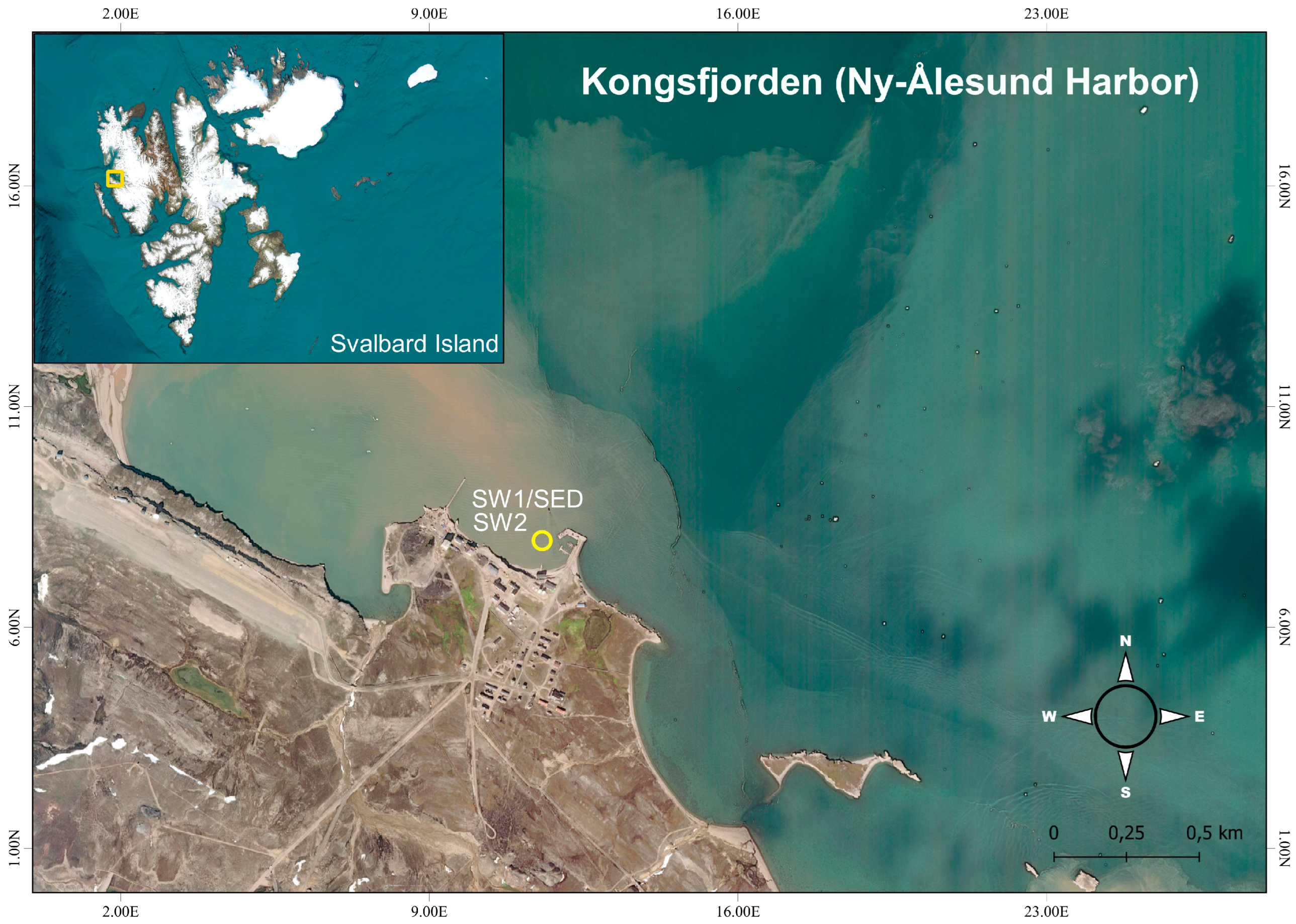
2.1. Sampling in the Study Area
2.2. Measurement of the Hydrocarbon Concentration (GC-FID Analysis)
2.3. Total Bacterial Count (DAPI)
2.4. Culturable Heterotrophic Bacterial Count (Marine Agar)
2.5. Hydrocarbon-Degrading Bacterial Count (MPN)
2.6. 16S rDNA Clone Library
Extraction and PCR Amplification of Total DNA, and 16S rRNA Cloning and Sequencing
2.7. Quantitative Determination of Enzymatic Activity Rates
2.8. Isolation of Psychrophilic Heterotrophic Bacteria
2.9. 16S rDNA Gene Phylogenetic Analyses
2.10. Qualitative Study of Enzymatic Expression of Bacterial Isolates
3. Results
3.1. Environmental Characteristics
3.2. Analysis of Hydrocarbons (GC-FID Analysis)
3.3. Total Bacterial (DAPI), Heterotrophic Bacterial (CFU), and Hydrocarbon-Degrading Bacterial Counts (MPN)
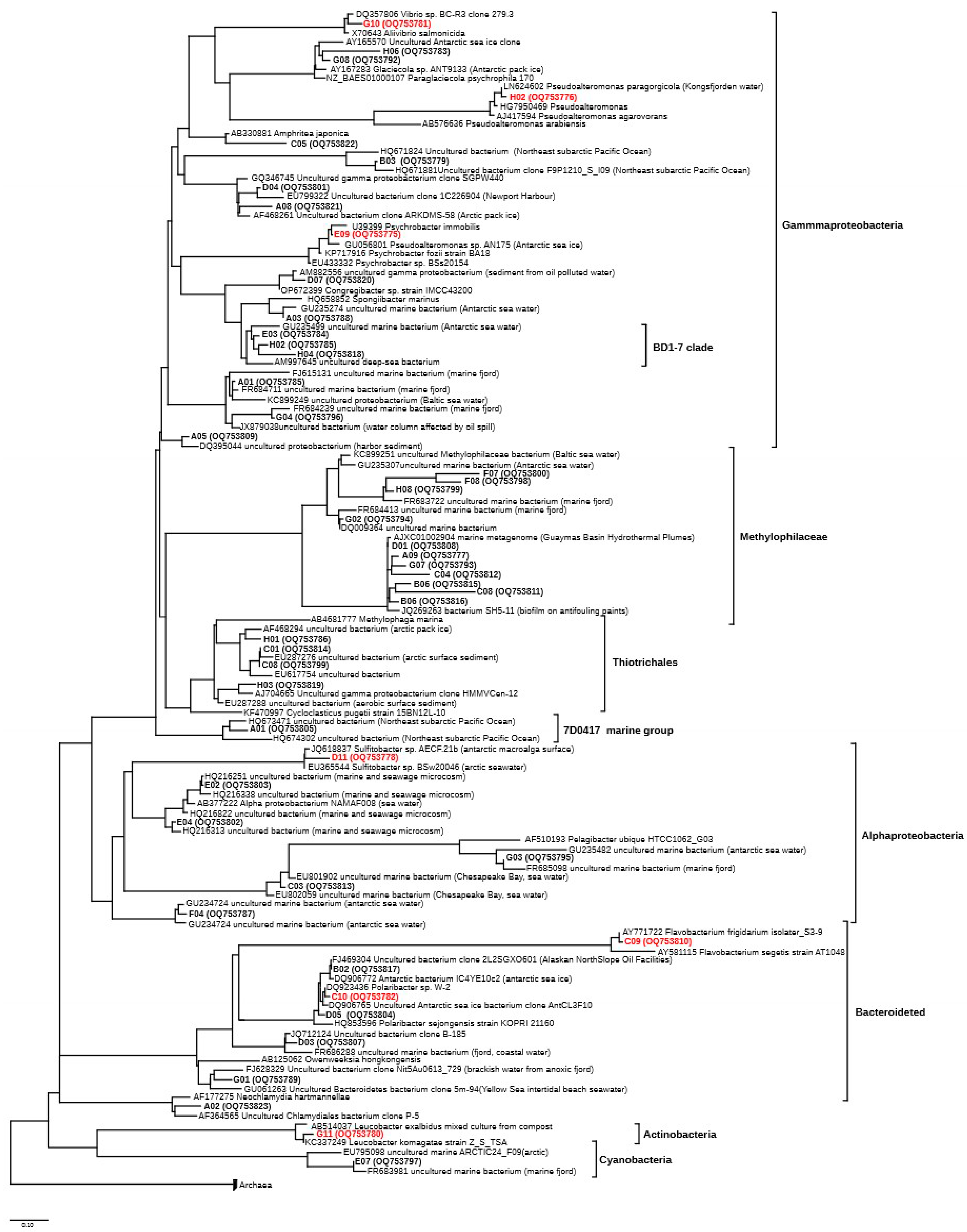
3.4. Structure of the Microbial Community (16S rRNA Clone Library)
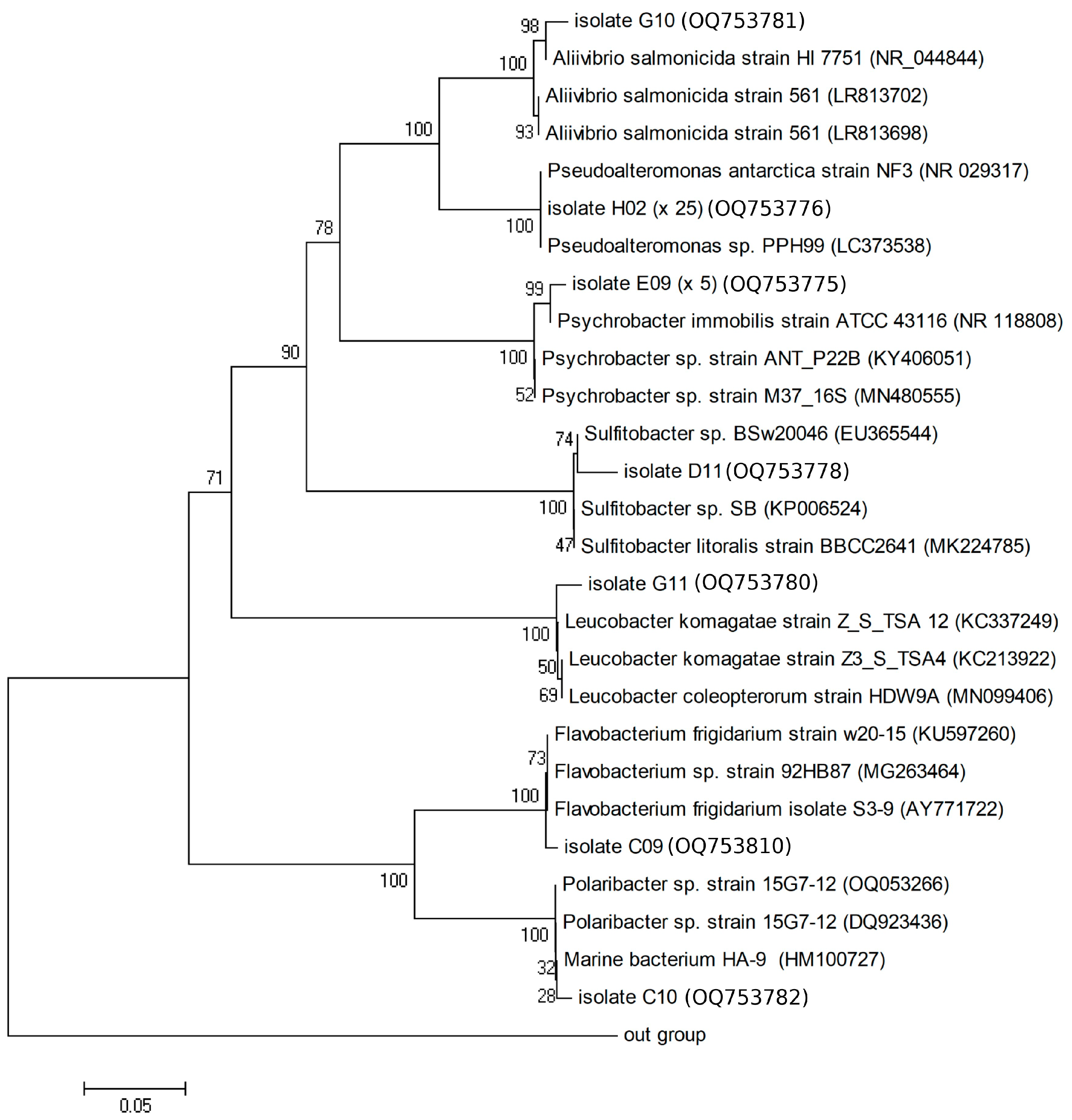
3.5. Taxonomic Identification of the Isolated Bacteria
3.6. Extracellular Enzymatic Activity Rates
3.7. Enzymatic Expression of Bacterial Isolates
4. Discussion
4.1. Bacterial Abundance in the Ny-Ålesund Harbour
4.2. Taxonomic Composition of the Microbial Community
4.3. Qualitative and Quantitative Characterisation of Microbial Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, Y.S.; Tam, N.F.Y.; Lau, P.S.; Xue, X.Z. The toxicity of marine sediments in Victoria Harbour, Hong Kong. Mar. Pollut. Bull. 1995, 31, 464–470. [Google Scholar] [CrossRef]
- Fabiano, M.; Marrale, D.; Misic, C. Bacteria and organic matter dynamics during a bioremediation treatment of organic-rich harbour sediments. Mar. Poll. Bull. 2003, 46, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Misic, C.; Covazzi Harriague, A. Organic matter characterisation and turnover in the sediment and seawater of a tourist harbour. Mar. Environ. Res. 2009, 68, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Danulat, E.; Muniz, P.; Garcia-Alonso, J.; Yannicelli, B. First assessment of the highly contaminated harbour of Montevideo, Uruguay. Mar. Poll. Bull. 2002, 44, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Guerra-García, J.M.; García-Gómez, J.C. Oxygen levels versus chemical pollutants: Do they have similar influence on macrofaunal assemblages? A case study in a harbour with two opposing entrances. Environ. Pollut. 2005, 135, 281–291. [Google Scholar] [CrossRef]
- Todorova, N.H.; Mironova, R.S.; Karamfilov, V.K. Comparative molecular analysis of bacterial communities inhabiting pristine and polluted with polycyclic aromatic hydrocarbons Black Sea coastal sediments. Mar. Pollut. Bull. 2014, 83, 231–240. [Google Scholar] [CrossRef]
- Caruso, G.; Giacobbe, M.G.; Azzaro, F.; Decembrini, F.; Leonardi, M.; Miserocchi, S.; Cao, X.; Song, C.; Zhou, Y. All-In-One: Microbial Response to Natural and Anthropogenic Forcings in a Coastal Mediterranean Ecosystem, the Syracuse Bay (Ionian Sea, Italy). J. Mar. Sci. Eng. 2022, 10, 19. [Google Scholar] [CrossRef]
- Blanchard, A.L.; Feder, H.M.; Shaw, D.G. Variations in benthic fauna underneath an effluent mixing zone at a marine oil terminal in Port Valdez, Alaska. Mar. Pollut. Bull. 2003, 46, 1583–1589. [Google Scholar] [CrossRef]
