
Plankton Genes and Extracellular Organic Substances in the Ocean


1
State Key Laboratory of Marine Ecology and Envronmental Sciences, Chinese Academy of Sciences Institute of Oceanology, 7 Nanhai Road, Qingdao 266071, China
2
Agency for Consultation and Research in Oceanography, 19320 La Roche Canillac, France
J. Mar. Sci. Eng. 2023, 11(4), 783; https://doi.org/10.3390/jmse11040783
Submission received: 16 February 2023
/
Revised: 27 March 2023
/
Accepted: 28 March 2023
/
Published: 4 April 2023
(This article belongs to the Special Issue Marine Phytoplankton and Their Evolution)
Abstract
:Dissolved organic matter (DOM) in the ocean represents about 662 billion tons of C, 200 times more than the living biomass. It is produced mainly by microbial primary production. The largest fraction of this DOM is old (>weeks to months) and both chemically and biologically recalcitrant. The remainder is young (seconds to weeks), more labile and surface active. Part of the latter fraction changes the rheological properties in the bulk phase of the water and at interfaces including the sea surface microlayer (SML). In order of abundance, this DOM consists of sugars, amino acids, fatty acids and nucleic acids, often incorporated into complex polymers. The DOM molecules are produced by microbial genes, and are further modified by enzymes themselves produced by genes. The properties of ocean water and its interfaces as well as biogeochemical fluxes may thus be modified by ocean plankton genes. These fluxes influence ocean and atmospheric climate, which in return acts on the biota. Viral infection may furthermore modify prokaryotic and eukaryotic genes and their expression. Therefore, the ocean plankton genomes and the fluxes and climates they influence may be subject to Darwinian-type selection. Research programs need to integrate ocean ecology, rheology, biogeochemistry and genomics, to find the associations among them.
1. Introduction
At 662 billion tons of carbon, marine dissolved organic matter (DOM) holds >200 times as much organic matter (OM) as the living marine biomass and around 100 times more than the dead particulate organic carbon [1]. Marine (DOM) contains as much carbon as the Earth’s atmosphere, and represents a critical component of the global carbon cycle [2]. In the ocean, this DOM is primarily produced by eukaryotic and prokaryotic phytoplankton, with some macroalgae, that reduce CO2 to OM, using sunlight as energy source. The molecular characteristics of the proteins produced are controlled by the organisms’ genes. Further proteins, and other OM, are then manufactured using enzymes, which also are proteins. This OM consists of particulate matter (POM), mainly the solid parts plankton organisms, and DOM that is secreted by phytoplankton and lost from cells during lysis. This paper is not concerned with the POM, treating only the DOM as well as mucus consisting of exopolymeric substances (EPS). Ocean DOM can be usefully characterized in two ways.
The first way to categorize DOM is by chemical composition. The most abundant primary component of DOM is sugars, followed in decreasing order by amino acids, fatty acids and nucleic acid bases, which correspond for EPS to polysaccharides, proteins, lipids and nucleic acids (DNA and RNA). Much of the EPS consists of polymer molecules bearing a variety of functional groups, which determine their physicochemical properties and roles in the ecosystem.
The second way to categorize DOM is by age. The largest fraction of DOM is weeks to hundreds or thousands of years old and has the highest proportion of DOM recalcitrant to biological breakdown [1,3], as well as showing little surfactant or rheological activity [4]. The second, smaller, fraction of DOM is seconds to days or weeks old. It is thus biologically and chemically labile [5]. Some of it is highly surface-active (hydrophilic, amphiphilic or hydrophobic) and some of it adds viscoelastic properties to the water, thus thickening it [4]. This category of DOM is relatively the most abundant in the photic zone [5], particularly in certain harmful algae blooms and mucus events [4]. It also includes most of the signaling, pheromone and toxin molecules [6], that may modulate ecosystem structure [7] and bidirectional vertical fluxes, of OM within the water column [8] and of matter and energy across the ocean–atmosphere interface [9,10].
Given that the allochthonous ocean OM is produced by marine organisms’ genes, and that some of this OM changes the physical properties of bulk ocean water, as well as its interfaces and fluxes across them, it follows that these properties and fluxes are partly under genetic control, and thus subject to natural selection and evolution [11,12].
Rapid progress is being made cataloguing the genes of the pelagic ecosystem, as well as discovering both their roles in producing proteins, and ultimately, via enzymes, of polysaccharides and lipids. Moreover, as Biddanda et al. [13] pointed out, viruses “flip” (i.e., change and transfer) the genes in and between prokaryotic microbes they infect [14], with the outcomes likely reverberating through the ocean microbial community. Progress is also being made in categorizing the rheological properties of ocean waters and soft polymeric structures, as well as how these properties and structures modulate processes and fluxes. The terrestrial/atmospheric and submarine climates are currently changing [15], which is stressing and altering the biota, including humans.
The first aim of this review is to provide a broad overview of the dissolved organic carbon (DOC) pool, including gels and aggregates, along with their surface-active and rheological properties, shape the oceanscape of biogeochemically important OM, with insights into related subtopics such as the genes responsible for microbial physiology and its products. The second aim is to suggest how expression of plankton genes may thereby promote biorheological changes that influence biogeochemical processes. The third aim is to point out the roles of gene expression in crafting signaling molecules, including toxins, which provide control and structure within ocean ecosystems. The fourth aim is to help build a bridge between ocean science and rheology, and to suggest how collaborative research programs including genomics, ocean rheology, biological signaling studies, toxicology and flux studies can be set up to understand how genes control internal ocean processes, and their interaction with the atmosphere. As an added bonus, such studies are likely to throw up commercially exploitable new natural bioactive products.
A sketch of these ideas is shown in Figure 1.
2. Inorganic Matter (IM) and Organic Matter (OM) in the Oceans
2.1. Sources of Available N and Fe
2.2. Origins and Classes of Allochthonous IM and OM
The main origins of dissolved IM (DIM) are soluble salts associated with riverine and terrestrial diffuse sources. They include nutrient salts, notably nitrate, nitrite, ammonia and urea, dissolved inorganic phosphate and dissolved silicate. Anthropogenic sources have increased, not only of nitrogenous nutrients and phosphate, but also of silicate due to disruption of soils and coastal development. The current estimated annual global budget for coastal waters (depth ≤ 200 m) [17] is shown in Table 1 for marine-reactive N. Further important sources and sinks of OC and reactive N in the ocean are atmospheric deposition, sedimentary burial [18].
2.3. Origins and Classes of Autochthonous OM
Most OM in the ocean is produced autochthonously. Ocean Phytoplankton and bacteria sensu lato produce a wide variety of organic molecules. They result from direct extracellular secretion by living cells, as well as leakage from lysed and predated cells as dissolved OM (DOM), normally defined as that passing through a 0.2 µm filter, as well as particulate OM (POM), which is retained by a 0.5 µm filter [20]. Between these extremes lies colloidal OM (COM). In order of abundance, the DOM and COM comprise sugars, amino-acids, fatty acids and nucleic acid bases, polymerized to varying degrees into carbohydrates, proteins, lipids and nucleic acids, respectively, along with complex molecules bearing different radical groups.
2.4. Autochthonous Particulate Organic Matter (POM)
POM produced within the pelagic ocean ecosystem consists principally of living cells and dead remains of cells. These particles act as mechanically solid surfaces allowing colonization by bacteria and protists such as ciliates. These particles also become included in marine aggregates, together with living bacteria and protists, within the more-or-less gluey polymeric matrix of COM. POM is believed to be negatively buoyant in general, and to act, together with autochthonous and allochthonous PIM to ballast marine organic aggregates, thus increasing downward organic flux [8]. More work is required on the functional density and sinking behavior of marine POM.
The relationships between composition, measured density and sinking/rising rates of both non-living and living POM have been subject to much research [8,21,22,23]; this has given rise to varied and confusing results. Largely, this may arise from the Derjaguin-Landau-Verwey-Overbeek assumption (DLVO), taught almost universally in engineering textbooks during the 20th century, that a quasi-universal non-stick non-slip interface could be assumed to exist between water and solid surfaces. Recent research on fluid dynamics at sub-mm and particularly sub-µm length scales, has shown very considerable departure from DLVO between even Newtonian liquids, such as water, and solid surfaces of different qualities, such as rough, smooth, hydrophobic or hydrophilic [24,25,26]. Moreover, the surfaces of POM and even PIM adsorb a covering of OM, which may show varied and complex surface properties. Furthermore, living cells, including those in aggregates and biofilms, manage their surface properties through electric fields, partly by means of glycocalyxes.
2.5. Autochthonous Dissolved Organic Matter (DOM)
Old DOM is mostly refractory (rDOM), whereas DOM newly produced by phytoplankton (pDOM) tends to be highly reactive and labile.
Most DOM in surface, mesopelagic and deep waters is old rDOM. In laboratory studies, Shen and Benner [20] found that, over a time scale of 180 days, about 6–7% of DOM in surface water (50 and 100 m), 1–3% in mesopelagic water (300 m and 750 m) and 0% in deep pelagic water (1500 m) was removed by microbial degradation. The authors found that the amount of degradation depended on the depth origin of the OM, not on that of the microbial community.
pDOM was sampled by filtering (0.7 µm) an in situ bloom was found to be utilized 50–75% within 7 d, and 76–94% within 180 d. By contrast, microbial utilization of DOM from 1500 m was not measurable even after 180 d: it consisted all of rDOM. Spectrophotometry at wavelengths from 250 to 750 nm of both pDOM and rDOM revealed that pDOM showed two absorbance shoulders, at 250–265 nm and at 300–350 nm because of chromophore molecules or groups, but rDOM did not. These two wavelength ranges correspond to labile compounds, including amino-acids and mycosporine-like amino-acids, respectively. Furthermore, elemental analysis revealed that, whereas pDOM had a C/N value, 6.2, similar to the Redfield ration, the corresponding value for rDOM was 36.2, confirming that microbes utilized N-rich DOM referentially, and thus caused the remaining DOM to become depleted in N [20], and thus probably in protein.
3. DOM with Signaling and Allelopathic Functions
Many algal and protist species produce allelopathic compounds. The raphidophyte, Heterosigma akashiwo, produces high-molecular mass (>1 MDa) allelopathic polysaccharide–protein complexes that inhibit its competitor, the diatom Skeletonema costatum (now S. marinoi), by binding to the latter’s cell surface. The authors suggested that viscoelastic, colloidal and/or selectively adhesive of these and other allelopathic polysaccharide–protein complexes (APPCs). The Skeletonema in turn were found to produce several fractions (separated by solid phase extraction) that allelopathically inhibited the Heterosigma in a dose-related manner [7].
At bloom concentrations, the diatoms, Skeletonema costatum, Chaetoceros danicus and Thalassiosira decipiens, all slowed the swimming of the dinoflagellate, Cochlodinium (now Margalefidinium) polykrikoides [27], and the authors suggested that this might have impaired vertical migration of the latter and hence its ability to form blooms. The nature of the allelopathic agent(s), however, was not determined.
Axenically cultured Margalefidinium polykrikoides was found to produce copious mucus, giving average yields of crude polysaccharide of 26 mg/L of culture medium., which on hydrolysis yielded mannose, galactose, glucose and uronic acid together with sulphate groups (7–8% w/w S). The purified polysaccharide inhibited 11 virus strains out of 15 tested at concentrations of 0.8 to 25 mg/L. This abundant occurrence of sulfur-rich molecules is compatible with mucous gel formation by inter-group disulfide links [28].
The first diatom pheromone was identified by Gillard et al. [29]; Pheromones are defined as molecules involved in intraspecific signaling, notably facilitating sexual encounter, whereas those used for interspecific communication are allelochemicals [30].
Many algal pheromones are organic acids or alcohols, whereas some are also proteins or glycoproteins [30]. For example, in the freshwater colonial heterothallic alga, Volvox carteri f. nagariensis, that survives drying out as dormant zygotes, the end of drought induces male colonies to secrete a pheromone inducer that is a large-molecular-mass glycoprotein, and that affects both male and female colonies. This pheromone inducer acts at a concentration of only 10−16 M. It is produced by somatic cells, but initiates gametogenesis by male and female cells. The gene that encodes the pheromone has been discovered [31] and the corresponding protein contains 208 amino acids with a molecular mass of 22 kDa [30,32].
Much laboratory work has been carried out on the influence of bacteria on phytoplankton biosynthesis of natural products [6]. The results obtained may give general clues to the production and effects of signaling compounds in the ocean, as well as the roles of these compounds in structuring ecosystems.
Particularly since the 2010s, the emphasis of work has evolved from allelopathic interactions to intraspecific signaling [33]. Recent work has also targeted phytoplankton chemical defenses, particularly where the molecules involved, e.g., paralytic shellfish toxins (PSTs) or amnesiac shellfish toxins (ASTs), can impact human health [20]. The use of –omics (metabolomics, proteomics and transcriptomics) is revealing mechanisms of biological response to many chemical signals and cues. Outside the area of compounds showing potential for medical or commercial innovation, characterization of the multitude of often unstable and very dilute compounds responsible for chemical mediation of pelagic interactions may remain impracticable [6]. Studies of molecules that modify turbulence, diffusion and binding processes, notably those produced in harmful algal blooms [34,35], in organic aggregates [36], in biofilms and interaction of such molecules with pollutants [37] may also benefit from investigation using omics (See Section 17).
A study by Poulson-Ellestad et al. [38] of the allelopathic effects of the toxic dinoflagellate, Karenia breve, on 9 species of planktonic diatoms typically present in the areas where K. breve blooms in the Gulf of Mexico, showed only weak stimulatory or inhibitory effects on the diatoms, leading the authors to suggest that allelopathic effects could not have been useful to K. breve at the start of the bloom, but could be useful in maintaining blooms once they were established. A second study by Poulson-Ellestad et al. [39] on the allelopathic effects of K. breve on two different diatoms, one of the species, Thalassiosira pseudonana, turned out to be the more susceptible. That K. breve affected nutrient limitation of T. pseudonana was considered unlikely as concentrations were non-limiting. Metabolomic and proteomic investigation using gene ontology categories for a long suite of proteins associated with metabolic pathways and functions in T. pseudonana revealed that many processes, including those involved in cellular carbohydrate metabolism, were strongly stimulated when exposed to K. breve, whereas others, including photosynthesis and chromatin assembly, were strongly inhibited. That T. pseudonana was susceptible to K. breve may be associated with the finding that the two species do not co-occur in nature. Although K. breve produces the toxin, brevetoxin, it is not known if this was an allelopathic agent acting against T. pseudonana.
In a study from the Baltic, Hakenen et al. [40] tested 10 µm filtrate of cultures of 10 local strains of a recurring dinoflagellate, Alexandrium ostenfeldii on different flagellates. At characteristic bloom concentrations, all the strains caused allelopathic effects on the cryptophyte, Rhodomonas salina and the dinoflagellates, Kryptoperidinium foliaceum, Levanderina fissa and Heterocapsa triquetra. All the strains of A. ostenfeldii showed allelopathic effects on all the target species. K. foliaceum reacted by encysting, but excysted again within 24 h, a proportion of the L. fissa cells lysed, and H. triquetra shed their thecae and became immotile, but within 24 h they had mostly recovered. The chemical nature of the allelopathic agent was not investigated.
4. Molecules in Intraspecific and Interspecific Signaling
Pheromones, secreted by copepods, such as Temora longicornis and Eurytemora affinis, change the swimming behavior of conspecifics of the opposite sex (both males and females). This is consistent with evolved use of pheromones to optimize encounters that may lead to mating [41].
