
Complete Mitochondrial DNA Genomes of Deep-Sea Eels Synaphobranchus brevidorsalis and S. affinis and New Record of S. brevidorsalis from the East Mariana Basin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Collection
2.2. DNA Extraction and Sequencing
2.3. Phylogenetic Relationships
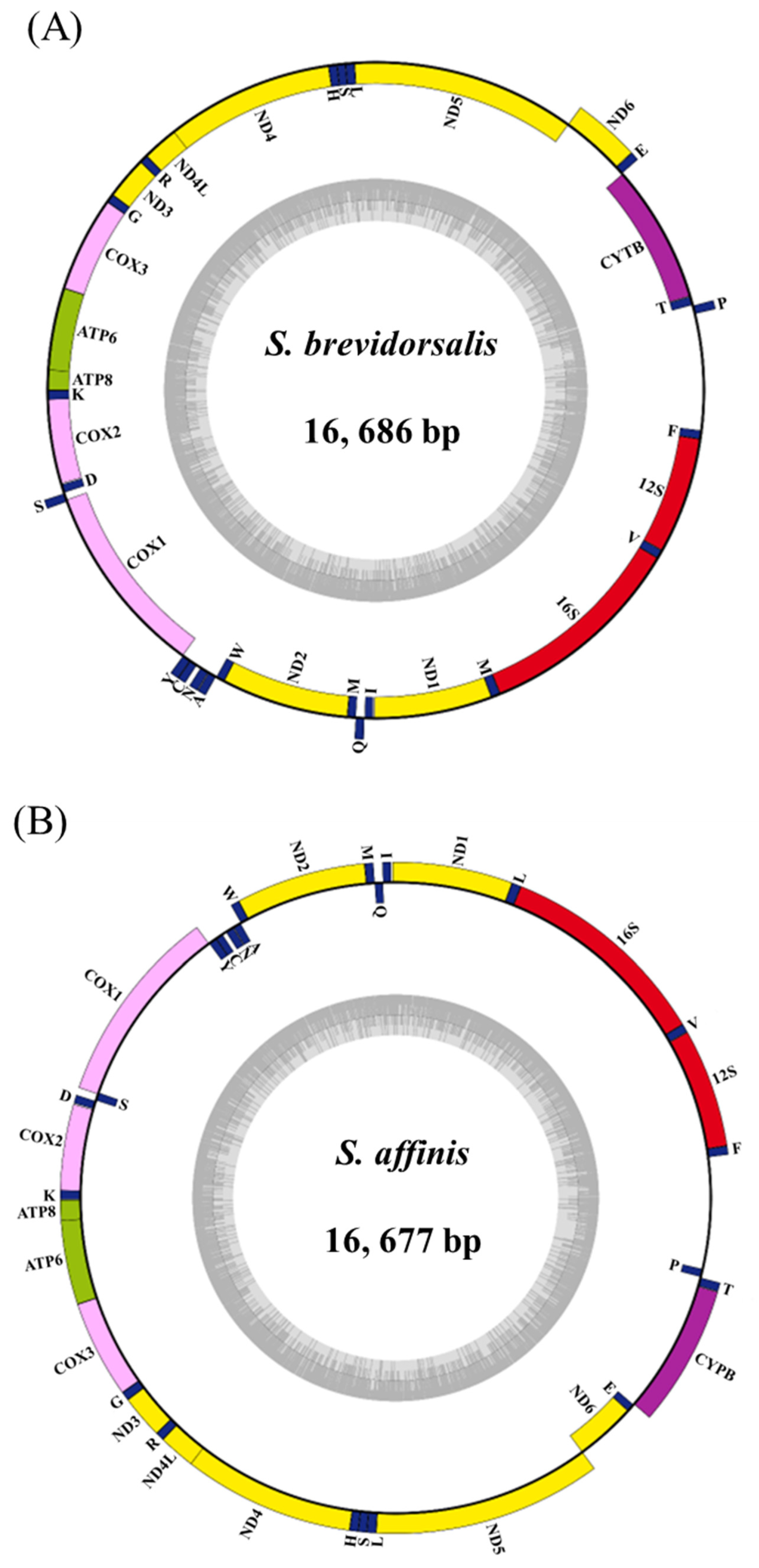
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rex, M.A. Community structure in the deep-sea Benthos. Annu. Rev. Ecol. Syst. 1981, 12, 331–353. [Google Scholar] [CrossRef]
- Woolley, S.N.; Tittensor, D.P.; Dunstan, P.K.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Wintle, B.A.; Worm, B.; O’Hara, T.D. Deepsea diversity patterns are shaped by energy availability. Nature 2016, 533, 393–396. [Google Scholar] [CrossRef]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; Mol, B.D.; Escobar, E.; German, C.R.; Levin, L.A.; Arbizu, P.M.; Menot, L.; Buhl-Mortensen, P.; et al. Deep, diverse and definitely different: Unique attributes of the world’s largest ecosystem. Biogeosciences 2010, 7, 2851–2899. [Google Scholar] [CrossRef]
- Sanders, H.L.; Hessler, R.R. Ecology of the deep-sea benthos. Science 1969, 163, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Robison, B.H. Deep pelagic biology. J. Exp. Mar. Biol. Ecol. 2004, 300, 253–272. [Google Scholar] [CrossRef]
- Naganoa, Y.; Nagahama, T. Fungal diversity in deep-sea extreme environments. Fungal Ecol. 2012, 5, 463–471. [Google Scholar] [CrossRef]