- Je, J.G.; Belan, T.; Levings, C.; Koo, B.J. Changes in benthic communities along a presumed pollution gradient in Vancouver Harbour. Mar. Environ. Res. 2003, 57, 121–135. [Google Scholar] [CrossRef]
- Caplat, C.; Texier, H.; Barillier, D.; Lelievre, C. Heavy metals mobility in harbour contaminated sediments: The case of Port-en-Bessin. Mar. Pollut. Bull. 2005, 50, 504–511. [Google Scholar] [CrossRef]
- Camphuysen, C.J. IFAW Chronic Oil Pollution in Europe. Status Report 2006; Royal Netherlands Institute for Sea Research: Texel, The Netherlands, 2007; pp. 1–88. [Google Scholar]
- Mali, M.; Dell’Anna, M.M.; Mastrorilli, P.; Damiani, L.; Ungaro, N.; Marinski, J.; Korsachka, M. Integrated Assessment of the Quality of Harbor Sediments: Case Study Based on a Comparative Analysis of Sediments Quality of Two Industrial Ports: Bourgas (BG) and Bari (IT). In Sustainable Development of Sea-Corridors and Coastal Waters; Stylios, C., Floqi, T., Marinski, J., Damiani, L., Eds.; Springer International Publishing: Cham, Switzerland, 2015; Chapter 1; pp. 1–20. [Google Scholar]
- Zaccone, R.; Caruso, G. Microbial enzymes in the Mediterranean Sea: Relationship with climate changes. AIMS Microbiol. 2019, 5, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Wessels Perelo, L. Review: In situ and bioremediation of organic pollutants in aquatic sediments. J. Haz. Mat. 2010, 177, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Misic, C.; Castellano, M.; Covazzi Harriague, A. Organic matter features, degradation and remineralisation at two coastal sites in the Ligurian Sea (NW Mediterranean) differently influenced by anthropogenic forcing. Mar. Environ. Res. 2011, 72, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Ford, T.E. Response of marine microbial communities to anthropogenic stress. J. Aquat. Ecosys. Stress Recov. 2000, 7, 75–89. [Google Scholar] [CrossRef]
- Ford, T.; Jay, J.; Patel, A.; Kile, M.; Prommasith, P.; Galloway, T.; Sanger, R.; Smith, K.; Depledge, M. Use of Ecotoxicological Tools to Evaluate the Health of New Bedford Harbor Sediments: A Microbial Biomarker Approach. Environ. Health Perspect. 2005, 113, 186–191. [Google Scholar] [CrossRef]
- Caruso, G.; Azzaro, M.; Caroppo, C.; Decembrini, F.; Monticelli, L.S.; Leonardi, M.; Maimone, G.; Zaccone, R.; La Ferla, R. Microbial community and its potential as descriptor of environmental status. ICES J. Mar. Sci. 2016, 73, 2174–2177. [Google Scholar] [CrossRef]
- Guibert, L.M.; Loviso, C.L.; Borglin, S.; Jansson, J.K.; Dionisi, H.; Lozada, M. Diverse Bacterial Groups Contribute to the Alkane Degradation Potential of Chronically Polluted Subantarctic Coastal Sediments. Microb. Ecol. 2016, 71, 100–112. [Google Scholar] [CrossRef]
- Dash, H.R.; Mangwani, N.; Chakraborty, J.; Kumari, S.; Das, S. Marine bacteria: Potential candidates for enhanced bioremediation. Appl. Microbiol. Biotechnol. 2013, 97, 561–571. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Caruso, G.; Rizzo, C.; Papale, M.; Azzaro, M. Bacterial communities versus anthropogenic disturbances in the Antarctic coastal marine environment. Environ. Sustain. 2019, 2, 297–310. [Google Scholar] [CrossRef]