Another use of intraspecific signaling is for communication in biofilms. Bacteria, Vibrio cholerae, cooperate to protect themselves and each other against predatory amoebae, Acanthamoeba casalanii, using vibriopolysaccharide, an extracellular matrix of proteins, nucleic acids, and sugars [42], which are in part controlled by the quorum sensing (QS) regulator, HapR [43]. The genes, vpsR and vpsT, were shown to confer enhanced resistance to amoeboid grazing, since grazing was enhanced in knockdown mutants. Further reduction in grazing resistance occurred when QS was interrupted by knocking out the hapR regulator [43].
As well as signaling by DMS (See Section 5), selection of bacterial prey by the ascidian tunicate, Microcosmus exasperatus, may depend on surface molecules of the bacteria more than on their shape or size. Marine picocyanobacteria, notably Synecococcus and Prochorococcus, have sticky, hydrophobic coatings, and are retained with relatively high efficiency by the feeding nets of the ascidian. In contrast, Pelagibacter ubique, and other species of the SAR11 guild, have a non-sticky, non-hydrophobic coatings, which allows them to slip through the mucous feeding nets and thus show low retention by this tunicate. While such a coating may reduce adhesion to nutrient-rich organic particles, it may confer resistance to grazing by mucous-net suspension feeders [44].
Rosa et al. [45] carried out a study on feeding selection by two lamellibranch mollusks, Mytilus edulis and Crassostrea virginica, of 10 species of nanoflagellates. The selection and sorting structures in these mollusks are complex and dynamic, but like in ascidians, mucus is heavily involved. Amongst the nanoflagellates studied, only Pavlova lutheri had a wettable, hydrophilic surface, and its retention by the mollusks was also the least. The wettability (hydrophilicity) and surface charge of surface of many cells is largely modulated by lectins in the glycolcalyx. Lectins are widely distributed glycoproteins, with important functions in molecular recognition.
5. Dimethylsulfide (DMS) in Signaling and Structuring Consortia
DMS is responsible for the characteristic smell of algal culture rooms. It is secreted by most dinoflagellates, including the hosts of the parasitic dinoflagellate, Parvilucifera sinerae. Garcés et al. [46] found that concentrations of 270–300 nM DMS end dormancy in P. sinerae. Correspondingly, all dinoflagellate species that produce DMS in sufficient concentration cause P. sinerae to wake up, but only some of these species become parasitized. Garcés et al. [46] suggested that this was a rare demonstrated example of the now classical idea of “watery arms race[s]” in the ocean [47]. Are watery arms races so rare, however? Schwartz et al. [33] review many examples of models as well as laboratory and field studies supporting complex dynamics between competing species, where one or more species deploys allelopathic compounds to inhibit or even lethally poison competitors. (See Section 3, Section 4 and Section 7). Outcomes can be the elimination of one competitor or else co-existence, depending on environmental conditions such as nutrient concentrations or even allelopathically influenced nutrient uptake dynamics.
Signaling molecules are not all aggressive, but may be mutually helpful. Studies of mutualism in the sea have long been concentrated on bitrophic interactions and it was widely assumed that more complex mutualistic systems in the ocean were rare [33]. Recently, however, Savoca and Nevitt [48] produced evidence for tritrophic mutualism in the Southern Ocean, where phytoplankton are grazed by crustaceans. This phytoplankton releases copious DMS, which in turn attracts procelliiform seabirds that feed on these primary consumers, thereby reducing grazing pressure on the phytoplankton. In a similar vein, Amo et al. [49] found experimentally that the nestlings of the krill-eating chinstrap penguin, Pygoscelis antarctica, are attracted to the scent of DMS. Thus, it is likely that DMS released by zooplankton grazing acts as a signaling molecule via the penguins within a consortium at length scales at least up to 100s of m. In this consortium, cohesion was achieved by the penguins’ behavioral attraction to DMS.
6. Consortia Structured by Rheological Properties, including Stickiness, of Polymers
In many other consortia, such as biofilms [50,51,52], lake and marine organic aggregates [53,54], however, cohesion is achieved by physical processes such as stickiness [55], gelling [56] or increased viscosity [57], mediated by exopolymeric substances (EPS). Schwartz et al. [33] suggested that consortia with more elements than 3 are likely to exist, but that the number of degrees of freedom would make their existence difficult to prove.
Aron et al. [58] have introduced Global Natural Product Social Molecular Networking (GNPS) as a tool for analyzing the infrastructure of molecular datasets and curating the data in a public database in the style of GenBase. GNPS may have the potential for investigating diverse marine consortia and other biotopes, and ultimately the whole ocean. It may be desirable to accompany such a molecular database with another on associated measured rheological properties in the ocean environment at different scales.
7. Prey-Capture and Predator-Avoidance
Chemical cues exuded by prey or occurring on their surface can be used by predators to locate and choose their prey. The copepod, Temora longicornis, feeds on sinking marine snow (MS) particles, which leave a chemical trail behind them [59]. The detection of tunicate exoskeletons (as an experimental proxy for MS) produced doubling of swimming velocity towards the falling particle; the authors suggested that detection must have been chemical, as distances were too great for hydromechanical detection. They also drew parallels with how pheromones secreted by the copepods, T. longicornis and Eurytemora affinis, change the swimming behavior of the opposite sex [41] (See Section 4).
8. Predator–Prey Interactions
Schwartz et al. [33] reviewed the effects of chemical signaling in protist–protist and copepod–protist pairs. Many different reactions have been reported, both on predator–prey reactions and on the swimming behavior of both prey and predators. Brown et al. [6] also reviewed the effects of signaling molecules on predator–prey pairs, particularly invoking the effects of phytoplankton toxins, acting either to harm potential predators or as aids in capturing prey.
Ianora and Miralto [60] review the short-chain polyunsaturated aldehydes (PUAs), secreted when copepods, such as Calanus finmarchicus, or Temora longicornis, graze on diatoms such as Thalassiosira rotula, T. pseudonana, Phaeodactylum tricornutum, Skeletonema marinoi, Chaetoceros affinis, C. decipiens or C. socialis, and as a result produce deformed and thus unviable larvae. Some species of the planktonic diatom genus, Pseudo-nitzschia, such as P. seriata, internally produce and partially release a nerve toxin that produces Amnesic Shellfish Poisoning (ASP) in humans, as well as behavioral changes in marine vertebrates and some invertebrates. Tammilehto et al. [61] found that P. seriata releases the secondary metabolite, domoic acid (DA), when damaged through grazing by copepods, such as C. hyperboreus and C. finmarchicus. The same copepods, however, were highly resistant to the DA [62], which suggested to Brown et al. [6] that some populations of copepods may have evolved resistance to DA.
Prince et al. [63]) showed that, as well as acting as a neurotoxin, DA, produced by Pseudo-nitzschia delicatissima, inhibits growth of another diatom, Skeletonema marinoi, only slightly under low (perhaps limiting) concentrations of Fe, (0.18 µmol L−1), but much more strongly when Fe was replete (~18 µmol L−1) for diatom growth. DA also inhibited growth of S. marinoi, while slightly stimulating growth by P. delicatissima. In the experimental work of Prince et al. [63], it was not clear whether P. delicatissima actually produced DA, like its congener, P. seriata. If it does, the DA might then play two roles: firstly, favoring diatoms that produce it; secondly, inhibiting competing phytoplankton possibly by scavenging Fe, and by poisoning potential predatory zooplankton.
9. Mucus Trophic Structures (“Mucus Traps”)
Rather like terrestrial web-weaving spiders, many marine organisms feed using structures of polymeric mucus that entrap passing prey by more or less sticky polymers that in some cases are toxic as well, killing, or just immobilizing the prey.
Some multi-cellular zooplankters release mucus to the environment, and appear to provide manure and “garden” their prey before eating it. The harpacticoid copepod, Diarthrodes nobilis, secretes mucus fibers through vents in its carapace. It then weaves these fibers to produce an enmeshing capsule, to which it adds its own feces. This allows prokaryotes to multiply on the capsules, which are then ingested by the copepod [64]. Whereas it is unclear how much of the mucus remains in the environment, the authors suggest this “gardening”, on “mucus-traps” is analogous to procedures previously described in marine nematodes [65].
Mucus traps produced by protists are structured in different ways. For example, the dinoflagellate genus, Dinophysis, produces various toxins, including okadaic acid, pectenotoxin (PTX2) and dinophysistoxin 1b (DTX1b) but from calculations of the amount secreted extracellularly, Nielsen et al. [66] calculated that field concentrations would have been too low to support the idea that these toxins act as allelopathic agents. Instead, the dinoflagellate, Dinophysis acuminata, uses “mucus threads” to trap its prey, the ciliate, Mesodinium rubrum, which is an obligate part of its mixotrophic life cycle [67,68].
Ostreopsis cf. ovata cells produce collective benthic webs of sticky mucous fibrils associated with the toxins, ovatoxin-a, -b, -c, -d/e and putative palytoxin [69]. Organisms that get stuck or entangled on the webs are then attacked, often by several cells collectively, and devoured.
The planktonic dinoflagellate, Alexandrium pseudogonyaulax, secretes transparent spheroidal mucous traps, which are sticky and entrap small flagellates. The trap often remains attached to the dinoflagellate by its trailing flagellum, then the dinoflagellate engulfs these prey individually, finally abandoning the spent mucus trap [70] (Blossom et al., 2012).
In gradients of viscosity significant at the length scale of cell size, motile cells are expected to be slowed more on their side in the more viscous water, thus being turned to swim into the more viscous region. Zones of high viscosity may act as traps for swimming cells. Such “viscous traps” are sometimes used by heterotrophic and mixotrophic protists to catch prey, and in some cases the protists even lace the high-viscosity zones with lytic toxins [71]. Stehnach et al. [72], however, observed that in gradients of viscosity the chlorophyte, Chlamydomonas, uses viscophobic turning to actually steer their swimming away from zones of higher viscosity. This behavior would allow these flagellates to avoid such viscous traps. It also implies that they are able to sense viscosity and the direction of viscosity gradients, a capability reminiscent of that of diatoms that detect and react to turbulence fields [73]. (See Section 13).
10. Mucus as a Retention Tool
At the ecosystem scale, endosymbiotic algae in the coral polyps, the zooxanthellae, account for most of the reef’s primary production (PP). In the Great Barrier Reef, for example, half of this PP is exuded by the polyps as mucus. This mucus traps organic matter from the water column, settles and carries energy and organic matter to the reef sediments. Dissolved mucus (~50–80% of the total mucus) is filtered through the lagoon sands, where it is quickly (~7%/h) degraded. Undissolved mucus aggregates trap particles, increasing their organic C and organic N by 2 orders of magnitude within 2 h. Currents concentrate these aggregates into the lagoon. Coral mucus thus provides light energy, harvested by the zooxanthellae and trapped particles to the heterotrophic benthic community of the reef [74]. Filtration of DOM and POM by reef sponges may significantly add to the reef-scale trapping of OM [75]. This constitutes a recycling loop that retains energy and nutrients within reef ecosystems, known for their outstandingly high biodiversity and productivity. Evolutionary processes and structures within the community-scale genome of coral reefs deserve investigation.
11. The Roles of Cross-Linked Gels, Rheological Changes and Reactive Oxygen Species in Toxicity to Fish
The sulphated amino acid, cysteine, and its derivative acetyl-N-L-cysteine (NAC) are mucolytic and antioxidant. In human medicine, the mucolytic effect of NAC is based on the presence of the free sulphydryl group (–SH), that opens up disulfide bonds (S–S) of the high-molecular-weight glycoproteins of human mucus, thus reducing the viscosity and elasticity of the mucus. NAC can also lyse DNA in sputum. NAC is also a direct and indirect antioxidant. The direct effect is produced by the free sulphydryl group, which is a source of electron donors that inactivate (i.e., scavenge) reactive oxygen species (ROS). NAC scavenges •NO2, CO3•−, and thiol radicals quickly, but O2•−, H2O2 and peroxynitrite only slowly and O2 or NO not at all [76,77].
Yang and Albright [78] sought treatment for coho salmon, Oncorhynchus kisutch, killed by blooms of the diatom, Chaetoceros concavicornis. This species bears sharp, pointed spines that are believed to irritate the gills and induce them to produce excess mucus. This mucus is a proteinaceous material that includes mucopolysaccharides, with the long, interconnected, fibrous molecules occurring within a gel. The physical properties of mucous secretions are largely determined by the high molecular weight glycoproteins that consist of a protein backbone with many oligosaccharide side chains, often called mucin. The peptide chain of mucin contains some non-glycosylated regions, which contain many cysteine residues. Many mucous glycoproteins are composed of polymerized glycoprotein subunits through the formation of disulfide bonds in the non-glycosylated region of each protein core, probably involving interaction between adjacent cysteine residues, which results in a network of matted molecules. Yang and Albright found that salmon administered L-cysteine ethyl ester (LCEE) in their feed showed increased survival. Because cysteine and its derivatives, NAC and LCEE, can break S=S bonds and thus fluidify mucus, including mucus secreted by gills [79], Yang and Albright concluded that this effect was responsible for reducing mortality. NAC, also included in feed, was additionally found to reduce the toxic effect to fish of cylindrospermopsin, a toxin produced by several harmful cyanobacteria [80].
After finding that gas bubbles were preventing from rising in a bloom of the fish- and invertebrate-killing dinoflagellate, Karenia mikimotoi (also then known as “Gyrodinium aureolum” or Gymnodinium nagasakiense) [81], and measuring the viscoelasticity of cultures of different species of phytoplankton [82,83,84], modelled that K. mikimotoi could slow flow and thus reduce O2 supply, thereby suffocating the fish when present in sufficient concentration. Jenkinson and Arzul [83,85] found that cultures of K. mikimotoi and the fish-killing raphidophyte, Heterosigma akashiwo, slowed flow (relative to that of pure culture medium) through fish gills, but cultures of the harmless haptophyte, Pavlova lutheri, did not. H. akashiwo, furthermore, showed more variable results than either K. mikimotoi or P. lutheri, suggesting that the EPS it produced was more heterogeneous. The relationship between flow rate and hydrostatic pressure difference over the gills suggested that K. mikimotoi and H. akashiwo added significant amounts of gel-like EPS to their ambient milieu, but that P. lutheri did not. In addition, however, K. mikimotoi was found to produce reactive oxygen species (ROS) [83,85] that contribute haemolytic toxicity [86], which was assayed by measuring the toxin’s lysis of red blood cells. K. mikimotoi isolated from European waters (France and Ireland) may be more active rheologically than that from East Asian waters. The term “rheotoxity” was used by Jenkinson and Arzul [85] to mean “toxicity” (i.e., harm) done to organisms by increased viscoelasticity. An allied meaning is locally increasing the concentration of chemical toxins through increases in viscoelasticity that reduce dispersal [79].
Harmful algae other than Karenia mikimotoi that produce conspicuous amounts of mucus, measured increases in viscosity and/or foam include blooms of other Karenia species, particularly K. selliformis [87], the dinoflagellates, Margalefidinium (=Cochlodinium) polykrikoides [88], Ostreopsis cf. ovata [89] and the haptophytes, Phaeocystis globosa [90,91], P. pouchetii [92] and P. antarctica [93].