- Linley, T.D.; Gerringer, M.E.; Yancey, P.H.; Drazen, J.C.; Weinstock, C.L.; Jamieson, A.J. Fishesof the hadal zone including new species, in situ observations and depth records of Liparidae. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 114, 99–110. [Google Scholar] [CrossRef]
- Drazen, J.C.; Sutton, T.T. Dining in the Deep: The Feeding Ecology of Deep-Sea Fishes. Annu. Rev. Mar. Sci. 2017, 9, 337–366. [Google Scholar] [CrossRef]
- Elphick, C.S. How you count counts: The importance of methods research in applied ecology. J. Appl. Ecol. 2008, 45, 1313–1320. [Google Scholar] [CrossRef]
- Farnsworth, E.J.; Chu, M.; Kress, W.J.; Neill, A.K.; Best, J.H.; Pickering, J.; Stevenson, R.D.; Courtney, G.W.; VanDyk, J.K.; Ellison, A.M. Next-Generation Field Guides. Bioscience 2013, 63, 891–899. [Google Scholar] [CrossRef]
- Austen, G.E.; Bindemann, M.; Griffiths, R.A.; Roberts, D.L. Species identification by experts and non-experts: Comparing images from field guides. Sci. Rep. 2016, 20, 33634. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Snelgrove, P.V.R. The deep-sea under global change. Curr. Biol. 2017, 27, R461–R465. [Google Scholar] [CrossRef] [PubMed]
- Sulak, K.J.; Shcherbachev, Y.N. Zoogeography and systematics of six deep-living genera of synaphobranchid eels, with a key to taxa and description of two new species of Ilyophis. Bull. Mar. Sci. 1997, 60, 1158–1194. [Google Scholar]
- Eschmeyer, W.N. Catalog of Fishes. Electronic Version. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 4 April 2018).