- Varotsos, C.A.; Krapivin, V.F. Pollution of Arctic Waters Has Reached a Critical Point: An Innovative Approach to This Problem. Water Air Soil Pollut. 2018, 229, 343. [Google Scholar] [CrossRef]
- Pouch, A.; Zaborska, A.; Mazurkiewicz, M.; Winogradow, A.; Pazdro, K. PCBs, HCB and PAHs in the seawater of Arctic fjords—Distribution, sources and risk assessment. Mar. Pollut. Bull. 2021, 164, 11980. [Google Scholar] [CrossRef] [PubMed]
- Bargiela, R.; Mapelli, F.; Rojo, D.; Chouaia, B.; Tornés, J.; Borin, S.; Richter, M.; Del Pozo, M.V.; Cappello, S.; Gertler, C.; et al. Bacterial population and biodegradation potential in chronically crude oil-contaminated marine sediments are strongly linked to temperature. Sci. Rep. 2015, 5, 11651. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Madonia, A.; Bonamano, S.; Miserocchi, S.; Giglio, F.; Maimone, G.; Azzaro, F.; Decembrini, F.; La Ferla, R.; Piermattei, V.; et al. Microbial abundance and enzyme activity patterns: Response to changing environmental characteristics along a transect in Kongsfjorden (Svalbard Islands). J. Mar. Sci. Eng. 2020, 8, 824. [Google Scholar] [CrossRef]
- Papale, M.; Caruso, G.; Maimone, G.; La Ferla, R.; Lo Giudice, A.; Rappazzo, A.C.; Cosenza, A.; Azzaro, F.; Ferretti, R.; Paranhos, R.; et al. Microbial Community Abundance and Metabolism Close to the Ice-Water Interface of the Blomstrandbreen Glacier (Kongsfjorden, Svalbard): A Sampling Survey Using an Unmanned Autonomous Vehicle. Water 2023, 15, 556. [Google Scholar] [CrossRef]
- Cottier, F.; Tverberg, V.; Inall, M.; Svendsen, H.; Nilsen, F.; Griffiths, C. Water mass modification in an Arctic fjord through cross-shelf exchange: The seasonal hydrography of Kongsfjorden, Svalbard. J. Geophys. Res. Ocean. 2005, 110, C12005. [Google Scholar] [CrossRef]
- Hop, H.; Pearson, T.; Nøst Hegseth, E.; Kovacs, K.M.; Wiencke, C.; Kwasniewski, S.; Eiane, K.; Mehlum, F.; Gulliksen, B.; Wlodarska-Kowalczuk, M.; et al. The marine ecosystem of Kongsfjorden, Svalbard. Polar Res. 2002, 21, 167–208. [Google Scholar] [CrossRef]
- Pedersen, Å.Ø.; Convey, P.; Newsham, K.K.; Mosbacher, J.B.; Fuglei, E.; Ravolainen, V.; Hansen, B.B.; Jensen, T.C.; Augusti, A.; Biersma, E.M.; et al. Five decades of terrestrial and freshwater research at Ny-Ålesund, Svalbard. Polar Res. 2022, 41, 1–39. [Google Scholar] [CrossRef]
- Piquet, A.M.-T.; Scheepens, J.F.; Bolhuis, H.; Wiencke, C.; Buma, A.G.J. Variability of protistan and bacterial communities in two Arctic fjords (Spitsbergen). Polar Biol. 2010, 33, 1521–1536. [Google Scholar] [CrossRef]
- Iversen, K.R.; Seuthe, L. Seasonal microbial processes in a high-latitude fjord (Kongsfjorden, Svalbard): I. Heterotrophic bacteria, picoplankton and nanoflagellates. Polar Biol. 2011, 34, 731–749. [Google Scholar] [CrossRef]
- Prasad, S.; Manasa, P.; Buddhi, S.; Tirunagari, P.; Begum, Z.; Rajan, S.; Shivaji, S. Diversity and Bioprospective Potential (Cold-Active Enzymes) of Cultivable Marine Bacteria from the Subarctic Glacial Fjord, Kongsfjorden. Curr. Microbiol. 2014, 68, 233–238. [Google Scholar] [CrossRef]