In July 1985, at Caño Island, close to the Pacific coast of Costa Rica, massive mortality of corals occurred, along with that of many species of fish (including scarids, acanthurids, pomacentrids, tetradontids and balistids), crabs and gastropods [94]. The plankton contained 8.3 × 105 live cells and >3 × 106 cells in total. The dinoflagellates, Margalefidinium catenatum and Gonyaulax monilata (=Alexandrium monilatum) comprised 97% and 1%, respectively. In October and November 1985 at Uva Island, close to the Pacific coast of Panama, about 300 km SE of Caño Island, a red-brown bloom of dinoflagellates lasted several days. Dinoflagellates and viscous foam co-occurred, suggesting to the authors that the former had produced the latter. Pocilliporid corals had bleached. The authors concluded that the mortality of the reef organisms at Caño and Uva Islands was most likely caused by adhering of the mucus and, in the case of polyps, interference by mucus with their expansion, although chemical toxicity and oxygen depletion may also have contributed [94].
Kim et al. [95] investigated toxicity in Margalefidinium polykrikoides and in the raphidophyte, Chattonella marina, using human epithelial carcinoma (HeLa) cells as target. In culture, growth of both species was at first exponential, reaching a plateau phase. After 12 days, in the cultures of M. polykrikoides and C. marina the polysaccharide concentrations had increased to respective concentrations of 47 and only 4 µg/mL glucose equivalent. The M. polykrikoides cultures became noticeably more viscous, whereas the C. marina cultures did not. Nevertheless, because antiviral activity had been previously reported in M. polykrikoides mucus [28], Kim et al. [95] proposed that cytotoxic agents may have contributed to ichthyotoxicity by M. polykrikoides.
During extensive blooms of M. polykrikoides around Oman and Muscat, associated with the deaths of hundreds of tons of fish and shellfish, Al Gheilani et al. [96] reported that whereas strong odors occurred, thought to be caused by methyl sulfide, no toxicity was detected in mouse tests. Scanning electron microscopy, however, showed mucus proliferation, which might have clogged gills, and fish gills also appeared damaged.
Working on M. polykrikoides from South Korea, Lee et al. [97] conversely showed haemolytic activity in both the water-soluble and chloroform-soluble fractions isolated from methanol extracts of M. polykrikoides. Yet, C.S. Kim et al. [98] found that M. polykrikoides, also isolated from South Korea, produced high quantities of reactive oxygen species (ROS), which they suggested was the primary cause of fish mortality, through damage to gills. Kim and Oda [88] investigated the fish-killing mechanisms of Chattonella marina and M. polykrikoides from the Yatsusiro Sea, Japan. Their results suggested that C. marina has an NADPH-dependent superoxide generation system in its glycocalyx. Their results also suggested that C. marina continuously releases H2O2 into the medium during culture, whereas M. polykrikoides may not release H2O2, at least under normal physiological conditions. Their results suggest that continuous accumulation of discharged glycocalyx on the gill surface occurs during C. marina exposure, which may be responsible for the ROS-mediated severe gill tissue damage leading to fish death. Compared to C. marina, the levels of O2− and H2O2 detected in M. polykrikoides were only trace amounts. Both lectins and mucus prepared from fish skin and gills of yellowtail, when administered separately, induced markedly increased production of O2− in C. marina, but not in M. polykrikoides. Further results suggested that the O2− generation system of C. marina is located on the cell surface, whereas only slight evidence of cell-surface generation was shown in M. polykrikoides. Evidence was shown, however, for the production of H2O2 in both species. Cell-free aqueous solutions prepared from both C. marina and M. polykrikoides were tested on HeLa cells as target. After 24 h treatment with 10% final concentration of each extract in α-minimal essential medium containing 10% fetal bovine serum, cytological changes took place on the HeLa cells and colony formation was reduced, whereas corresponding extract from C. marina produced almost no effect. Kim and Oda [88] suggested that the difference between Lee et al.’s [97] findings and their own could have reflected differences in strain characteristics.
Flores-Leñero et al. [99], studying ichthyotoxicity induced by the raphidophyte, Heterosigma akashiwo, in Patagonian fjords, found that ROS and polyunsaturated fatty acid (PUFA) was too weak to explain the fish kills that occurred, and the authors suggested that further studies should explore other fish-killing mechanisms, such as the production of mucus or extracellular polymeric substances (EPS). These results may reflect the conclusions of Yamasaki et al. [100] that the role of PUFAs and ROS in H. akashiwo blooms may be rather part of a suite of non-lethal signaling molecules that controls marine microbial community structure and function. (See Section 1 and Section 3).
Blooms with the presence of either of the dinoflagellates, Gonyaulax fragilis or G. hyalina, have been associated with viscous/slimy water and foam, frequently associated with mass mortality of marine organisms. Such phenomena have been recorded from the Sea of Marmara, Turkey (G. fragilis—foamy mucilage) [92]; the Tasman Bay, New Zealand [101]; the Northern Adriatic (G. fragilis—mucilaginous masses) [102,103]. G. fragilis and G. hyalina may have been confused by some authors, as they are similar, but separate species [104]. Both produce copious mucus from their apical pore, resulting in noticeable mucilage in the field even at cell concentrations as low as several thousand cells per liter.
Riccardi et al. [105] studied the role of G. fragilis in producing mucilage events in the N. Adriatic, as well as sterols as potential lipid biomarkers associated with this mucus. They also extracted DNA from G. fragilis, and using PCR to characterize it, successfully developed a species-specific DNA probe. In culture, moreover, G. fragilis was associated with the 4α-methylsterols. That the association between G. fragilis and specific sterols was found not to be clear in N. Adriatic mucilage-rich field samples was ascribed to rapid decay or transformation of the dinoflagellate cells and of the sterols. However, the authors suggested that in future, rapid genomic detection coupled with identification of phytoplankton cells in the field could be used to investigate their association with lipid biomarkers, such as sterols.
The dinoflagellate, Karlodinium arminger, produces karlotoxins 1, 2, 8 and 9, which have all been implicated in fish kills [6]. This toxin lysed trout gill cells with a LC50 of 125 nM, and it showed a somewhat higher LC50 of 400 nM for the copepod Acartia tonsa, a potential predator. Huge numbers of algal toxins, particularly those harmful to humans, have been revealed in the last 20–30 years (reviewed by refs [6,33]). While further details are outside the present paper’s scope, evaluating the genes associated with these toxins should be given priority.
Species of the dinoflagellate genus, Dinophysis, produce various toxins, including pectenotoxin 2 (PTX2) and dinophysistoxin 1b (DTX1b) [66]. According to Schwartz et al. [33], however, calculated field concentrations would have been too low to allow to support the idea these toxins act as allelopathic agents.
Some mechanochemical aspects of disulfide bonds (cross-links).
- Zhang and Zhang [106], using a mechanochemical technique, single-molecule force spectroscopy with an atomic force microscope (AFM), explored the relation between rate-of-strain and opposing stress in pulling and breaking single molecular bonds in both polysaccharides and proteins. For example, they found breaking forces in the typically algal polysaccharides, λ-, β-, and ι-carrageenan all to be around 500 to 700 pN.
- In contrast, disulfide links in protein were found by Wiita et al. [107] to have a lower breaking strength of around 100 to 200 pN, and they can be weakened by dithiothreitol (DTT). Disulfide links are important as they strengthen both proteins [108,109] and polysaccharides [109] in biological systems by forming cross-links between polymer chains.
12. Mechanisms of Killing Microbes
Direct killing may be the strongest form of signal. Some bacteria produce toxins that are lethally toxic to microalgae. Hu et al. [110] found that algicidal bacterium, CZBC1, is lethal to the cyanobacteria, Oscillatoria chlorina, O. tenuis and O. planctonica, to the extent that the authors have patented culturing these bacteria to control cyanobacteria in aquaculture facilities. The same authors [110] also reviewed the effects of other algicidal bacteria on microalgae, including cyanobacteria. Bacteria of the genera, Cytophaga and Saprospura, contact and lyse dinoflagellates and diatoms. A strain of Pseudomonas putida kills the diatom, Stephanopyxis, by direct alginolysis, but most algicidal bacteria act indirectly by secreting algicidal compounds. Bacillus strain LZH-5 from Lake Taihu, China, acts strongly on Microcystis aeruginosa.
Concerning the size of the signaling (toxic) molecules, the marine bacterium, Bacillus cereus Strain CZBC1, produces an alginolytic compound retained by a 10 kDa filter [110]. On the other hand, Lee et al. [111] found that Pseudoalteromonas Strain A28 produces a serine protease activity responsible for algal lysis, but they also showed DNase activity; the supernatant from Strain A28, which passed through a 10 kDa filter could kill the diatom, Skeletonema costatum. In addition, Hu et al. [110] showed that that Strain CZBC1 lysed O. chlorina and O. tenuis by direct alginolysis, whereas its extracellular products lysed O. planctonica. Various authors cited by Hu et al. [110] showed that algolytic effects of bacteria are concentration-dependent.
13. Quorum Sensing
Quorum sensing (QS) is a density-dependent communicating mechanism that allows organisms to regulate a wide range of important processes and can be inhibited by quorum quenching (QQ), for example, in marine organic aggregates (MOAs) [112] and probably also at larger scales. Falciatore et al. [73] may have weakened Svedrup et al.’s [113] old paradigm that plankton are passive and incapable of resisting physical forces at any scale. Reviewing post-Sverdrup findings that “[some] plankton control buoyancy, local fluid viscosity and life cycles”, Falciatore et al. [73] showed experimentally that diatoms detect and respond to physicochemical changes in their environment using sophisticated perception systems based on changes in cytoplasm concentrations of Ca2+ as a second messenger [114].
Microbes communicate with each other using diffusible molecules such s N-acetylhomosereine lactones (AHL). Communication of the information in a signal requires sensing of the signal and it is not clear whether the different types of sensing use secondary messengers sensu Endo [114]. Nevertheless, these different signals are likely physiological and behavioral cell-density-dependent gene regulators [115], which involve QS and control microbial processes (Figure 1).
Whether in bacteria, algae or metazoans, tighter spatial association will increase intensity of interactions, as well as reducing the distances and, mostly, the times of signal transmission [116], whether by diffusion or by radiation, of information agents such as pheromones, light, sound or other electrical or mechanical signals. Association may be increased (decreased) by attractive (repulsive) behavior or by rheological means, such as increasing (decreasing) the viscosity or the yield stress of the ambient medium. When yield stress is larger than the shear stresses tending to deform the ambient medium, the spatial distribution of particles, such as organisms, in the medium is gelled. Even when the yield stress is less than the ambient mechanical stresses, the increases in viscosity will slow the medium’s deformation and hence dispersion of the particles and molecules.
Increases in mechanical (rheological) properties, such as viscosity [34,57,91,117] of the transmitting medium, will also tend to reduce the length-scales of mechanical or chemical signal transmission. Such modification is largely carried out by secreted EPS, particularly that bearing saccharide groups. Such rheological modification by EPS will therefore also modify the signals that signaling molecules, including toxins, transmit. Such EPS may thus be considered to include “auxiliary signalling molecules”, which participate in niche engineering [118,119,120], also formerly called “physical environmental management” [121].
MOAs may be considered as 3D equivalents of marine biofilms (MBs) [9,122,123]. In MOA- and BF-associated prokaryotes, including the Gram-negative alphabacterium, Paracoccus carotinifaciens, and the gammaproteobacterium, Pantoea ananatis, resistance to viral infection and to protozoan predation is achieved by secretion of various homosereine lactones and ammonium, respectively [112,124,125]. In the bacterium, Vibrio cholerae, living in biofilms, resistance to protozoan grazing is effected by secretion of the metabolite, ammonium, at concentrations of up to 3.5 mM, which was found to reduce concentrations of the protistan grazer, Rhynchomonas nasuta, by >80% [126].
By contrast, other bacteria, including the fish pathogen, Vibrio anguillarum, defend against viral (phage) infection by increasing expression of the ompK gene, which correlates with the degree of cell aggregation, being low in free-living variants [127]. The gene, ompK, produces N-acylhomoserine lactone (AHL) a molecule with QS signaling and many metabolic functions, and there may thus be a link among ompK, AHL and aggregation. So far, however, no causal mechanism among ompK, AHL and aggregation has been demonstrated. Indeed, investigation of which genes are involved in producing enzymes involved in producing sugars and the enzymes catalyzing assembly into polysaccharides, such as EPS, as well as in EPS destruction, either in pro- or eukaryotic aquatic, single-celled organisms, appears to have started only recently [112].
AHLs are probably the most intensively studied class of mediators in cell-density-dependent gene regulation [115,128], and have been found in bacterial biofilms and marine snow, in which it is believed that bacteria, interacting with ambient pressures and constraints, control its form and phenotypic traits [129,130]. In biofouling, surface sensing via AHLs of bacterial biofilms is the initial step in the settling of the macroalga, Ulva [131,132].
Outside of biofilms and MOAs, microalgae, too, may alter behavior in multi-celled organisms. Increases in water viscosity caused by secretion of polymers by the haptophyte, Phaeocystis globosa, were observed to make swimming patterns in the copepod, Temora longicornis, more compact [90], whereas the same species reduced measured feeding rates and filtering rates in Temora stylifera [133].
In bacterially dominated biofilms, reviewed by Karn et al. [52] for their role in promotion or inhibition of corrosion, carbohydrates are generally the most abundant constituents, accounting for 40–95% by mass, while proteins typically contribute 1–60%, lipids 1–40% and nucleic acids 1–10%. The biofilm matrix acts as a recycling center, by preventing the molecular products of live cells from dispersing and becoming lost to the consortium [134]. These products include DNA, which could represent a reservoir of genes for horizontal gene transfer over small distances. Proteins, along with humic substances, might play a role as electron donors or acceptors by forming bacterial pili and nanowires. Modulation of rheological properties in biofilms, such as binding and stabilization, which affect retention of enzymes, may take place mainly by interactions with polysaccharides and proteins in the biofilm in reaction to mechanical forces, including those caused by deformation in the surrounding milieu [134,135].
In a study of light-induced aggregation related to reactive oxygen species (ROS), Sun et al. [136] found that MOA size was positively related to protein/carbohydrate (P/C) ratio. The authors measured MOA size after ultrafiltration through 0.2 µm polycarbonate filters, which are hydrophilic, and polymer particles then re-formed spontaneously in the filtrate. The authors’ results indicated that the inter-protein bonds were stronger than inter-polysaccharide bonds. In this respect, microrheological measurement carried out near phytoplankton cells [57] found that viscosity increased by up to a factor of 2 to 5 at distances of 2 to 5 µm from cell surfaces, typically declining to a factor of 1.2 at 10 µm from the cell.
Like marine organic aggregates (MOAs), sewage sludge organic aggregates (SOAs) consist mainly of bacteria and diverse debris held together by loosely-bound EPS, in a slimy matrix of closely-bound or unbound EPS. The more concentrated nature of SOAs compared to MOAs, together with the need to dewater sludge for economical transport and disposal, drives much research activity on the rheology and surface science of SOAs [137], as well as on ecological chemistry of SOAs and biofilms [52,138]. This activity provides results and expertise with strong potential to inspire and guide research on MOAs.
14. Scales (Granulometry) of Toxicity
The term, toxicity, is usually used in the sense of causing harm by toxic molecules to living organisms. This implies one length scale of the chemical action (nm) and another at that of distance from transmitter to receiver between microorganisms (1–100 µm) or even that between multicellular organisms. Toxic effects of molecules produced by genomes may act at an environmental scale of km to 1000s of km and be more difficult to identify. In both spontaneously aggregated OM [139,140] and biologically produced mucus, fluid flow and often molecular diffusion are reduced, whereas physical density may be increased or decreased [141,142].