- Ho, H.-C.; Smith, D.G.; McCosker, J.E.; Hibino, Y.; Loh, K.-H.; Tighe, K.A.; Shao, K.-T. Annotated checklist of eels (orders Anguilliformes and Saccopharyngiformes) from Taiwan. Zootaxa 2015, 4060, 140–189. [Google Scholar] [CrossRef]
- Johnson, J.Y. Descriptions of some new genera and species of fishes obtained at Madeira. In Proceedings of the Zoological Society of London; Blackwell Publishing Ltd.: Oxford, UK, 1862; Volume 30, pp. 167–180. [Google Scholar]
- Günther, A. Preliminary notes on new fishes collected in Japan during the Expedition of H.M.S. “Challenger”. Ann. Mag. Nat. Hist. 1877, 20, 433–446. [Google Scholar] [CrossRef]
- Günther, A. Report on the Deep-Sea Fishes Collected by H.M.S. Challenger during the Years 1873–1876; Wentworth Press: London, UK, 1887; Volume 5, pp. 1–335. [Google Scholar]
- Lea, E. Muraenoid Larvae: From the ”Michael Sars” North Atlantic Deep-Sea Expedition, 1910. Rep. Sci. Res. Sars Exped. 1913, 3, 1–59. [Google Scholar]
- Castle, P.H.J. Deep-Water Eels from Cook Strait, New Zealand; Zoology Publications from Victoria University Wellington: Wellington, New Zealand, 1961; Volume 27, pp. 1–30. [Google Scholar]
- Melo, M.R.S. A new synaphobranchid eel (Anguilliformes: Synaphobranchidae) from Brazil, with comments on the species from the Western South Atlantic. Copeia 2007, 2, 315–323. [Google Scholar] [CrossRef]
- Matarese, A.C.; Spies, I.B.; Busby, M.S.; Orr, J.W. Early larvae of Zesticelus profundorum (family Cottidae) identified using DNA barcoding. Ichthyol. Res. 2011, 58, 170–174. [Google Scholar] [CrossRef]
- Triantafyllidis, A.; Bobori, D.; Koliamitra, C.; Gbandi, E.; Mpanti, M.; Petriki, O.; Karaiskou, N. DNA barcoding analysis of fish species diversity in four north Greek lakes. Mitochondrial DNA 2011, 22, 37–42. [Google Scholar] [CrossRef]
- Zhang, J.B.; Hanner, R. DNA barcoding is a useful tool for the identification of marine fishes from Japan. Biochem. Syst. Ecol. 2011, 39, 31–42. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Timm, J.; Figiel, M.; Kochzius, M. Contrasting patterns in species boundaries and evolution of anemone fishes (Amphiprioninae, Pomacentridae) in the centre of marine biodiversity. Mol. Phylogenet. Evol. 2008, 49, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.; Schneider, H.; Sampaio, I. Genetic differentiation of Macrodon ancylodon (Sciaenidae, Perciformes) populations in Atlantic coastal waters of South America as revealed by mtDNA analysis. Genet. Mol. Biol. 2003, 26, 151–161. [Google Scholar] [CrossRef]
- Vinson, C.; Grazielle, G.; Schneider, H.; Sampaio, I. Sciaenidae fish of the Caeté river estuary, Northern Brazil: Mitochondrial DNA suggests explosive radiation for the Western Atlantic assemblage. Genet. Mol. Biol. 2004, 27, 174–180. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- An, H.-S.; Jee, Y.-J.; Min, K.-S.; Kim, B.-L.; Han, S.-J. Phylogenetic analysis of six pacific abalone (Haiotidae) based on DNA sequences of 16S rRNA and cytochrome c oxidase subunit I mitochondrial genes. Mar. Biotechnol. 2005, 7, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Cabezas, P.; Lin, C.W.; Chan, T.Y. Two new species of the deep-sea squat lobster genus Munida Leach, 1820 (Crustacea: Decapoda: Munididae) from Taiwan: Morphological and molecular evidence. Zootaxa 2011, 3036, 26–38. [Google Scholar] [CrossRef]