- Bourgeois, S.; Kerhervé, P.; Calleja, M.L.; Many, G.; Morata, N. Glacier inputs influence organic matter composition and prokaryotic distribution in a high Arctic fjord (Kongsfjorden, Svalbard). J. Mar. Syst. 2016, 164, 112–127. [Google Scholar] [CrossRef]
- Jain, A.; Krishnan, K.P. Differences in free-living and particle-associated bacterial communities and their spatial variation in Kongsfjorden, Arctic. J. Basic Microbiol. 2017, 57, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Salam, S.; Lekshmi, S.; Silvester, R.; Krishnan, K.P.; Saramma, A.V.; Hatha, A.A.M. Effect of environmental factors on growth and enzyme production of cold adapted bacteria from water and sediment of Kongsfjord, Ny-Alesund, Arctic. J. Environ. Biol. 2017, 38, 579–585. [Google Scholar] [CrossRef]
- Sinha, R.K.; Krishnan, K.P.; Hatha, A.A.M. Diversity of retrievable heterotrophic bacteria in Kongsfjorden, an Arctic fjord. Braz. J. Microbiol. 2017, 48, 51–61. [Google Scholar] [CrossRef]
- Sinha, R.K.; Krishnan, K.P.; Kerkar, S.; David, T.D. Spatio-Temporal Monitoring and Ecological Significance of Retrievable Pelagic Heterotrophic Bacteria in Kongsfjorden, an Arctic Fjord. Indian J. Microbiol. 2017, 57, 116–120. [Google Scholar] [CrossRef]
- Conte, A.; Papale, M.; Amalfitano, S.; Mikkonen, A.; Rizzo, C.; De Domenico, E.; Michaud, L.; Lo Giudice, A. Bacterial community structure along the subtidal sandy sediment belt of a high Arctic fjord (Kongsfjorden, Svalbard Islands). Sci. Total Environ. 2018, 619–620, 203–211. [Google Scholar] [CrossRef]
- Fang, X.M.; Zhang, T.; Li, J.; Wang, N.-F.; Wang, Z.; Yu, L.-Y. Bacterial community pattern along the sediment seafloor of the Arctic fjorden (Kongsfjorden, Svalbard). Antonie Van Leeuwenhoek 2019, 112, 1121–1136. [Google Scholar] [CrossRef]
- Bischof, K.; Convey, P.; Duarte, P.; Gattuso, J.-P.; Granberg, M.E.; Hop, H.; Hoppe, C.; Jiménez, C.; Lisitsyn, L.; Martinez, B.; et al. Kongsfjorden as harbinger of the future Arctic: Knowns, unknowns and research priorities. In The Ecosystem of Kongsfjorden, Svalbard; Hop, H., Wiencke, C., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 537–562. [Google Scholar]
- Srinivas, T.N.R.; Rao, S.; Reddy, P.V.V.; Pratibha, M.S.; Sailaja, B.; Kavya, B.; Kishore, K.H.; Begum, Z.; Singh, S.M.; Shivaji, S. Bacterial diversity and bioprospecting for cold-active lipases, amylases and proteases, from culturable bacteria of Kongsfjorden and Ny-Alesund, Svalbard, Arctic. Curr. Microbiol. 2009, 59, 537–547. [Google Scholar] [CrossRef]
- Thomas, F.A.; Mohan, M.; Krishnan, K.P. Bacterial diversity and their metabolic profiles in the sedimentary environments of Ny-Ålesund, Arctic. Antonie Van Leeuwenhoek 2021, 114, 1339–1360. [Google Scholar] [CrossRef]
- Venkatachalam, S.; Gopinath, A.; Krishnan, K.P. Fjords of the western and northern regions of Svalbard harbour distinct bacterioplankton community structures. World J. Microbiol. Biotechnol. 2023, 39, 57. [Google Scholar] [CrossRef]