Much research is currently under way on the allelopathic effects of polyunsaturated aldehydes (PUAs), which act as signaling compounds [6]. Bartual et al. [143], working with laboratory blooms of the diatom, Thalassiosira rotula, studied the effect of adding PUA (2.5 µM mixture of three PUAs, 2E,4E-heptadienal; 2E,4E-octadienal; 2E,4E-decadienal) on the aggregation of diatom-secreted TEP and associated formation of OAs. They found that the presence of PUA resulted in larger OAs and they therefore suggested that PUA acts as glue, consolidating the OAs. Because larger OAs sink faster as marine snow, the diatom-produced PUA might be contributing to increased vertical organic flux. The physico-chemical mechanisms of how these PUAs stick within TEP, as well as the genes responsible for PUA production, require investigation.
15. Hydrophobicity, Organic Aggregate Size and Rheology
The polymeric secretions of algae and bacteria include polysaccharides, proteins, lipids and nucleic acids. Proteins are amphiphilic: they bear both hydrophilic (polar, wettable) and hydrophobic (non-polar) domains and they are considered to contribute most of the hydrophobicity of EPS. In contrast, polysaccharides are mostly hydrophilic through their polar oxygen groups. An increase in the degree of internal hydrogen bonding, however, can increase their relative hydrophobicity [55]. Klun et al. [144] investigated colloidal OM secreted in culture by a chlorophyte nanoflagellate, Tetraselmis sp., a diatom, Chaetoceros socialis and a dinoflagellate, Prorocentrum minimum, all isolated from the Gulf of Trieste. Table 2 summarizes their results. The OM had been 0.45 µm pre-filtered. It was then ultrafiltered through membranes with nominal molecular-weight cut-off of 5 kDa. The polysaccharide fraction was the highest in the retentate of Tetraselmis sp. (61%), whereas lipids and proteins each accounted for 19%. In the permeate, protein represented the highest portion (41%). The C. socialis retentate and permeate contained the highest polysaccharide levels (63% and 46%, respectively), followed by proteins (22% and 36%) and lipids (14% and 16%). P. minimum retentate and permeate showed very different compositions of secreted OM, with polysaccharide proportions of only 32 % and 25%, respectively, compared with high proportions of proteins (46% and 57%, respectively) and with intermediate proportions of lipids (22% and 17%, respectively). For all three taxa, the proportion of polysaccharides was thus higher in the retentate than in the permeate, whereas the proportion of proteins was higher in the permeate, and for lipids the corresponding situation varied. The overall proportion of lipids found in both the retentates and the permeates was surprisingly high, from 14% to 36%.
16. Molecules, Produced by other Organisms and Associated Bacteria, That Are Toxic and Allelopathic to Phytoplankton
Vidal-Melgosa et al. [145] reported a polysaccharide of still unknown structure, called fucose-containing sulphated polysaccharide (FCSP), which appears to be directly secreted by diatoms. Detected by the monoclonal antibody technique, it was found that abundantly distributed on cell surfaces and spines of the diatom, Chaetoceros socialis, in spring blooms in the Helgoland Bight. It appears to have a complicated structure, and unlike other polysaccharides present, it lasted ≫10 days in laboratory culture. This suggests that, over a certain concentration, this relatively recalcitrant polymer might be very important in aggregating to particulate organic matter (POM), and thus mediating vertical flux [146]. This FCSP is strongly negatively charged, and thus forms part of the complex of acid polysaccharides secreted by phytoplankton as diverse as dinoflagellates [69] and cyanobacteria [147].
Some seaweeds and seagrasses also exert allelopathic action on harmful algae. Laabir et al. [148] showed that the methanolic and aqueous extracts of the seagrasses, Zostera marina and Z. noltii, inhibited the harmful dinoflagellate, Alexandrium catenella. These extracts contained flavinoids and phenolic acids, which were themselves toxic to A. catenella. The authors suggested that phenolic acids were the likely candidates for allelopathic action, and that increases in A. catenella blooms in lagoons and other French waters may have been partly caused by declines in seagrass beds.
Subsequently, Onishi et al. [149] showed that only whole-plant extracts of seagrasses, and not exudates, were algicidal. Two strains of Flavobacteriaceae isolated from biofilms on the seagrass leaves show algicidal activity on Alexandrium tamarense. This suggests that the above-mentioned findings [150] might have been caused by bacteria epiphytic on seagrass leaves rather than to the seagrasses themselves, particularly as Flavobacteriaceae have been shown to be algicidal towards several fish-killing rhaphidophytes, dinoflagellates and diatoms. Similarly in the freshwater Lake Biwa, Japan, the bacterium, Agrobacterium vitis, which colonizes surfaces of leaves of the water plant, Egeria densa, is strongly allelopathic to the harmful cyanobacterium, Microcystis aeruginosa, and an increase over several years of E. densa has coincided with a reduction in blooms of M. aeruginosa [150].
Bacteria in the mucous phycosphere of the diatom, Skeletonema costatum, and of the dinoflagellate, Scrippsiella trochoidea, were found by Yang et al. [151] to be able to lyse their host cells, leading to mass deaths of the hosts. The mechanisms for this lysis were not elucidated.
Research on metabolomics in consortia (see Section 6), particularly in biofilms, is intense in the fields of biocorrosion [52], medicine [152] and sewage sludge [153]. It appears, however, still too rare in the aquatic plankton ecosystem [120]. In such biofilm consortia, progress is being made in semi-automated data mining of intra- and extra- cellular DNA to identify gene sets involved in microbial metabolism and production of different molecules. The software is inspired by homologous data mining of texts on the internet. Thakur et al. [154] have made a promising start on bacterial consortia in biofilms associated with biocorrosion. The technique may be capable of development to mine pro- and eu-karyotic gene sets from ecosystems, such as the world ocean, stored in GenBase. Singh [155] pointed out that in human bodies, as in the ocean, glycoproteins play crucial roles in biological processes, including cell signaling, host–pathogen interaction and disease. Glycoproteomics aims to determine the positions and identities of the complete repertoire of glycans and glycosylated proteins in a given cell or tissue. The roles of glycoproteomics in ocean consortia may be analogous, and the links with genes, as least for key processes, should be established. Whereas the main drivers of Thakur et al.’s [154] study may have been the need to understand and control biocorrosion of steel by sulfur-reducing bacteria, and those of the studies cited in Singh’s [155] mini-review were the need to improve human health, the purposes of mining ocean-derived data on gene sets might be to understand and control bio-geo-physico- chemical fluxes of matter and energy in the oceans, as well as cell–cell signaling and their roles in ecological control [7].
17. Organic Polymers and Gaia
Some biopolymers strengthen organic aggregates, biofilms and volumes of water with increased viscosity [4,138,156]. Biopolymers tend to concentrate at surfaces, particularly in the surface microlayer (SML). Here, they change properties including surface tension and 2D viscoelasticity [157], as well as 3D viscosity [158,159], thus modulating fluxes of of matter and energy within the ocean and at its boundaries, particularly with the atosphere [160]. Because organisms are genetically controlled and they participate in the production, metamorphoses and breakdown of these biomolecules, their genomes influence oceanic and atmospheric physico-chemical conditions. Where they are harmful, this may be considered toxicity at the scale of the Planetary ecosystem, consistent with evolution of Gaia [161,162] by natural selection of its genes [12]. An example of such Planetary scale toxicity, at least from an anthropic point of view, is the current increase in atmospheric and oceanic CO2 and global warming.
18. Polymer Modulation of Fluxes: Discovering the Genomes Involved
Fluxes of matter and energy between the ocean and the atmosphere are functions of physical, chemical and biological parameters. In particular, viruses, bacteria sensu lato and eukaryotic protists are expelled towards the atmosphere by bursting bubbles, and some of them are carried by updrafts and winds from a few meters [163] to thousands of kilometers [164]. The surface charge and hydrophobicity of cells and non-living matter strongly influence the tendency to be expelled [10]. For species in which such expulsion leads to dispersion and greater Darwinian fitness, it may represent a pre-adaptation leading to further evolution of hydrophobic/hydrophilic control of ocean–atmosphere flux. Irrespective of the length, time or hyperspace scales at which such natural selection is driven, it will have effects also over other biological and chemical species (collateral effects) in driving ocean–atmosphere fluxes, which are important for climate control [10,15,160].
Notably in the SML and in biofoam, both important to modulating ocean–atmosphere fluxes of matter and energy, the EPS present shows different characteristics. The different species and their expressed genes that produce the EPS thus determine the different ways in which EPS modulates these fluxes.
19. Vertical Organic Flux of OM: Succinct Examples of Modulation by Polymers
The use of metabolomics and proteomics has already illuminated the mechanisms of biological response to many chemical cues and may be helpful in determining their molecular targets. Overall, the ocean represents a vast source of novel interactions as well as new molecules [33]. As Schwartz [33] pointed out, both models and laboratory experiments are needed to evaluate the roles of toxic, allelopathic and signaling interactions in the pelagic. To these interactions might be added those involving modulation of fluxes, diffusion and fluid deformation.
As mentioned in Section 16, Vidal-Melgosa et al. [145] showed the importance of fucose-containing sulphated polysaccharide (FCSP), which is secreted by diatoms, particularly Chaetoceros socialis, which produces mucilaginous colonies and which dominated the North Sea blooms they worked on. FCSP was far more resistant than the non-sulphated polysaccharide, laminarin, to degradation by Bacteroidea and Gammaproteobacteria. These organisms encode enzymes for laminarin degradation, and they are key degraders of laminarin in the field [165]. Because FCSP is degraded far more slowly than laminarin, Vidal-Melgosa et al. [145] suggested that C. socialis and other diatoms that secrete large amounts of FCSP, are likely among the key species that modulate vertical carbon flux in aggregates, thus contributing to the sequester of CO2 from the atmosphere.
20. Vertical Organic Flux between the Photic Zone and the Deep Ocean: The Bigger Picture
Boyd et al. [166] reviewed carbon sequestration processes within the ocean from the photic zone to deep waters below the permanent pycnocline, which is driven by particle pumps and retarded by conversion of the POC into DOC. The authors identify two main mechanisms of sequestration: the Biological Gravity Pump (BGP) and the Particle Injection Pump (PIP). Four types of PIP are identified: 1—the Mixed Layer Pump, in which suspended/sinking particles are trapped under the pycnocline when it shallows, for example in spring in temperate waters or following deepening of the surface mixed layer during wind events, followed by formation of a new, shallow pycnocline when calm weather follows; 2—the Seasonal Lipid Pump, driven by seasonal migration of lipid-rich copepods; 3—the Mesopelagic Migrant Pump, driven by pelagic fish, krill as well as other mesoplankton and nekton, that undertake diurnal migration from the photic zone, where they feed, to below the pycnocline where they defecate and excrete both POC and DOC, as well as releasing CO2 by respiration; 4—the Eddy Subduction Pump (ESP).
Boyd et al. [166] divide the ESP into: a—the Large-Scale ESP, a 3D advective mechanism that transports particles from the seasonal mixed layer into the interior of the ocean; b—the Frontal-Associated ESP, which subducts particle-rich surface waters on timescales of days and across spatial scales to 1 to 10 km.
The importance of the BGP relative to those of the different PIP mechanisms is still not clearly elucidated because this would require intense and carefully designed sampling in 4D [166]. The currently estimated upper bound of all global PIP is 8.7 PgCyr−1, which is comparable to BGP flux (Table 3).
It has been concluded [166] that POC exported from temperate waters is sequestered for the longest time period, whereas that exported in the Tropics and in Subtropical gyres is sequestered for an intermediate time and the shortest time, respectively. Changes in the physicochemical characteristics of those polymers that stick particles together may therefore have a greater impact on vertical carbon flux and sequestration when they occur in temperate waters than in waters of the Tropics or Subtropical gyres.
“Predicting future changes in ocean carbon sequestration will require a better understanding of the injection versus mineralization processes to the sequestration efficiency given the different environmental sensitivity of these processes” [166]. Such prediction may be helped further by better understanding of the different types, amounts, sources and sinks the polymeric OM produced and metamorphosed by the pelagic microbial community through expression of its genes. Research needs to be targeted to the amounts, chemical species, rheological strengths, hydrophobicity and resistance to degradation of this polymeric OM. The effects of gene expression and evolution in the pelagic community may thus be of great importance to carbon flux and sequestration through their control of OM production, metamorphosis and degradation, as well as of organism behavior and signaling/toxicity.
21. Ocean Foam
Ocean foams, such as whitecaps, help to keep the Earth cool [10]. The fraction of incident solar radiant energy that is reflected is known as albedo. Foam-free ocean surface has a typical albedo of only ~0.05, whereas most foam is white and its albedo is ~0.5. Therefore, when foam is more stabilized by algal DOM the percentage foam cover increases and the oceans reflect more solar energy back into space, reducing solar heating of the Earth [169,170]. The decay of these foams proceeds by the coalescence of its constituent bubbles as the inter-bubble water drains, allowing the surfaces of adjoining bubbles to touch and burst [171]. Draining is retarded and foam lifetime becomes longer if the liquid is a surfactant that binds to bubble surfaces or if it is more viscous, or both. Relative to their duration in waters of low phytoplankton biomass (PB) or low primary production (PP), ocean foam duration is increased in waters of high PP or high PB [172], particularly in the presence of certain specific taxa. Blooms that produce a lot of foam are dominated by dinoflagellates on North Atlantic and Asian coasts [173], including Margalefidinium polykrikoides [174], Karenia spp. blooms dominated by K. selliformis in Russian Pacific waters [87], as well as the haptophyte, Phaeocystis globosa, in the North Sea [175]. Blooms of the fish-killing dinoflagellate, Alexandrium monilatum, on the Pacific coast of Costa Rica have been associated with cream- or beige-colored foam, with the water producing violent itching. This species also produces goniodomines [176,177]. The primary chemistry of the EPS and associated genomes of these organisms, which also frequently increase viscosity and stabilize foam, requires further investigation.
22. Conclusions
Ocean ecosystems that need data mining of pro- and eu-karyotic genomes may include biofilms on marine plastic debris, macroalgae and seagrass, sea-ice structures, marine organic aggregates, the sea-surface microlayer and ultimately the whole ocean. Boundaries of consortia, at all granularities, need to be investigated, particularly the ocean–atmosphere boundary. Because the physical properties of polymers, both proteins and polysaccharides, control water shearing and molecular diffusion, such studies should be combined with rheological studies at appropriate scales.
Fluxes of many types of matter and energy in the oceans are modulated, not only by proteins and lipids, but more especially by different dissolved and particulate polysaccharides. An important key to understanding, and perhaps ultimately controlling, these oceanic fluxes thus lies in exploring associations between these polymers and the genes present and expressed in the planktonic microbial community. Polysaccharides, as well as glycoproteins, glycolipids and some proteins are assembled by enzymes, which are themselves proteins. Different polysaccharides are therefore unlikely to be “mirrored” directly by corresponding genomes. This difficulty has slowed progress in ocean glycomics. Recently, however, enzymes responsible for selective cleavage of specific links in different marine microbial polymers have been linked to their roles in bacteria and diatoms in facilitating or preventing aggregation and de-aggregation of OM and adhering microbes [145]. This exciting avenue needs to be accelerated and combined with quantitative multi-scale sampling and analysis of biomolecules as well as matching sampling of genes (many are already in GenBase), organisms, with the addition of a new database of measured rheological properties in the same environment. Intelligent data-mining of these databases may take advantage of GNPS [58] and techniques already developed for bacterial biofilms involved in corrosion [154] (see Section 6 and Section 18). Expansion of such studies to ocean exploration is likely to vastly improve understanding of ecological and metabolic interactions, as well as fluxes, in the ocean. As a bonus, they are almost certain to reveal commercially exploitable natural products.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This review is not associated with any data.