- Mu, W.; Liu, J.; Zhang, H. Complete mitochondrial genome of Benthodytes marianensis (Holothuroidea: Elasipodida: Psychropotidae): Insight into deep sea adaptation in the sea cucumber. PLoS ONE 2018, 13, e0208051. [Google Scholar] [CrossRef]
- Kim, H.-J.; Han, J.; Kim, B.-J.; Lee, K.-W.; Hyeong, K.; Choi, Y.-U. New records of two deep-sea eels collected from the Western Pacific Ocean based on COI and 16S rRNA genes. Mol. Biol. Rep. 2021, 48, 5795–5801. [Google Scholar] [CrossRef]
- Han, J.; Kim, H.-J.; Kim, B.-J.; Hyoung, K.; Noh, C.; Choi, Y.-U. New record of Grey cutthroat, Synaphoranchus affinis (Anguilliformes: Synaphobranchidae) from off the east Mariana basin, western Pacific Ocean. J. Mar. Sci. Eng. 2022, 10, 1567. [Google Scholar] [CrossRef]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.J.; Biscoito, M. New records of Synaphobranchus (Anguilliformes, Synaphobranchidae) from the Azores, (eastern Atlantic Ocean). Cybium 2007, 31, 391–392. [Google Scholar]



{kind=link}
{kind=link}
| Gene | Strand | Position | Length (bp) | Start | Stop | Intergenic Region * |
|---|---|---|---|---|---|---|
| trnP | H | 624–693 | 70 | - | - | 5 |
| trnT | L | 699–771 | 73 | - | - | 2 |
| CYTB | L | 774–1913 | 1140 | ATG | TAA | 4 |
| trnE | H | 1918–1986 | 69 | - | - | 1 |
| ND6 | H | 1988–2509 | 522 | ATG | TAG | −4 |
| ND5 | L | 2506–4344 | 1839 | ATG | TAA | 0 |
| trnL | L | 4345–4417 | 73 | - | - | 0 |
| trnS | L | 4418–4486 | 69 | - | - | 0 |
| trnH | L | 4487–4555 | 69 | - | 0 | |
| ND4 | L | 4556–5936 | 1381 | ATG | TAT | −7 |
| ND4L | L | 5930–6226 | 297 | ATG | TAA | 0 |
| trnR | L | 6227–6296 | 70 | - | - | −2 |
| ND3 | L | 6295–6645 | 351 | ATG | TAG | 0 |
| trnG | L | 6646–6717 | 72 | - | - | −1 |
| COX3 | L | 6717–7502 | 786 | ATG | TAA | −1 |
| ATP6 | L | 7502–8185 | 684 | ATG | TAA | −10 |
| ATP8 | L | 8176–8343 | 168 | ATG | TAA | 1 |
| trnK | L | 8345–8419 | 75 | - | - | −14 |
| COX2 | L | 8406–9110 | 705 | ATG | TAA | 6 |
| trnD | L | 9117–9186 | 70 | - | - | 5 |
| trnS | H | 9192–9262 | 72 | - | - | −11 |
| COX1 | L | 9252–10,845 | 1594 | GTG | GAA | 1 |
| trnY | H | 10,847–10,917 | 71 | - | - | 1 |
| trnC | H | 10,918–10,982 | 65 | - | - | −73 |
| trnN | H | 11,020–11,092 | 73 | - | - | 2 |
| trnA | H | 11,095–11,163 | 69 | - | - | 3 |
| trnW | L | 11,167–11,238 | 72 | - | - | −2 |
| ND2 | L | 11,237–12,283 | 1047 | ATG | TAG | 0 |
| trnM | L | 12,284–12,353 | 70 | - | - | −1 |
| trnQ | H | 12,353–12,423 | 71 | - | - | 0 |
| trnI | L | 12,424–12,492 | 69 | - | - | 11 |
| ND1 | L | 12,504–13,472 | 969 | ATG | TAG | 0 |