- Dowdall, M.; Vicat, K.; Frearson, I.; Gerland, S.; Lind, B.; Shaw, G. Assessment of the radiological impacts of historical coal mining operations on the environment of Ny-Ålesund, Svalbard. J. Environ. Radioact. 2004, 71, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Crisafi, F.; Giuliano, L.; Yakimov, M.M.; Az.zaro, M.; Denaro, R. Isolation and degradation potential of a cold-adapted oil/PAH degrading marine bacterial consortium from Kongsfjorden (Arctic region). Rend. Fis. Acc. Lincei 2016, 27 (Suppl. S1), S261–S270. [Google Scholar] [CrossRef]
- Genovese, M.; Crisafi, F.; Denaro, R.; Cappello, S.; Russo, D.; Calogero, R.; Santisi, S.; Catalfamo, M.; Modica, A.; Smedile, F.; et al. Effective bioremediation strategy for rapid in situ cleanup of anoxic marine sediments in mesocosm oil spill simulation. Front. Microbiol. 2014, 5, 162. [Google Scholar] [CrossRef]
- Denaro, R.; D’Auria, G.; Di Marco, G.; Genovese, M.; Troussellier, M.; Yakimov, M.M.; Giuliano, L. Assessing terminal restriction fragment length polymorphism suitability for the description of bacterial community structure and dynamics in hydrocarbon-polluted marine environments. Environ. Microbiol. 2005, 7, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Cappello, S.; Caruso, G.; Zampino, D.; Monticelli, L.S.; Maimone, G.; Denaro, R.; Tripodo, B.; Troussellier, M.; Yakimov, M.M.; Giuliano, L. Microbial community dynamics during assays of harbor oil spill bioremediation: A microscale simulation study. J. Appl. Microbiol. 2007, 102, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Cappello, S.; Calogero, R.; Santisi, S.; Genovese, M.; Denaro, R.; Genovese, L.; Giuliano, L.; Mancini, G.; Yakimov, M.M. Bioremediation of oil polluted marine sediments: A bio-engineering treatment. Int. Microbiol. 2015, 18, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Kuwae, T.; Hosokawa, Y. Determination of abundance and biovolume of bacteria in sediments by dual staining with 4′,6-diamidino-2-phenylindole and acridine orange: Relationship to dispersion treatment and sediment characteristics. Appl. Environ. Microbiol. 1999, 65, 3407–3412. [Google Scholar] [CrossRef]
- Cappello, S.; Caruso, G.; Bergami, E.; Macrì, A.; Venuti, V.; Majolino, D.; Corsi, I. New insights into the structure and function of the prokaryotic communities colonizing plastic debris collected in King George Island (Antarctica): Preliminary observations from two plastic fragments. J. Hazard. Mater. 2021, 414, 125586. [Google Scholar] [CrossRef]
- Brown, E.J.; Braddock, J.F. Sheen screen, a miniaturized most-probable-number method for enumeration of oil-degrading microorganisms. Appl. Environ. Microbiol. 1990, 56, 3895–3896. [Google Scholar] [CrossRef]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Waste Water, 18th ed.; APHA: Washington, DC, USA, 1992; pp. 1–1100. [Google Scholar]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackerbrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Hoppe, H.-G. Use of fluorogenic model substrate for extracellular enzyme activity (EEA) measurement of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publishers: Boca Raton, FL, USA, 1993; pp. 423–431. [Google Scholar]
- La Ferla, R.; Azzaro, M.; Caruso, G.; Monticelli, L.S.; Maimone, G.; Zaccone, R.; Packard, T.T. Prokaryotic abundance and heterotrophic metabolism in the deep Mediterranean Sea. Adv. Oceanogr. Limnol. 2010, 1, 143–166. [Google Scholar] [CrossRef]