Acknowledgments
This review has enormously benefited from discussions and exchanges with members of the International Working Group on Rheology, nano- and micro-Fluidics, bioFouling and bioFoam in the Oceans and other natural waters (RheFFFO), as well as with Linda Medlin and Beatriz Reguera. Bopaiah Biddanda and anonymous reviewers greatly improved this review.
Conflicts of Interest
The author declares no conflict of interest.
References
- Hansell, D.A.; Carlson, C.A.; Repeta, D.J.; Schlitzer, R. Dissolved organic matter in the ocean: A controversy stimulates new insights. Oceanography 2009, 22, 202–211. [Google Scholar] [CrossRef] [Green Version]
- McCarren, J.; Becker, J.W.; Repeta, D.J.; Shi, Y.; Young, C.R.; Malmstrom, R.R.; Chisholm, S.W.; DeLong, E.F. Microbial community transcriptomes reveal microbes and metabolic pathways associated with dissolved organic matter turnover in the sea. Proc. Natl. Acad. Sci. USA 2010, 107, 16420–16427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, N.; Herndl, G.J.; Hansell, D.A.; Benner, R.; Kattner, G.; Wilhelm, S.W.; Kirchman, D.L.; Weinbauer, M.G.; Luo, T.; Chen, F.; et al. Microbial production of recalcitrant dissolved organic matter: Long-term carbon storage in the global ocean. Nat. Rev. Genet. 2010, 8, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, I.R.; Sun, X.X.; Seuront, L. Thalassorheology, organic matter and plankton: Towards a more viscous approach in plankton ecology. J. Plankton Res. 2015, 37, 1100–1109. [Google Scholar] [CrossRef] [Green Version]
- Benner, R.; Amon, R.M. The size-reactivity continuum of major bioelements in the ocean. Annu. Rev. Mar. Sci. 2015, 7, 185–205. [Google Scholar] [CrossRef]
- Brown, E.R.; Cepeda, M.R.; Mascuch, S.J.; Poulson-Ellestad, K.L.; Kubanek, J. Chemical ecology of the marine plankton. Nat. Prod. Rep. 2019, 36, 1093–1116. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, Y.; Shikata, T.; Nukata, A.; Ichiki, S.; Nagasoe, S.; Matsubara, T.; Shimasaki, Y.; Nakao, M.; Yamaguchi, K.; Oshima, Y.; et al. Extracellular polysaccharide-protein complexes of a harmful alga mediate the allelopathic control it exerts within the phytoplankton community. ISME J. 2009, 3, 808–817. [Google Scholar] [CrossRef] [Green Version]
- Mari, X.; Passow, U.; Migon, C.; Burd, A.B.; Legendre, L. Transparent exopolymer particles: Effects on carbon cycling in the ocean. Prog. Oceanogr. 2016, 151, 13–37. [Google Scholar] [CrossRef] [Green Version]
- Wurl, O.; Ekau, W.; Landing, W.M.; Zappa, C.J. Sea surface microlayer in a changing ocean—A perspective. Elem. Sci. Anthr. 2017, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, I.R.; Berdalet, E.; Chin, W.-C.; Denis, M.; Ding, H.; Duan, J.; Elias, F.; Emri, I.; Karn, S.K.; Li, Z.; et al. The rôles of plankton and neuston microbial organic matter in climate regulation. J. Plankton Res. 2021, 43, 801–821. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species; Harvard University Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Dawkins, R. The Extended Selfish Gene; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Biddanda, B.; Dila, D.; Weinke, A.; Mancuso, J.; Villar-Argaiz, M.; Medina-Sánchez, J.M.; González-Olalla, J.M.; Carrillo, P. Housekeeping in the hydrosphere: Microbial cooking, cleaning, and control under stress. Life 2021, 11, 152. [Google Scholar] [CrossRef]
- Chiura, H. Generalized gene transfer by virus-like particles from marine bacteria. Aquat. Microb. Ecol. 1997, 13, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. (Eds.) IPCC Summary for Policy Makers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Rosenwasser, S.; Ziv, C.; van Creveld, S.G.; Vardi, A. Virocell metabolism: Metabolic innovations during host–virus interactions in the ocean. Trends Microbiol. 2016, 24, 821–832. [Google Scholar] [CrossRef]
- Liu, X.; Stock, C.A.; Dunne, J.P.; Lee, M.; Shevliakova, E.; Malyshev, S.; Milly, P.C.D. Simulated global coastal ecosystem responses to a half-century increase in river nitrogen loads. Geophys. Res. Lett. 2021, 48, e2021GL094367. [Google Scholar] [CrossRef]
- Hamilton, D.S.; Perron, M.M.; Bond, T.C.; Bowie, A.R.; Buchholz, R.R.; Guieu, C.; Ito, A.; Maenhaut, W.; Myriokefalitakis, S.; Olgun, N.; et al. Earth, Wind, Fire, and Pollution: Aerosol Nutrient Sources and Impacts on Ocean Biogeochemistry. Annu. Rev. Mar. Sci. 2022, 14, 303–330. [Google Scholar] [CrossRef] [PubMed]
- Jickells, T.D.; Buitenhuis, E.; Altieri, K.; Baker, A.R.; Capone, D.; Duce, R.A.; Dentener, F.; Fennel, K.; Kanakidou, M.; LaRoche, J.; et al. A reevaluation of the magnitude and impacts of anthropogenic atmospheric nitrogen inputs on the ocean. Glob. Biogeochem. Cycles 2017, 31, 289–305. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Benner, R. Molecular properties are a primary control on the microbial utilization of dissolved organic matter in the ocean. Limnol. Oceanogr. 2019, 65, 1061–1071. [Google Scholar] [CrossRef] [Green Version]
- Bienfang, P.; Laws, E.; Johnson, W. Phytoplankton sinking rate determination: Technical and theoretical aspects, an improved methodology. J. Exp. Mar. Biol. Ecol. 1977, 30, 283–300. [Google Scholar] [CrossRef]
- Bienfang, P.K. Phytoplankton sinking rates in oligotrophic waters off Hawaii, USA. Mar. Biol. 1980, 61, 69–77. [Google Scholar] [CrossRef]
- Wakeham, S.G.; Lee, C.; Farrington, J.W.; Gagosian, R.B. Biogeochemistry of particulate organic matter in the oceans: Results from sediment trap experiments. Deep. Sea Res. 1984, 31, 509–528. [Google Scholar] [CrossRef]
- Rothstein, J.P. Slip on Superhydrophobic Surfaces. Annu. Rev. Fluid Mech. 2010, 42, 89–109. [Google Scholar] [CrossRef]
- Conlisk, A.T. Essentials of Micro- and Nano-Fluidics; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Jenkinson, I.R.; Sun, J. Drag increase and drag reduction found in phytoplankton and bacterial cultures in laminar flow: Are cell surfaces and EPS producing rheological thickening and a Lotus-leaf Effect? Deep. Sea Res. 2014, 101, 216–230. [Google Scholar] [CrossRef]
- Lim, A.S.; Jeong, H.J.; Jang, T.Y.; Jang, S.H.; Franks, P.J. Inhibition of growth rate and swimming speed of the harmful dinoflagellate Cochlodinium polykrikoides by diatoms: Implications for red tide formation. Harmful Algae 2014, 37, 53–61. [Google Scholar] [CrossRef]
- Hasui, M.; Matsuda, M.; Okutani, K.; Shigeta, S. In vitro antiviral activities of sulfated polysaccharides from a marine microalga (Cochlodinium polykrikoides) against human immunodeficiency virus and other enveloped viruses. Int. J. Biol. Macromol. 1995, 17, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Gillard, J.; Frenkel, J.; Devos, V.; Sabbe, K.; Paul, C.; Rempt, M.; Inzé, D.; Pohnert, G.; Vuylsteke, M.; Vyverman, W. Metabolomics enables the structure elucidation of a diatom sex pheromone. Angew. Chem. Int. Ed. 2012, 52, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, J.; Vyverman, W.; Pohnert, G. Pheromone signaling during sexual reproduction in algae. Plant J. 2014, 79, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Tschochner, H.; Lottspeich, F.; Sumper, M. The sexual inducer of Volvox carteri: Purification, chemical characterization and identification of its gene. EMBO J. 1987, 6, 2203–2207. [Google Scholar] [CrossRef]
- Mages, H.-W.; Tschochner, H.; Sumper, M. The sexual inducer of Volvox carteri: Primary structure deduced from cDNA se-quence. FEBS Lett. 1988, 234, 407–410. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E.R.; Poulin, R.X.; Mojib, N.; Kubanek, J. Chemical ecology of marine plankton. Nat. Prod. Rep. 2016, 33, 843–860. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Sun, J. Rheological properties of natural waters with regard to plankton thin layers. A short review. J. Mar. Syst. 2010, 83, 287–297. [Google Scholar] [CrossRef]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlusich, J.J.P.; Pelletier, E.; Zinger, L.; Lombard, F.; Zingone, A.; Colin, S.; Gasol, J.M.; Dorrell, R.G.; Henry, N.; Scalco, E.; et al. A robust approach to estimate relative phytoplankton cell abundances from metagenomes. Mol. Ecol. Resour. 2022, 23, 16–40. [Google Scholar] [CrossRef] [PubMed]
- Santschi, P.H.; Chin, W.-C.; Quigg, A.; Xu, C.; Kamalanathan, M.; Lin, P.; Shiu, R.-F. Marine gel interactions with hydrophilic and hydrophobic pollutants. Gels 2021, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Poulson-Ellestad, K.; Mcmillan, E.; Montoya, J.P.; Kubanek, J. Are offshore phytoplankton susceptible to Karenia brevis allelopathy? J. Plankton Res. 2014, 36, 1344–1356. [Google Scholar] [CrossRef] [Green Version]
- Poulson-Ellestad, K.L.; Jones, C.M.; Roy, J.; Viant, M.R.; Fernández, F.M.; Kubanek, J.; Nunn, B.L. Metabolomics and proteomics reveal impacts of chemically mediated competition on marine plankton. Proc. Natl. Acad. Sci. USA 2014, 111, 9009–9014. [Google Scholar] [CrossRef] [Green Version]
- Hakanen, P.; Suikkanen, S.; Kremp, A. Allelopathic activity of the toxic dinoflagellate Alexandrium ostenfeldii: Intra-population variability and response of co-occurring dinoflagellates. Harmful Algae 2014, 39, 287–294. [Google Scholar] [CrossRef]
- Seuront, L.; Stanley, H.E. Anomalous diffusion and multifractality enhance mating encounters in the ocean. Proc. Natl. Acad. Sci. USA 2014, 111, 2206–2211. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, F.; Fong, J.; Sadovskaya, I.; Grard, T.; Vinogradov, E. Structural characterization of the extracellular polysaccharide from Vibrio cholerae O1 El-Tor. PLoS ONE 2014, 9, e86751. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Kjelleberg, S.; McDougald, D. Relative contributions of vibrio polysaccharide and quorum sensing to the resistance of Vibrio cholerae to predation by heterotrophic protists. PLoS ONE 2013, 8, e56338. [Google Scholar] [CrossRef] [Green Version]
- Dadon-Pilosof, A.; Conley, K.R.; Jacobi, Y.; Haber, M.; Lombard, F.; Sutherland, K.R.; Steindler, L.; Tikochinski, Y.; Richter, M.; Glöckner, F.O.; et al. Surface properties of SAR11 bacteria facilitate grazing avoidance. Nat. Microbiol. 2017, 2, 1608–1615. [Google Scholar] [CrossRef]
- Rosa, M.; Ward, J.E.; Holohan, B.A.; Shumway, S.E.; Wikfors, G.H. Physicochemical surface properties of microalgae and their combined effects on particle selection by suspension-feeding bivalve molluscs. J. Exp. Mar. Biol. Ecol. 2017, 486, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Garcés, E.; Alacid, E.; Reñé, A.; Petrou, K.; Simó, R. Host-released dimethylsulphide activates the dinoflagellate parasitoid Parvilucifera sinerae. ISME J. 2013, 7, 1065–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smetacek, V. A watery arms race. Nature 2001, 411, 745. [Google Scholar] [CrossRef]
- Savoca, M.S.; Nevitt, G.A. Evidence that dimethyl sulfide facilitates a tritrophic mutualism between marine primary producers and top predators. Proc. Natl. Acad. Sci. USA 2014, 111, 4157–4161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amo, L.; Rodríguez-Gironés, M.; Barbosa, A. Olfactory detection of dimethyl sulphide in a krill-eating Antarctic penguin. Mar. Ecol. Prog. Ser. 2013, 474, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Kerfahi, D.; Harvey, B.P.; Kim, H.; Yang, Y.; Adams, J.M.; Hall-Spencer, J.M. Whole community and functional gene changes of biofilms on marine plastic debris in response to ocean acidification. Microb. Ecol. 2022, 1, 1–13. [Google Scholar] [CrossRef]
- Imai, I.; Inaba, N.; Yamamoto, K. Harmful algal blooms and environmentally friendly control strategies in Japan. Fish. Sci. 2021, 87, 437–464. [Google Scholar] [CrossRef]
- Karn, S.K.; Bhambri, A.; Jenkinson, I.R.; Duan, J.; Kumar, A. The roles of biomolecules in corrosion induction and inhibition of corrosion: A possible insight. Corros. Rev. 2020, 38, 403–421. [Google Scholar] [CrossRef]
- Qin, B.; Deng, J.; Shi, K.; Wang, J.; Brookes, J.; Zhou, J.; Zhang, Y.; Zhu, G.; Paerl, H.W.; Wu, L. extreme climate anomalies enhancing cyanobacterial blooms in eutrophic Lake Taihu, China. Water Resour. Res. 2021, 57, e2020WR029371. [Google Scholar] [CrossRef]
- McManus, M.; Greer, A.; Timmerman, A.; Sevadjian, J.; Woodson, C.; Cowen, R.; Fong, D.; Monismith, S.; Cheriton, O. Characterization of the biological, physical, and chemical properties of a toxic thin layer in a temperate marine system. Mar. Ecol. Prog. Ser. 2021, 678, 17–35. [Google Scholar] [CrossRef]
- Santschi, P.H.; Xu, C.; Schwehr, K.A.; Lin, P.; Sun, L.; Chin, W.-C.; Kamalanathan, M.; Bacosa, H.P.; Quigg, A. Can the protein/carbohydrate (P/C) ratio of exopolymeric substances (EPS) be used as a proxy for their ‘stickiness’ and aggregation propensity? Mar. Chem. 2020, 218, 103734. [Google Scholar] [CrossRef]
- Duan, Z.; Tan, X.; Zeng, Q. Key physiological traits and chemical properties of extracellular polymeric substances determining colony formation in a cyanobacterium. J. Oceanol. Limnol. 2022, 40, 1720–1731. [Google Scholar] [CrossRef]
- Guadayol, Ò.; Mendonca, T.; Segura-Noguera, M.; Wright, A.J.; Tassieri, M.; Humphries, S. Microrheology reveals microscale viscosity gradients in planktonic systems. Proc. Natl. Acad. Sci. USA 2020, 118, e2011389118. [Google Scholar] [CrossRef]
- Aron, A.T.; Gentry, E.C.; McPhail, K.L.; Nothias, L.-F.; Nothias-Esposito, M.; Bouslimani, A.; Petras, D.; Gauglitz, J.M.; Sikora, N.; Vargas, F.; et al. Reproducible molecular networking of untargeted mass spectrometry data using GNPS. Nat. Protoc. 2020, 15, 1954–1991. [Google Scholar] [CrossRef] [PubMed]