| trnL | L | 13,473–13,547 | 75 | - | - | 0 |
| l-rRNA | L | 13,548–15,259 | 1712 | - | - | 0 |
| trnV | L | 15,260–15,330 | 71 | - | - | 1 |
| s-rRNA | L | 15,332–16,289 | 958 | - | - | 0 |
| trnF | L | 16,290–16,360 | 71 | - | - | - |
| Gene | Strand | Position | Length (bp) | Start | Stop | Intergenic Region * |
|---|---|---|---|---|---|---|
| trnF | H | 342–413 | 71 | - | - | 0 |
| s-rRNA | H | 414–1371 | 959 | - | - | 1 |
| trnV | H | 1373–1442 | 70 | - | - | 0 |
| l-rRNA | H | 1443–3152 | 1710 | - | - | 0 |
| trnL | H | 3153–3227 | 75 | - | - | 0 |
| ND1 | H | 3228–4197 | 970 | ATG | AGT | 9 |
| trnI | H | 4207–4275 | 69 | - | - | 0 |
| trnQ | L | 4276–4346 | 71 | - | - | −1 |
| trnM | H | 4346–4415 | 70 | - | - | 0 |
| ND2 | H | 4416–5462 | 1047 | ATG | TAG | −2 |
| trnW | H | 5461–5532 | 72 | - | - | 3 |
| trnA | L | 5536–5604 | 69 | - | - | 2 |
| trnN | L | 5607–5679 | 73 | - | - | 35 |
| trnC | L | 5715–5779 | 65 | - | - | 0 |
| trnY | L | 5780–5850 | 71 | - | - | 1 |
| COX1 | H | 5852–7443 | 1595 | GTG | AAG | −9 |
| trnS | L | 7435–7505 | 71 | - | 5 | |
| trnD | H | 7511–7580 | 70 | - | 6 | |
| COX2 | H | 7587–8283 | 697 | ATG | TAA | −6 |
| trnK | H | 8278–8352 | 75 | - | - | 1 |
| ATP8 | H | 8354–8521 | 168 | ATG | TAA | −10 |
| ATP6 | H | 8512–9195 | 684 | ATG | TAA | −1 |
| COX3 | H | 9195–9980 | 786 | ATG | TAA | −1 |
| trnG | H | 9980–10,051 | 72 | - | - | 0 |
| ND3 | H | 10,052–10,402 | 351 | ATG | TAG | −2 |
| trnR | H | 10,401–10,470 | 70 | - | - | 0 |
| ND4L | H | 10,471–10,767 | 297 | ATG | TAA | −7 |
| ND4 | H | 10,761–12,141 | 1381 | ATG | TAT | 0 |
| trnH | H | 12,142–12,210 | 69 | - | - | 0 |
| trnS | H | 12,211–12,279 | 69 | - | - | 0 |
| trnL | H | 12,280–12,352 | 73 | - | - | 0 |
| ND5 | H | 12,353–14,191 | 1839 | ATG | TAA | −4 |
| ND6 | L | 14,188–14,710 | 522 | ATG | TAG | 0 |
| trnE | L | 14,711–14,779 | 69 | - | - | 4 |
| CYTB | H | 14,784–15,923 | 1140 | ATG | TAG | 2 |
| trnT | H | 15,926–15,998 | 73 | - | - | 5 |
| trnP | L | 16,004–16,073 | 70 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.; Kim, H.-J.; Lee, K.-W.; Choi, Y.-U. Complete Mitochondrial DNA Genomes of Deep-Sea Eels Synaphobranchus brevidorsalis and S. affinis and New Record of S. brevidorsalis from the East Mariana Basin. J. Mar. Sci. Eng. 2023, 11, 860. https://doi.org/10.3390/jmse11040860
Han J, Kim H-J, Lee K-W, Choi Y-U. Complete Mitochondrial DNA Genomes of Deep-Sea Eels Synaphobranchus brevidorsalis and S. affinis and New Record of S. brevidorsalis from the East Mariana Basin. Journal of Marine Science and Engineering. 2023; 11(4):860. https://doi.org/10.3390/jmse11040860
Chicago/Turabian StyleHan, Jeonghoon, Han-Jun Kim, Kyun-Woo Lee, and Young-Ung Choi. 2023. "Complete Mitochondrial DNA Genomes of Deep-Sea Eels Synaphobranchus brevidorsalis and S. affinis and New Record of S. brevidorsalis from the East Mariana Basin" Journal of Marine Science and Engineering 11, no. 4: 860. https://doi.org/10.3390/jmse11040860