- Ashelford, K.E.; Chuzhanova, N.A.; Fry, J.C.; Jones, A.J.; Weightman, A.J. At least 1 in 20 16S rRNA sequence records currently held in public repositories is estimated to contain substantial anomalies. Appl. Environ. Microbiol. 2005, 71, 7724–7736. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhukumar Buchner, A.; Lai, T.; Steppi, S.; Jobb, G.; Förster, W.; et al. ARB: A software environment for sequence data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef]
- Caruso, G.; Dell’Acqua, O.; Caruso, R.; Azzaro, M. Phenotypic characterization of bacterial isolates from marine waters and plastisphere communities of the Ross Sea (Antarctica). J. Clin. Microbiol. Biochem. Technol. 2022, 8, 1–9. [Google Scholar] [CrossRef]
- Bozal, N.; Tudela, E.; Rossello-Mora, R.; Lalucat, J.; Guinea, J. Pseudoalteromonas antarctica sp. nov., isolated from an Antarctic coastal environment. Int. J. Syst. Bacteriol. 1997, 47, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Juni, E.; Heym, G.A. Psychrobacter immobilis gen. nov., sp. nov.: Genospecies composed of gram-negative, aerobic, oxidase-positive Coccobacilli. Int. J. Syst. Bacteriol. 1986, 36, 388–391. [Google Scholar] [CrossRef]
- Egidius, E.; Wiik, R.; Andersen, K.; Hoff, K.A.; Hjeltnes, B. Vibrio salmonicida sp. nov., a new fish pathogen. Int. J. Syst. Bacteriol. 1986, 36, 518–520. [Google Scholar] [CrossRef]
- Ji, Q.; Yu, Y.; Chen, B. Bacteria Isolated from Arctic Seawater; direct submission to GenBank database 23 December 2007; Polar Research Institute of China, Biology Department: Shangai, China, 2007; Unpublished data. [Google Scholar]
- Barreto, C.C.; Lima, D.V.; de Oliveira, E.C.; Absher, T.M.; Pellizari, V.H. Species-Specific Microbiota Associated with Antarctic Macroalgae; direct submission to GenBank database 3 February 2012; Universidade de Sao Paulo, Instituto de Ciencias Biomedicas: Sao Paulo, Brazil, 2012; Unpublished data. [Google Scholar]
- Skrodenyte-Arbaciauskiene, V.; Pliuraite, V.; Virbickas, T.; Bartasiute, A. Intestinal Microbial Community of Roach (Rutilus rutilus) and Its Changes under the Impact of River Pollution; direct submission to GenBank database 10 December 2012; Institute of Ecology, Nature Research Centre: Vilnius, Lithuania, 2012; Unpublished data. [Google Scholar]
- Lee, H.K.; Sung, K.C.; Cho, H.H.; Lee, Y.K. Diversity of Arctic Bacteria; direct submission to GenBank database 5 October 2004; Korea Polar Research Institute (KOPRI), Microbiology Lab.: Ansan, Korea, 2004; Unpublished data. [Google Scholar]
- Lin, X.; Yi, D.; Wang, C. Investigation of Oceanic Microbial Diversity in Qingdao Fushan Bay; direct submission to GenBank datahase 28 August 2006; First Institute of Oceanography (SOA), Key Lab of Marine Bioactive Substances: Qingdao, China, 2006; Unpublished data.
- Guo, C.Y.; He, J.F.; Zhang, F.; Cai, M.H.; Eang, G.Z. Spatial heterogeneity of a microbial community in Kongsfjorden, Svalbard during late summer 2006 and its relationship to biotic and abiotic factors. Adv. Polar Sci. 2011, 22, 55–66. [Google Scholar] [CrossRef]