- Lombard, F.; Koski, M.; Kiørboe, T. Copepods use chemical trails to find sinking marine snow aggregates. Limnol. Oceanogr. 2012, 58, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2009, 19, 493–511. [Google Scholar] [CrossRef]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Induction of domoic acid production in the toxic diatom Pseudo-nitzschia seriata by calanoid copepods. Aquat. Toxicol. 2015, 159, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Harðardóttir, S.; Pančić, M.; Tammilehto, A.; Krock, B.; Møller, E.F.; Nielsen, T.G.; Lundholm, N. Dangerous relations in the arctic marine food web: Interactions between toxin producing Pseudo-nitzschia diatoms and Calanus copepodites. Mar. Drugs 2015, 13, 3809–3835. [Google Scholar] [CrossRef] [Green Version]
- Prince, E.K.; Irmer, F.; Pohnert, G. Domoic Acid Improves the Competitive Ability of Pseudo-nitzschia delicatissima against the Diatom Skeletonema marinoi. Mar. Drugs 2013, 11, 2398–2412. [Google Scholar] [CrossRef] [Green Version]
- Hicks, G.R.F.; Grahame, J. Mucus production and its role in the feeding behaviour of Diarthrodes nobilis (Copepoda: Harpacticoida). J. Mar. Biol. Assoc. UK 1979, 59, 321–330. [Google Scholar] [CrossRef]
- Riemann, F.; Schrage, M. The mucus-trap hypothesis on feeding of aquatic nematodes and implications for biodegradation and sediment texture. Oecologia 1978, 34, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.T.; Krock, B.; Hansen, P.J. Production and excretion of okadaic acid, pectenotoxin-2 and a novel dinophysistoxin from the DSP-causing marine dinoflagellate Dinophysis acuta—Effects of light, food availability and growth phase. Harmful Algae 2013, 23, 34–45. [Google Scholar] [CrossRef]
- Ojamäe, K.; Hansen, P.J.; Lips, I. Mass entrapment and lysis of Mesodinium rubrum cells in mucus threads observed in cultures with Dinophysis. Harmful Algae 2016, 55, 77–84. [Google Scholar] [CrossRef]
- Jiang, H.; Kulis, D.M.; Brosnahan, M.L.; Anderson, D.M. Behavioral and mechanistic characteristics of the predator-prey interaction between the dinoflagellate Dinophysis acuminata and the ciliate Mesodinium rubrum. Harmful Algae 2018, 77, 43–54. [Google Scholar] [CrossRef]
- Honsell, G.; Bonifacio, A.; De Bortoli, M.; Penna, A.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Sosa, S.; Yasumoto, T.; et al. New Insights on Cytological and Metabolic Features of Ostreopsis cf. ovata Fukuyo (Dinophyceae): A Multidisciplinary Approach. PLoS ONE 2013, 8, e57291. [Google Scholar] [CrossRef]
- Blossom, H.E.; Daugbjerg, N.; Hansen, P.J. Toxic mucus traps: A novel mechanism that mediates prey uptake in the mixotrophic dinoflagellate Alexandrium pseudogonyaulax. Harmful Algae 2012, 17, 40–53. [Google Scholar] [CrossRef]
- Blossom, H.E.; Hansen, P.J. The loss of mixotrophy in Alexandrium pseudogonyaulax: Implications for trade-offs between toxicity, mucus trap production, and phagotrophy. Limnol. Oceanogr. 2020, 66, 528–542. [Google Scholar] [CrossRef]
- Stehnach, M.R.; Waisbord, N.; Walkama, D.M.; Guasto, J.S. Viscophobic turning dictates microalgae transport in viscosity gradients. Nat. Phys. 2021, 17, 926–930. [Google Scholar] [CrossRef]
- Falciatore, A.; Ribera d’Alcalà, M.; Croot, P.; Bowler, C. Perception of environmental signals by a marine diatom. Science 2000, 288, 2363–2366. [Google Scholar] [CrossRef]
- Wild, C.; Huettel, M.; Klueter, A.; Kremb, S.G.; Rasheed, M.Y.M.; Jørgensen, B.B. Coral mucus functions as an energy carrier and particle trap in the reef ecosystem. Nature 2004, 428, 66–70. [Google Scholar] [CrossRef]
- de Goeij, J.M.; van Oevelen, D.; Vermeij, M.J.A.; Osinga, R.; Middelburg, J.J.; de Goeij, A.F.P.M.; Admiraal, W. Surviving in a Marine Desert: The Sponge Loop Retains Resources Within Coral Reefs. Science 2013, 342, 108–110. [Google Scholar] [CrossRef]
- Samuni, Y.; Goldstein, S.; Dean, O.M.; Berk, M. The chemistry and biological activities of N-acetylcysteine. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2013, 1830, 4117–4129. [Google Scholar] [CrossRef]
- Calzetta, L.; Matera, M.G.; Rogliani, P.; Cazzola, M. Multifaceted activity of N-acetyl-l-cysteine in chronic obstructive pulmonary disease. Expert Rev. Respir. Med. 2018, 12, 693–708. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Z.; Albright, L.J. Anti-phytoplankton therapy of finfish: The mucolytic agent L-cysteine ethyl ester. Dis. Aquat. Org. 1994, 20, 197–202. [Google Scholar] [CrossRef]
- Powell, M.D.; Ransome, J.; Barney, M.; Duijf, R.M.M.; Flik, G. Effect of Dietary Inclusion of N-Acetyl Cysteine on Mucus Viscosity and Susceptibility of Rainbow Trout, Oncorhynchus mykiss, and Atlantic Salmon, Salmo salar, to Amoebic Gill Disease. J. World Aquac. Soc. 2007, 38, 435–442. [Google Scholar] [CrossRef]
- Gutiérrez-Praena, D.; Risalde, M.; Pichardo, S.; Jos, A.; Moyano, R.; Blanco, A.; Vasconcelos, V.; Cameán, A. Histopathological and immunohistochemical analysis of Tilapia (Oreochromis niloticus) exposed to cylindrospermopsin and the effectiveness of N-Acetylcysteine to prevent its toxic effects. Toxicon 2014, 78, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, I.R.; Connors, P.P. The occurrence of the red-tide organism, Gyrodinium aureolum Hulburt (Dinophyceae) around the south and west of Ireland in August and September, 1979. J. Sherkin Isl. 1980, 1, 127–146. [Google Scholar]
- Jenkinson, I.R. Oceanographic implications of non-newtonian properties found in phytoplankton cultures. Nature 1986, 323, 435–437. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Arzul, G. Mitigation by cysteine compounds of rheotoxicity, cytotoxicity and fish mortality caused by the dinoflagellates, Gymnodinium mikimotoi and G. cf. maguelonnense. In Harmful Algal Blooms 2000; Hallegraeff, G., Blackburn, S., Bolch, C., Lewis, R., Eds.; IOC of UNESCO: Paris, France, 2002; pp. 461–464. [Google Scholar]
- Jenkinson, I.R. Increases in viscosity may kill fish in some blooms. In Red Tides; Okaichi, T., Anderson, D.M., Nemoto, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 435–438. [Google Scholar]
- Jenkinson, I.R.; Arzul, G. Effect of the flagellates, Gymnodinium mikimotoi, Heterosigma akashiwo, and Pavlova lutheri, on flow through fish gills. In Harmful Algae; Reguera, B., Blanco, J., Fernandez, M., Wyatt, T., Eds.; Xunta de Galicia: Pontevedra, Spain; IOC of UNESCO: Paris, France, 1998; pp. 425–428. [Google Scholar]
- Gentien, P.; Lunven, M.; Lazure, P.; Youenou, A.; Crassous, M.P. Motility and autotoxicity in Karenia mikimotoi (Dinophyceae). Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1937–1946. [Google Scholar] [CrossRef] [Green Version]
- Orlova, T.Y.; Aleksanin, A.I.; Lepskaya, E.V.; Efimova, K.V.; Selina, M.S.; Morozova, T.V.; Stonik, I.V.; Kachur, V.A.; Karpenko, A.A.; Vinnikov, K.A.; et al. A massive bloom of Karenia species (Dinophyceae) off the Kamchatka coast, Russia, in the fall of 2020. Harmful Algae 2022, 120, 102337. [Google Scholar] [CrossRef]
- Kim, D.; Oda, T. Possible Factors Responsible for the Fish-Killing Mechanisms of the red tide phytoplankton, Chattonella marina and Cochlodinium polykrikoides. In Coastal Environmental and Ecosystem Issues of the East China Sea; Ishimatsu, A., Lie, H.-J., Eds.; Terrapub and Nagasaki University: Nagasaki, Japan, 2010; pp. 245–268. [Google Scholar]
- Berdalet, E.; Tester, P.A.; Chinain, M.; Fraga, S.; Lemée, R.; Litaker, W.; Penna, A.; Usup, G.; Vila, M.; Zingone, A. Harmful Algal Blooms in Benthic Systems: Recent Progress and Future Research. Oceanography 2017, 30, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Seuront, L.; Vincent, D. Increased seawater viscosity, Phaeocystis globosa spring bloom and Temora longicornis feeding and swimming behaviours. Mar. Ecol. Prog. Ser. 2008, 363, 131–145. [Google Scholar] [CrossRef]
- Kang, Z.; Yang, B.; Lai, J.; Ning, Y.; Zhong, Q.; Lu, D.; Liao, R.; Wang, P.; Dan, S.F.; She, Z.; et al. Phaeocystis globosa Bloom Monitoring: Based on P. globosa Induced Seawater Viscosity Modification Adjacent to a Nuclear Power Plant in Qinzhou Bay, China. J. Ocean Univ. China 2020, 19, 1207–1220. [Google Scholar] [CrossRef]
- Balkis-Ozdelice, N.; Durmuş, T.; Balci, M. A Preliminary Study on the Intense Pelagic and Benthic Mucilage Phenomenon Observed in the Sea of Marmara. Int. J. Environ. Geoinform. 2021, 8, 414–422. [Google Scholar] [CrossRef]
- Seuront, L.; Leterme, S.C.; Seymour, J.R.; Mitchell, J.G.; Ashcroft, D.; Noble, W.; Thomson, P.G.; Davidson, A.T.; Enden, R.V.D.; Scott, F.J.; et al. Role of microbial and phytoplanktonic communities in the control of seawater viscosity off East Antarctica (30–80° E). Deep. Sea Res. 2010, 57, 877–886. [Google Scholar] [CrossRef]
- Guzmán, H.M.; Cortés, J.; Glynn, P.W.; Richmond, R.H. Coral mortality associated with dinoflagellate blooms in the eastern Pacific. Mar. Ecol. Prog. Ser. 1990, 60, 299–303. [Google Scholar] [CrossRef]
- Kim, D.; Oda, T.; Muramatsu, T.; Kim, D.; Matsuyama, Y.; Honjo, T. Possible factors responsible for the toxicity of Cochlodinium polykrikoides, a red tide phytoplankton. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 132, 415–423. [Google Scholar] [CrossRef]
- Al Gheilani, H.; Al Azri, A.; Piontkovoski, S.; Debrotsov, S.; Al Amri, I.; Al Ambo Ali, I.; Al Jufaili, S.; Al Bousaidi, S.; Al Hajri, S.; Al Aisari, A.; et al. Blooms of Cochlodinium polykrikoides along the coast of Oman and their effect. In Proceedings of the 14th International Conference Harmful Algae, Hersonissos, Crete, Greece, 1–5 November 2010; Pagou, K., Hallegraeff, G., Eds.; International Society for the Study of Harmful Algae: Copenhagen, Denmark; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2013; pp. 129–131. [Google Scholar]
- Lee, J.-S. Bioactive components from red tide plankton, Cochlodinium polykrikoides. Korean J. Fish. Aquat. Sci. 1996, 29, 165–173. [Google Scholar]
- Kim, C.S.; Lee, S.G.; Lee, C.K.; Kim, H.G.; Jung, J. Reactive oxygen species as causative agents in the ichthyotoxicity of the red tide dinoflagellate Cochlodinium polykrikoides. J. Plankton Res. 1999, 21, 2105–2115. [Google Scholar] [CrossRef]
- Flores-Leñero, A.; Vargas-Torres, V.; Paredes-Mella, J.; Norambuena, L.; Fuenzalida, G.; Lee-Chang, K.; Mardones, J.I. Heterosigma akashiwo in Patagonian Fjords: Genetics, Growth, Pigment Signature and Role of PUFA and ROS in Ichthyotoxicity. Toxins 2022, 14, 577. [Google Scholar] [CrossRef]
- Yamasaki, Y.; Nagasoe, S.; Tameishi, M.; Shikata, T.; Zou, Y.; Jiang, Z.; Matsubara, T.; Shimasaki, Y.; Yamaguchi, K.; Oshima, Y.; et al. The role of interactions between Prorocentrum minimum and Heterosigma akashiwo in bloom formation. Hydrobiologia 2010, 641, 33–44. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, L.; Sims, I.; Beuzenberg, V.; Gillespie, P. Mass accumulation of mucilage caused by dinoflagellate polysaccharide exudates in Tasman Bay, New Zealand. Harmful Algae 2002, 1, 69–83. [Google Scholar] [CrossRef]
- Honsell, G.; Cabrini, M.; Darin, M. Gonyaulax fragilis (Schütt) Kofoid: A dinoflagellate from gelatinous aggregates of the Northern Adriatic Sea. G. Bot. Ital. 1992, 126, 749–751. [Google Scholar] [CrossRef]
- Pompei, M.; Mazziotti, C.; Guerrini, F.; Cangini, M.; Pigozzi, S.; Benzi, M.; Palamidesi, S.; Boni, L.; Pistocchi, R. Correlation between the presence of Gonyaulax fragilis (Dinophyceae) and the mucilage phenomena of the Emilia-Romagna coast (northern Adriatic Sea). Harmful Algae 2003, 2, 301–316. [Google Scholar] [CrossRef]
- Carbonell-Moore, M.C.; Mertens, K.N. Should Gonyaulax hyalina and Gonyaulax fragilis (Dinophyceae) remain two different taxa? Phycologia 2019, 58, 685–689. [Google Scholar] [CrossRef]
- Riccardi, M.; Guerrini, F.; Serrazanetti, G.P.; Ventrella, V.; Pagliarani, A.; Pistocchi, R. Lipid and DNA features of Gonyaulax fragilis (Dinophyceae) as potential biomarkers in mucilage genesis. Harmful Algae 2010, 9, 359–366. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, X. Single molecule mechanochemistry of macromolecules. Prog. Polym. Sci. 2003, 28, 1271–1295. [Google Scholar] [CrossRef]
- Wiita, A.P.; Ainavarapu, S.R.K.; Huang, H.H.; Fernandez, J.M. Force-dependent chemical kinetics of disulfide bond reduction observed with single-molecule techniques. Proc. Natl. Acad. Sci. USA 2006, 103, 7222–7227. [Google Scholar] [CrossRef] [Green Version]
- Alting, A.C.; Hamer, R.J.; de Kruif, C.G.; Paques, M.; Visschers, R.W. Number of thiol groups rather than the size of the aggregates determines the hardness of cold set whey protein gels. Food Hydrocoll. 2003, 17, 469–479. [Google Scholar] [CrossRef]
- Quigg, A.; Santschi, P.; Burd, A.; Chin, W.-C.; Kamalanathan, M.; Xu, C.; Ziervogel, K. From Nano-Gels to Marine Snow: A Synthesis of Gel Formation Processes and Modeling Efforts Involved with Particle Flux in the Ocean. Gels 2021, 7, 114. [Google Scholar] [CrossRef]
- Hu, X.-J.; Xu, Y.; Su, H.-C.; Xu, W.-J.; Wang, L.-H.; Xu, Y.-N.; Li, Z.-J.; Cao, Y.-C.; Wen, G.-L. Algicidal bacterium CZBC1 inhibits the growth of Oscillatoria chlorina, Oscillatoria tenuis, and Oscillatoria planctonica. AMB Express. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-O.; Kato, J.; Takiguchi, N.; Kuroda, A.; Ikeda, T.; Mitsutani, A.; Ohtake, H. Involvement of an Extracellular Protease in Algicidal Activity of the Marine Bacterium Pseudoalteromonas sp. Strain A28. Appl. Environ. Microbiol. 2000, 66, 4334–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Yang, Y.; Zhu, X.-Y.; Zhang, X.-H.; Yu, M. Metagenomic Insights Into the Microbial Assemblage Capable of Quorum Sensing and Quorum Quenching in Particulate Organic Matter in the Yellow Sea. Front. Microbiol. 2021, 11, 602010. [Google Scholar] [CrossRef] [PubMed]
- Svedrup, H.U.; Johnson, M.W.; Fleming, R.H. The Oceans: Their Physics, Chemistry, and General Biology, 1st ed.; Prentice Hall: New York, NY, USA, 1942. [Google Scholar]
- Endo, M. Calcium Ion as a Second Messenger With Special Reference to Excitation-Contraction Coupling. J. Pharmacol. Sci. 2006, 100, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Ianora, A.; Boersma, M.; Casotti, R.; Fontana, A.; Harder, J.; Hoffmann, F.; Pavia, H.; Potin, P.; Poulet, S.A.; Toth, G. New trends in marine chemical ecology. Estuaries Coasts 2006, 29, 531–551. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, I.R.; Wyatt, T. Selection and control of Deborah numbers in plankton ecology. J. Plankton Res. 1992, 14, 1697–1721. [Google Scholar] [CrossRef]