- Kalinowska, A.; Ameryk, A.; Jankowska, K. Microbiological Survey in Two Arctic Fjords: Total Bacterial Number and Biomass Comparison of Hornsund and Kongsfjorden. In Impact of Climate Changes on Marine Environments; Zielinski, T., Weslawski, M., Kulinski, K., Eds.; GeoPlanet: Earth and Planetary Sciences; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Vishnupriya, S.; Jabir, T.; Krishnan, K.P.; Hatha, A.A.M. Bacterial community structure and functional profiling of high Arctic fjord sediments. World J. Microbiol. Biotechnol. 2021, 37, 133. [Google Scholar] [CrossRef]
- Brinkmeyer, R.; Knittel, K.; Jurgens, J.; Weyland, H.; Amann, R.; Helmke, E. Diversity and structure of bacterial communities in Arctic versus Antarctic pack ice. Appl. Environ. Microbiol. 2003, 69, 6610–6619. [Google Scholar] [CrossRef] [PubMed]
- Allers, E.; Wright, J.J.; Konwar, K.M.; Howes, C.G.; Beneze, E.; Hallam, S.J.; Sullivan, M.B. Diversity and population structure of Marine Group A bacteria in the Northeast subarctic Pacific Ocean. ISME J. 2013, 7, 256–268. [Google Scholar] [CrossRef] [PubMed]
- von Scheibner, M.; Dorge, P.; Biermann, A.; Sommer, U.; Hoppe, H.G.; Jurgens, K. Impact of warming on phyto-bacterioplankton coupling and bacterial community composition in experimental mesocosms. Environ. Microbiol. 2014, 16, 718–733. [Google Scholar] [CrossRef] [PubMed]
- Zaikova, E.; Walsh, D.A.; Stilwell, C.P.; Mohn, W.W.; Tortell, P.D.; Hallam, S.J. Microbial community dynamics in a seasonally anoxic fjord: Saanich Inlet, British Columbia. Environ. Microbiol. 2010, 12, 172–191. [Google Scholar] [CrossRef] [PubMed]
- Paissé, S.; Coulon, F.; Goni-Urriza, M.; Peperzak, L.; McGenity, T.J.; Duran, R. Structure of bacterial communities along a hydrocarbon contamination gradient in a coastal sediment. FEMS Microbiol. Ecol. 2008, 66, 295–305. [Google Scholar] [CrossRef]
- Guo, F.; School of Life Sciences, Xiamen University, Xiamen, China; Ke, C.-H.; State Key Laboratory of Marine Environmental Science, Xiamen University, Xiamen, China; Huang, Z.-B.; State Key Laboratory of Marine Environmental Science, Xiamen University, Xiamen, China. Bacteria on the Surface of Antifouling Paint Immersed in the Estuary Zone of Jiulong River, Xiamen University, Xiamen, China. Personal communication, 2012. [Google Scholar]
- Duncan, K.E.; Gieg, L.M.; Parisi, V.A.; Tanner, R.S.; Tringe, S.G.; Bristow, J.; Suflita, J.M. Biocorrosive Thermophilic Microbial Communities in Alaskan North Slope Oil Facilities. Environ. Sci. Technol. 2009, 43, 7977–7984. [Google Scholar] [CrossRef]
- Zappalà, G.; Bruzzone, G.; Caruso, G.; Azzaro, M. Development of an automatic sampler for extreme polar environments: First in situ application in Svalbard Islands. Rend. Fis. Acc. Lincei 2016, 27, 251–259. [Google Scholar] [CrossRef]
- Perlinski, P.; Mudryk, Z.J.; Antonowicz, J. Enzymatic activity in the surface microlayer and subsurface water in the harbour channel. Estuar. Coast. Shelf Sci. 2017, 196, 150–158. [Google Scholar] [CrossRef]
- Obayashi, Y.; Suzuki, S. Proteolytic enzymes in coastal surface seawater: Significant activity of endopeptidases and exopeptidases. Limnol. Oceanogr. 2008, 50, 722–726. [Google Scholar] [CrossRef]