- Seuront, L.; Lacheze, C.; Doubell, M.J.; Seymour, J.R.; Van Dongen-Vogels, V.; Newton, K.; Alderkamp, A.C.; Mitchell, J.G. The influence of Phaeocystis globosa on microscale spatial patterns of chlorophyll a and bulk-phase seawater viscosity. Biodegradation 2007, 83, 173–188. [Google Scholar] [CrossRef]
- Wyatt, T.; Ribera d’Alcalà, M. Dissolved organic matter and planktonic engineerin. CIESM Workshop Monogrs. 2006, 28, 13–23. [Google Scholar]
- Hastings, A.; Byers, J.; Crooks, J.A.; Cuddington, K.; Jones, C.G.; Lambrinos, J.G.; Talley, T.S.; Wilson, W.G. Ecosystem engineering in space and time. Ecol. Lett. 2007, 10, 153–164. [Google Scholar] [CrossRef]
- Reddington, K.; Eccles, D.; O’Grady, J.; Drown, D.M.; Hansen, L.H.; Nielsen, T.K.; Ducluzeau, A.-L.; Leggett, R.M.; Heavens, D.; Peel, N.; et al. Metagenomic analysis of planktonic riverine microbial consortia using nanopore sequencing reveals insight into river microbe taxonomy and function. Gigascience 2020, 9, giaa053. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Wyatt, T. Management by phytoplankton of physical oceanographic parameters. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier Intercept: Paris, France, 1995; pp. 603–607. [Google Scholar]
- Camacho-Chab, J.C.; Lango-Reynoso, F.; Castañeda-Chávez, M.D.R.; Galaviz-Villa, I.; Hinojosa-Garro, D.; Ortega-Morales, B.O. Implications of Extracellular Polymeric Substance Matrices of Microbial Habitats Associated with Coastal Aquaculture Systems. Water 2016, 8, 369. [Google Scholar] [CrossRef]
- Sretenovic, S.; Stojković, B.; Dogsa, I.; Kostanjšek, R.; Poberaj, I.; Stopar, D. An early mechanical coupling of planktonic bacteria in dilute suspensions. Nat. Commun. 2017, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Jatt, A.N.; Tang, K.; Liu, J.; Zhang, Z.; Zhang, X.-H. Quorum sensing in marine snow and its possible influence on production of extracellular hydrolytic enzymes in marine snow bacterium Pantoea ananatis B9. FEMS Microbiol. Ecol. 2014, 91, 1–13. [Google Scholar] [CrossRef]
- Decho, A.W.; Gutierrez, T. Microbial Extracellular Polymeric Substances (EPSs) in Ocean Systems. Front. Microbiol. 2017, 8, 922. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Tay, Q.X.M.; Kjelleberg, S.; Rice, S.A.; McDougald, D. Quorum sensing-regulated chitin metabolism provides grazing resistance to Vibrio cholerae biofilms. ISME J. 2015, 9, 1812–1820. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.; Svenningsen, S.L.; Middelboe, M. Quorum Sensing Determines the Choice of Antiphage Defense Strategy in Vibrio anguillarum. Mbio 2015, 6, e00627-15. [Google Scholar] [CrossRef] [Green Version]
- Pappas, K.M.; Weingart, C.L.; Winans, S.C. Chemical communication in proteobacteria: Biochemical and structural studies of signal synthases and receptors required for intercellular signalling. Mol. Microbiol. 2004, 53, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Grossart, H.-P.; Schlingloff, A.; Kiørboe, T. Possible Quorum Sensing in Marine Snow Bacteria: Production of Acylated Homoserine Lactones by Roseobacter Strains Isolated from Marine Snow. Appl. Environ. Microbiol. 2002, 68, 4111–4116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsek, M.R.; Fuqua, C.; Bordi, C.; Ansaldi, M.; Gon, S.; Jourlin-Castelli, C.; Iobbi-Nivol, C.; Méjean, V. Biofilms 2003: Emerging Themes and Challenges in Studies of Surface-Associated Microbial Life. J. Bacteriol. 2004, 186, 4502–4509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tait, K.; Joint, I.; Daykin, M.; Milton, D.L.; Williams, P.; Camara, M. Disruption of quorum sensing in seawater abolishes attraction of zoospores of the green alga Ulva to bacterial biofilms. Environ. Microbiol. 2004, 7, 229–240. [Google Scholar] [CrossRef]
- Wheeler, G.L.; Tait, K.; Taylor, A.; Brownlee, C.; Joint, I. Acyl-homoserine lactones modulate the settlement rate of zoospores of the marine alga Ulva intestinalis via a novel chemokinetic mechanism. Plant Cell Environ. 2006, 29, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, J.; Lan, C.; Lai, J.; Wang, J.; Wei, F. Inhibitory effect of increase in water viscosity of Phaeocystis globosa on Feeding of Temora stylifera. Guangxi Sci. 2021, 28, 56–64, (In Chinese, English abstract). [Google Scholar]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Hohne, D.N.; Younger, J.G.; Solomon, M.J. Flexible Microfluidic Device for Mechanical Property Characterization of Soft Viscoelastic Solids Such as Bacterial Biofilms. Langmuir 2009, 25, 7743–7751. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Chin, W.-C.; Chiu, M.-H.; Xu, C.; Lin, P.; Schwehr, K.A.; Quigg, A.; Santschi, P.H. Sunlight induced aggregation of dissolved organic matter: Role of proteins in linking organic carbon and nitrogen cycling in seawater. Sci. Total. Environ. 2018, 654, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, N.; Dai, X.; Tao, W.; Jenkinson, I.R.; Li, Z. Enhanced dewaterability of sludge during anaerobic digestion with thermal hydrolysis pretreatment: New insights through structure evolution. Water Res. 2018, 131, 177–185. [Google Scholar] [CrossRef]
- Lear, G. (Ed.) Biofilms in Bioremediation; Caister Academic Press: Poole, UK, 2016; Available online: https://www.lulu.com/spotlight/caister (accessed on 15 February 2023).
- Verdugo, P. Marine Microgels. Annu. Rev. Mar. Sci. 2012, 4, 375–400. [Google Scholar] [CrossRef]
- Verdugo, P. Marine Biopolymer Dynamics, Gel Formation, and Carbon Cycling in the Ocean. Gels 2021, 7, 136. [Google Scholar] [CrossRef]
- Drost-Hansen, W. Vicinal hydration of biopolymers: Cell biological consequences. In Water and the Cell; Pollack, G.H., Cameron, I.L., Wheatley, D.N., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 175–217. [Google Scholar]
- Mari, X. Does ocean acidification induce an upward flux of marine aggregates? Biogeosciences 2008, 5, 1023–1031. [Google Scholar] [CrossRef] [Green Version]
- Bartual, A.; Cera, I.V.; Flecha, S.; Prieto, L. Effect of dissolved polyunsaturated aldehydes on the size distribution of transparent exopolymeric particles in an experimental diatom bloom. Mar. Biol. 2017, 164, 120. [Google Scholar] [CrossRef]
- Klun, K.; Šket, P.; Beran, A.; Falnoga, I.; Faganeli, J. Composition of Colloidal Organic Matter in Phytoplankton Exudates. Water 2022, 15, 111. [Google Scholar] [CrossRef]
- Vidal-Melgosa, S.; Sichert, A.; Ben Francis, T.; Bartosik, D.; Niggemann, J.; Wichels, A.; Willats, W.G.T.; Fuchs, B.M.; Teeling, H.; Becher, D.; et al. Diatom fucan polysaccharide precipitates carbon during algal blooms. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Denis, M.; Lefevre, D.; Thyssen, M.; Jenkinson, I.R.; Grégori, G. Pulsed export of carbon in the north-western Mediterranean Sea. J. Oceanol. Limnol. 2023, 41, 189–202. [Google Scholar] [CrossRef]
- Liu, L.; Qin, B.; Zhu, G.; Zhang, Y.; Gao, G.; Gong, Z.; Huang, Q. Distribution of dissolved acidic polysaccharides (dAPS) during cyanobacteria blooms in northern Lake Taihu. Limnology 2014, 16, 21–29. [Google Scholar] [CrossRef]
- Laabir, M.; Grignon-Dubois, M.; Masseret, E.; Rezzonico, B.; Soteras, G.; Rouquette, M.; Rieuvilleneuve, F.; Cecchi, P. Algicidal effects of Zostera marina L. and Zostera noltii Hornem. extracts on the neuro-toxic bloom-forming dinoflagellate Alexandrium catenella. Aquat. Bot. 2013, 111, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Onishi, Y.; Mohri, Y.; Tuji, A.; Ohgi, K.; Yamaguchi, A.; Imai, I. The seagrass Zostera marina harbors growth-inhibiting bacteria against the toxic dinoflagellate Alexandrium tamarense. Fish. Sci. 2014, 80, 353–362. [Google Scholar] [CrossRef]
- Imai, I.; Kido, T.; Yoshinaga, I.; Ohgi, K.; Nagai, S. Isolation of Microcystis-killer bacterium Agrobacterium vitis from the biofilm onthe surface of the water plant Egeria densa. In Proceedings of the 14th International Conference Harmful Algae, Hersonissos, Crete, Greece, 1–5 November 2010; Pagou, K.A., Hallegraeff, G.M., Eds.; ISSHA: Copenhagen, Denmark; IOC of UNESCO: Paris, France, 2013; pp. 150–152. [Google Scholar]
- Yang, Y.; Hu, X.; Zhang, J.; Gong, Y. Community level physiological study of algicidal bacteria in the phycospheres of Skeletonema costatum and Scrippsiella trochoidea. Harmful Algae 2013, 28, 88–96. [Google Scholar] [CrossRef]
- Qian, X.; Chen, L.; Sui, Y.; Chen, C.; Zhang, W.; Zhou, J.; Dong, W.; Jiang, M.; Xin, F.; Ochsenreither, K. Biotechnological potential and applications of microbial consortia. Biotechnol. Adv. 2020, 40, 107500. [Google Scholar] [CrossRef]
- Kotay, S.M.; Das, D. Microbial hydrogen production from sewage sludge bioaugmented with a constructed microbial consortium. Int. J. Hydrog. Energy 2010, 35, 10653–10659. [Google Scholar] [CrossRef]
- Thakur, P.; Alaba, M.O.; Rauniyar, S.; Singh, R.N.; Saxena, P.; Bomgni, A.; Gnimpieba, E.Z.; Lushbough, C.; Goh, K.M.; Sani, R.K. Text-Mining to Identify Gene Sets Involved in Biocorrosion by Sulfate-Reducing Bacteria: A Semi-Automated Workflow. Microorganisms 2023, 11, 119. [Google Scholar] [CrossRef]
- Singh, A. Glycoproteomics. Nat. Methods 2021, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Alldredge, A.L.; Passow, U.; Logan, B.E. The abundance and significance of a class of large, transparent organic particles in the ocean. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1993, 40, 1131–1140. [Google Scholar] [CrossRef]
- Williams, P.; Carlucci, A.; Henrichs, S.; Van Vleet, E.; Horrigan, S.; Reid, F.; Robertson, K. Chemical and microbiological studies of sea-surface films in the Southern Gulf of California and off the West Coast of Baja California. Mar. Chem. 1986, 19, 17–98. [Google Scholar] [CrossRef]
- Carlson, D.J. Viscosity of sea-surface slicks. Nature 1987, 329, 823–825. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, L.; Liu, C.; Cai, W. Viscosity of sea surface microlayer in Jiaozhou Bay and adjacent sea area. Chin. J. Oceanol. Limnol. 2003, 21, 351–357. [Google Scholar] [CrossRef]
- Jenkinson, I.R.; Seuront, L.; Ding, H.; Elias, F. Biological modification of mechanical properties of the sea surface microlayer, influencing waves, ripples, foam and air-sea fluxes. Elem. Sci. Anthr. 2018, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Miles, J.; Lovelock, J. The Ages of Gaia. A Biography of Our Living Earth. J. Appl. Ecol. 1989, 26, 736. [Google Scholar] [CrossRef]
- Lenton, T.M. Gaia and natural selection. Nature 1998, 394, 439–447. [Google Scholar] [CrossRef]
- Thornton, D.C. Phytoplankton mucilage production in coastal waters: A dispersal mechanism in a front dominated system? Ethol. Ecol. Evol. 1999, 11, 179–185. [Google Scholar] [CrossRef]
- Hamilton, W.; Lenton, T. Spora and Gaia: How microbes fly with their clouds. Ethol. Ecol. Evol. 1998, 10, 1–16. [Google Scholar] [CrossRef]
- Teeling, H.; Fuchs, B.M.; Becher, D.; Klockow, C.; Gardebrecht, A.; Bennke, C.M.; Kassabgy, M.; Huang, S.; Mann, A.J.; Waldmann, J.; et al. Substrate-Controlled Succession of Marine Bacterioplankton Populations Induced by a Phytoplankton Bloom. Science 2012, 336, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Boyd, P.W.; Claustre, H.; Levy, M.; Siegel, D.A.; Weber, T. Multi-faceted particle pumps drive carbon sequestration in the ocean. Nature 2019, 568, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Friedlingstein, P.; O’Sullivan, M.; Jones, M.W.; Andrew, R.M.; Gregor, L.; Hauck, J.; Le Quéré, C.; Luijkx, I.T.; Olsen, A.; Peters, G.P.; et al. Global Carbon Budget 2022. Earth Syst. Sci. Data 2022, 14, 4811–4900. [Google Scholar] [CrossRef]
- Gulev, S.K.; Thorne, P.W. Chapter 2: Changing state of the climate system. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; pp. 287–422. [Google Scholar]
- Stabeno, P.J.; Monahan, E.C. The Influence of whitecaps on the albedo of the sea surface. In Oceanographic Science Library; Monahan, E.C., Mac Niocaill, G., Eds.; D. Reidel: Dordrecht, Germany; Galway University Press: Galway, Ireland, 1986; pp. 261–266. [Google Scholar]
- Evans, J.R.G.; Stride, E.P.J.; Edirisinghe, M.J.; Andrews, D.J.; Simon, R.R. Can oceanic foams limit global warming? Clim. Res. 2010, 42, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Cantat, I.; Cohen-Addad, S.; Elias, F.; Graner, F.; Höhler, R.; Pitois, O.; Rouyer, F.; Saint-Jalmes, A.; Cox, S. (Eds.) Foams; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Callaghan, A.H.; Deane, G.B.; Stokes, M.D.; Ward, B. Observed variation in the decay time of oceanic whitecap foam. J. Geophys. Res. Atmos. 2012, 117, C09015. [Google Scholar] [CrossRef]
- Pierce, R.H.; Henry, M.S.; Blum, P.C.; Lyons, J.; Cheng, Y.S.; Yazzie, D.; Zhou, Y. Brevetoxin Concentrations in Marine Aerosol: Human Exposure Levels During a Karenia brevis Harmful Algal Bloom. Bull. Environ. Contam. Toxicol. 2003, 70, 161–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Montero, M.; Freer, E.; Jiménez-Montealegre, R.; Guzmán, J. Occurrence and predominance of the fish killer Cochlod-inium polykrikoides on the Pacific coast of Costa Rica. Afr. J. Mar. Sci. 2006, 28, 215–217. [Google Scholar] [CrossRef]
- Kesaulya, I.; Leterme, S.C.; Mitchell, J.G.; Seuront, L. The impact of turbulence and phytoplankton dynamics on foam formation, seawater viscosity and chlorophyll concentration in the eastern English Channel. Oceanologia 2008, 50, 167–182. [Google Scholar]
- Calvo, E.; Viquez, R.; Garcia, A. Alexandrium monilatum (Howell) Balech bloom in the Gulf of Nicoya, Puntarenas. Harmful Algae News 2005, 29, 1–2. [Google Scholar]
- Lassus, P.; Chomérat, N.; Hess, P.; Nézan, E. Toxic and Harmful Microalgae of the World Ocean; ISSHA: Copenhagen, Denmark; IOC of UNESCO: Paris, France, 2016. [Google Scholar]
Figure 1.
Three categories of gene-bearing (“living”) organisms are presented: viruses, prokaryotes and eukaryotes. Prokaryotes and eukaryotes are shown in the same compartment, as generally they both have an independent machinery to form amino acids and proteins (including enzymes) from “instructions” in their genes. Enzymes modulate (catalyze) intracellular molecular transformations (modulation pathways not shown). Viruses are placed in a separate box as they need to introduce their genes into prokaryotic or eukaryotic cells, taking some control of these cells in order to reproduce their genes and outer casing. Red arrows represent the pathways of viral reproduction and transfer of viral genes to replace or modulate prokaryotic or eukaryotic genes [16]. Red arrows are used also to represent transfers between the major types of organic molecules inside cells. Black arrows represent transformations between the major suggested extracellular pools of organic matter (OM). Blue arrows represent modulation by extracellular polymeric substances (EPS) on physical and physicochemical processes in the ocean and at the ocean–atmosphere interface, as well as secondary effects in the ocean and the atmosphere (inside red box). Blue arrows also represent signaling and intoxication pathways by signaling molecules and toxins, respectively. Eukaryotic plankton is represented by A—a dinoflagellate; B—a diatom; C—a jellyfish medusa; D—a raphidophyte. This is not a sketch of trophic or energy pathways.
Figure 1.
Three categories of gene-bearing (“living”) organisms are presented: viruses, prokaryotes and eukaryotes. Prokaryotes and eukaryotes are shown in the same compartment, as generally they both have an independent machinery to form amino acids and proteins (including enzymes) from “instructions” in their genes. Enzymes modulate (catalyze) intracellular molecular transformations (modulation pathways not shown). Viruses are placed in a separate box as they need to introduce their genes into prokaryotic or eukaryotic cells, taking some control of these cells in order to reproduce their genes and outer casing. Red arrows represent the pathways of viral reproduction and transfer of viral genes to replace or modulate prokaryotic or eukaryotic genes [16]. Red arrows are used also to represent transfers between the major types of organic molecules inside cells. Black arrows represent transformations between the major suggested extracellular pools of organic matter (OM). Blue arrows represent modulation by extracellular polymeric substances (EPS) on physical and physicochemical processes in the ocean and at the ocean–atmosphere interface, as well as secondary effects in the ocean and the atmosphere (inside red box). Blue arrows also represent signaling and intoxication pathways by signaling molecules and toxins, respectively. Eukaryotic plankton is represented by A—a dinoflagellate; B—a diatom; C—a jellyfish medusa; D—a raphidophyte. This is not a sketch of trophic or energy pathways.


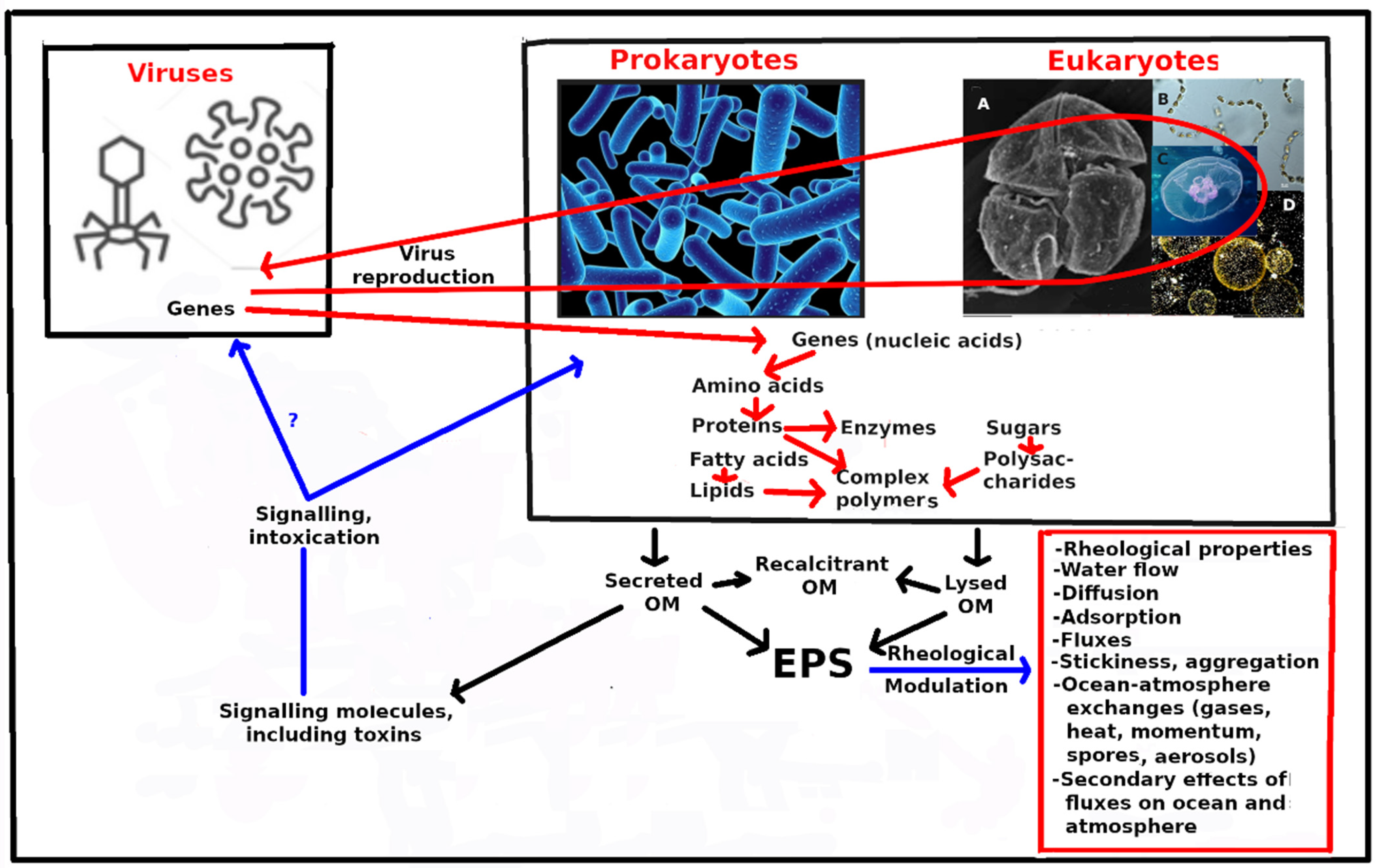{kind=link}
Table 1.
Global coastal ocean nitrogen, N, and iron, Fe, budget terms (Tg y−1) (estimated annual means).
Table 1.
Global coastal ocean nitrogen, N, and iron, Fe, budget terms (Tg y−1) (estimated annual means).
| Coastal Ocean Ref. [17] | Whole Ocean Refs. [18,19] | Open Ocean Ref. [19] | |
|---|---|---|---|
| Inorganic N (IN) | |||
| Atmospheric deposition (DIN) | +4.5 | ||
| River input (DIN) | +20.4 | +23 [19] | +17 (a) |
| Denitrification (water + sediments) (DIN) | −51.9 | ||
| Coast-ward net influx from offshore (DIN) | +47.4 | ||
| Total ΔTIN | +20.4 | ||
| Organic and inorganic N | |||
| Sedimentary burial (TON) | −12.3 | ||
| River input (TON) | +27.1 | ||
| River input (DON) | +11 (a) [19] | >0 to <11 (a) | |
| N2 fixation (TON) | +15.4 | 164 [19] | 0 |
| Oceanward net outflux to offshore (TON) | −50.2 | ||
| Total Δ TON | −20.0 | ||
| Discrepancy (ΔTIN − ΔTON) | +0.4 | ||
| Net community production (DIN + TON) | 35.5 | ||
| Atmospheric N deposition | 39 (b) [19] | >30 (b) | |
| Atmosphere-ocean Fe budget | |||
| Fe emissions from fires (<20 µm) | 1.1 [18] | ||
| Soluble Fe flux to the ocean (from dust) | 0.19 to 0.28 [18] | ||
| Soluble Fe flux to the ocean (from fires) | 0.035 to 0.063 [18] | ||
| Soluble Fe flux to the ocean (anthropogenic) | 0.016 to 0.034 [18] | ||
| Soluble Fe flux to the ocean (Total) | 0.24 to 0.38 [18] | ||
| Atmosphere-ocean P budget Soluble P flux to the ocean (from dust) | 0.031 to 0.094 [18] | ||
| Soluble P flux to the ocean (from fires) | 0.005 [18] | ||
| Soluble P flux to the ocean (anthropogenic) | 0.0094 to 0.11 [18] | ||
| Soluble P flux to the ocean (Total) | 0.045 to 0.21 [18] |
a—75% of riverine input escapes beyond the shelf break; b—75% of atmospheric input deposited outside the shelf break.
Table 2.
Distribution of the integrated main groups of proton resonances (lipids, proteins and carboxyl-rich alicyclic molecules (CRAM), polysaccharides and formate) in 1H MNR spectra (6/ppm), concentrations of Corg. (µmol L−1) in retentates and permeates and % COC from exudates of cultured phytoplankters. Protons that resonated in certain chemical shift range of integrated main groups are in bold (second row). Summarized from Klun et al. [144].
Table 2.
Distribution of the integrated main groups of proton resonances (lipids, proteins and carboxyl-rich alicyclic molecules (CRAM), polysaccharides and formate) in 1H MNR spectra (6/ppm), concentrations of Corg. (µmol L−1) in retentates and permeates and % COC from exudates of cultured phytoplankters. Protons that resonated in certain chemical shift range of integrated main groups are in bold (second row). Summarized from Klun et al. [144].
| Lipids | Proteins and CRAM | Polysaccharides | Formate | Corg. | % COC * | |
|---|---|---|---|---|---|---|
| HCH2-CH2- | HC-HCOR | HC-OH HC-O-C | HCOO | |||
| δ **/ppm | 0–1.8 | 1.8–3.0 | 3.0–4.6 | 8.0–9.0 | µmol L−1 | % |
| Tetraselmis sp. | ||||||
| 0.2 µm filtrate | 915.1 | |||||
| Retentate | 19.4 | 18.8 | 61.4 | 0.4 | 364 | 39.8 |
| Permeate | 35.7 | 40.9 | 20.8 | 2.6 | 507 | |
| Chaetoceros socialis | ||||||
| 0.2 µm filtrate | 2285 | |||||
| Retentate | 14.4 | 21.9 | 62.9 | 1.1 | 526 | 23.0 |
| Permeate | 16.4 | 36.4 | 45.5 | 1.7 | 1765 | |
| Prorocentrum minimum | ||||||
| 0.2 µm filtrate | 439.3 | |||||
| Retentate | 21.8 | 45.6 | 31.9 | 0.8 | 154 | 35.1 |
| Permeate | 17.4 | 56.7 | 24.6 | 1.2 | 418 | |
Note(s): * Colloidal organic carbon percentage: %COC = Corg (retentate) × 100%/Corg. (0.2 µm filtrate). ** δ is chemical shift range in 1H NMR spectra for each group of compounds.
Table 3.
Synthesis of major inputs of organic matter to the open ocean.
| Mechanism | Global Magnitude (PgCyr−1) | Depth Horizon | Remarks |
|---|---|---|---|
| Downward transport from the photic zone into the ocean interior. (Recompiled from Boyd et al. [167] and references therein.) | |||
| Biological gravitational pump | 4.9 to 9.1 | 100 m or euphotic zone. | |
| Mesopelagic migrant pump | 0.9 to 3.6 | ||
| Seasonal lipid pump | 0.25 to 1.0 | ||
| Mixed layer pump | 0.1 to 1.0 | Mixed layer. | |
| Large-scale subduction pump | 0.13 | ||
| Eddy subduction pump | 2.0 | 100 m. | Low confidence of precision |
| Sum of physical pumps | 1.8 to 2.1 | Mixed layer, euphotic zone or 74 m. | Low confidence of precision |
| Aquatic input of anthropogenic OC from the land, and from total primary production in situ. | |||
| Input of land-derived OC to the open ocean through the land-ocean aquatic continuum (LOAC) | 0.65 | Probably mostly input to photic zone, but an unknown fraction is rapidly subducted to below the photic zone. | Low confidence of precision [168] |
| Global marine primary productivity | 47 (±8) | IPCC [169] N.B. This estimate includes coastal waters. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jenkinson, I.R. Plankton Genes and Extracellular Organic Substances in the Ocean. J. Mar. Sci. Eng. 2023, 11, 783. https://doi.org/10.3390/jmse11040783
AMA Style
Jenkinson IR. Plankton Genes and Extracellular Organic Substances in the Ocean. Journal of Marine Science and Engineering. 2023; 11(4):783. https://doi.org/10.3390/jmse11040783
Chicago/Turabian StyleJenkinson, Ian R. 2023. "Plankton Genes and Extracellular Organic Substances in the Ocean" Journal of Marine Science and Engineering 11, no. 4: 783. https://doi.org/10.3390/jmse11040783
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.