- Arnosti, C. Microbial Extracellular Enzymes and the Marine Carbon Cycle. Ann. Rev. Mar. Sci. 2011, 3, 401–425. [Google Scholar] [CrossRef]
- Hughes, S.G. Variations in Enzyme Production as a Function of the Physiological State of Bacteria. Master’s Thesis, University of Edinburgh, Edinburgh, UK, 1971; pp. 1–143. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| SW1 | SW2 | SED1 | |
|---|---|---|---|
| DAPI (CFU mL−1/g−1) | 1.60 × 104 ± 1.52 × 103 | 2.50 × 104 ± 3.00 × 103 | 1.12 × 106 ± 1.1 × 105 |
| Culturable (CFU mL−1/g−1) | 4.90 × 102 ± 4.6 × 101 | 6.10 × 102 ± 5.8 × 101 | 2.1 × 103 ± 1.80 × 102 |
| Hydrocarbon-degrading bacteria (MPN mL−1/g−1) | 1.00 × 102 ± 8.00 × 101 | 1.30 × 102 ± 1.9 × 101 | 2.10 × 102 ± 0.25 × 101 |
| GenBank Accession Number | Isolate Code | Length (bp) | Closest Hit | Accession No. | ID, % |
|---|---|---|---|---|---|
| OQ753781 | G10 | 780 | Aliivibrio salmonicida strain HI 7751 | NR_044844 | 99.3 |
| OQ753776 | H02 | 840 | Pseudoalteromonas antarctica strain NF3 | NR_029317 | 98.7 |
| OQ753775 | E09 | 568 | Psychrobacter immobilis strain ATCC 43116 | NR_118808 | 99.1 |
| OQ753778 | D11 | 630 | Sulfitobacter sp. BSw20046 | EU365544 | 98.1 |
| OQ753810 | C09 | 590 | Flavobacterium frigidarium isolate S3-9 | AY771722 | 98.9 |
| OQ753782 | C10 | 880 | Polaribacter sp. W-2 | DQ923436 | 98.1 |
| OQ753780 | G11 | 680 | Leucobacter komagatae strain Z_S_TSA 12 | KC337249 | 98.2 |
| LAP | GLU | AP | |
|---|---|---|---|
| SW1 (nmol L−1 h−1) | 1.40 ± 0.13 | 1.1 ± 0.10 | 2.7 ± 0.3 |
| SW2 (nmol L−1 h−1) | 1.50 ± 0.14 | 1.30 ± 0.14 | 3.8 ± 0.41 |
| SED (nmol g−1 h−1) | 2.90 ± 3.0 | 3.50 ± 0.32 | 5.2 ± 0.55 |
| SW 1 | SW2 | SED1 | |
|---|---|---|---|
| No. of isolates | 12 | 12 | 8 |
| Alpha-glucosidase | 0.0 ± 0.0 | 9.2 ± 0.8 | 0.0 ± 0.0 |
| Beta-glucosidase | 18.2 ± 1.6 | 18.1 ± 2.0 | 0.0 ± 0.0 |
| Protease | 33.1 ± 3.5 | 33.4 ± 3.4 | 76.0 ± 8.0 |
| Phosphatase | 33.0 ± 2.8 | 33.2 ± 2.9 | 0.0 ± 0.0 |
| Lipase | 24.2 ± 2.2 | 58.1 ± 0.6 | 86.1 ± 0.9 |
| Esterase | 0.0 ± 0.0 | 9.0 ± 0.9 | 0.0 ± 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappello, S.; Smedile, F.; Caruso, G.; Patania, S.; Lunetta, A.; Azzaro, M. A Snapshot of the Taxonomic Composition and Metabolic Activity of the Microbial Community in an Arctic Harbour (Ny-Ålesund, Kongsfjorden, Svalbard). J. Mar. Sci. Eng. 2023, 11, 1018. https://doi.org/10.3390/jmse11051018
Cappello S, Smedile F, Caruso G, Patania S, Lunetta A, Azzaro M. A Snapshot of the Taxonomic Composition and Metabolic Activity of the Microbial Community in an Arctic Harbour (Ny-Ålesund, Kongsfjorden, Svalbard). Journal of Marine Science and Engineering. 2023; 11(5):1018. https://doi.org/10.3390/jmse11051018
Chicago/Turabian StyleCappello, Simone, Francesco Smedile, Gabriella Caruso, Sabrina Patania, Alessia Lunetta, and Maurizio Azzaro. 2023. "A Snapshot of the Taxonomic Composition and Metabolic Activity of the Microbial Community in an Arctic Harbour (Ny-Ålesund, Kongsfjorden, Svalbard)" Journal of Marine Science and Engineering 11, no. 5: 1018. https://doi.org/10.3390/jmse11051018