
Study of Phytoplankton Biomass and Environmental Drivers in and around the Ross Sea Marine Protected Area

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
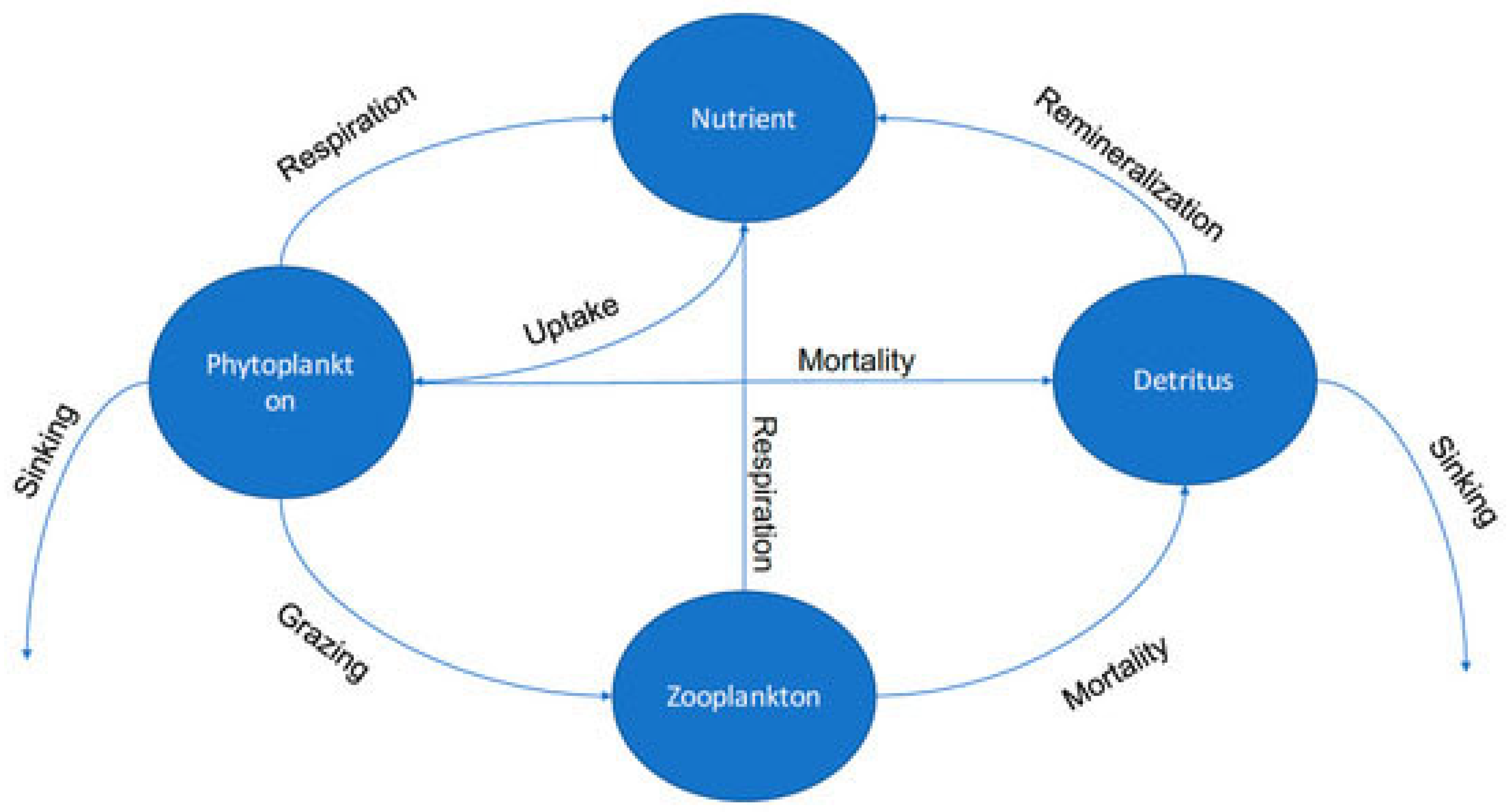
2.2. Model Description
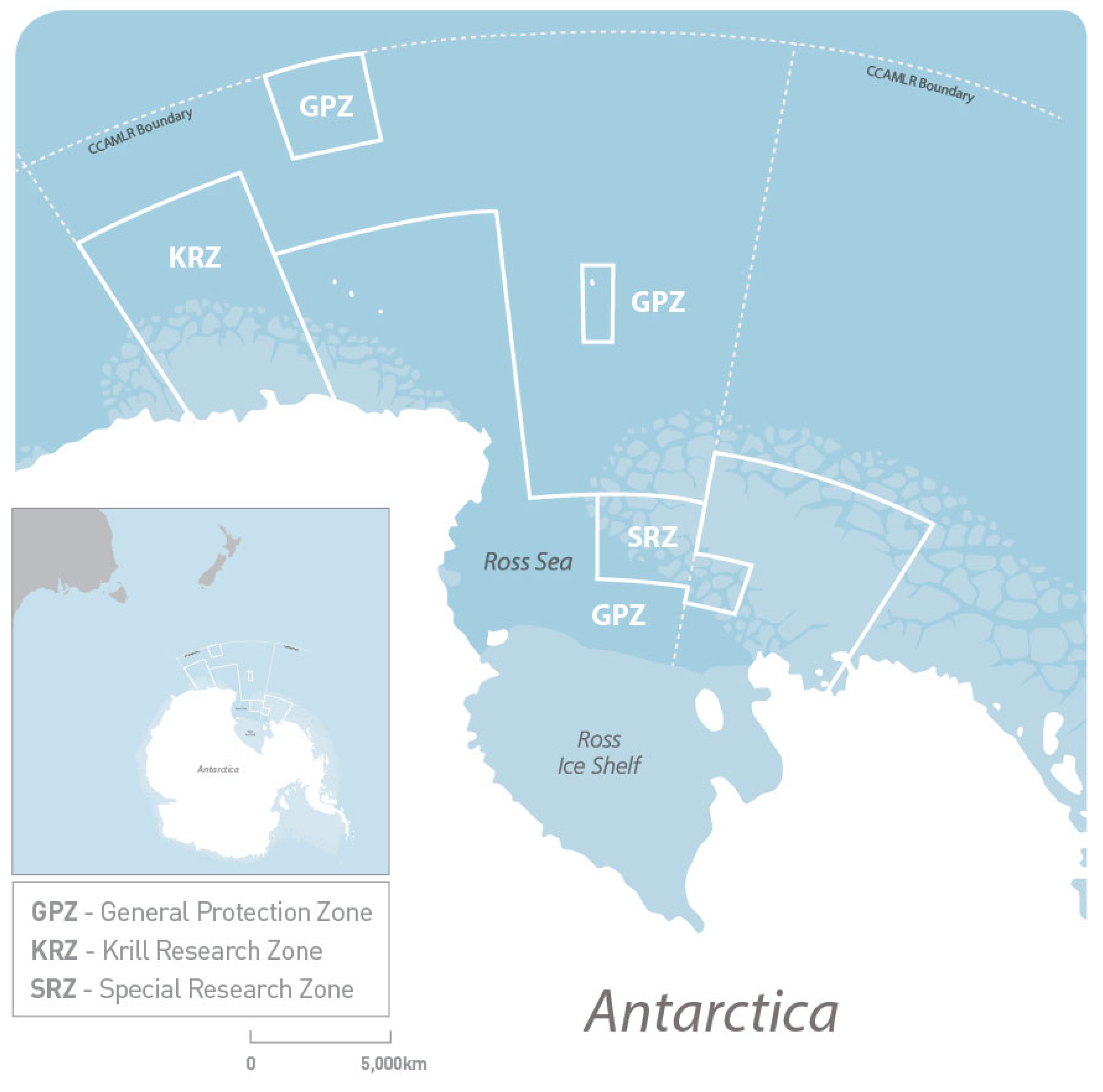
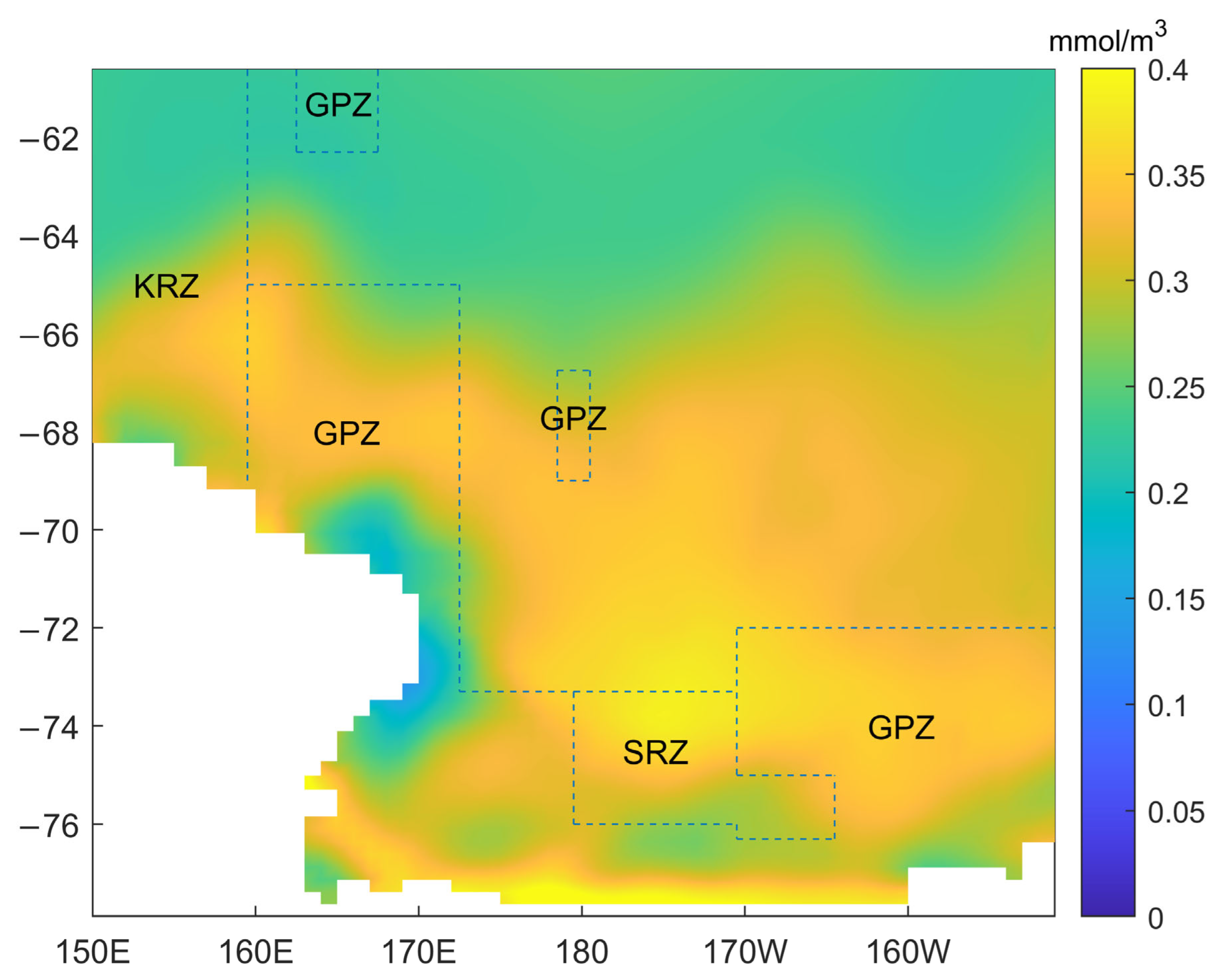
2.3. Study Area Division
- (1)
- The area bounded by a line starting where the meridian at 160° E intersects the coastline, thence due north to 65° S, thence due east to 173°45′ E, thence due south to 73°30′ S, thence due east to 180°, thence due south to 76° S, thence due east to 170° W, thence due south to 76°30′ S, thence due east to 164° W, thence due north to 75° S, thence due west to 170° W, thence due north to 72° S, thence due east to 150° W, thence due south to the coastline, and thence along the coastline to the starting point.
- (2)
- The area bounded by a line starting at 62°30′ S 163° E, thence due north to 60° S, thence due east to 168° E, thence due south to 62°30′ S, and thence due west to the starting point.
- (3)
- The area bounded by a line starting at 69° S 179° E, thence due north to 66°45′ S, thence due east to 179° W, thence due south to 69° S, and thence due west to the starting point.
2.4. Method
3. Results
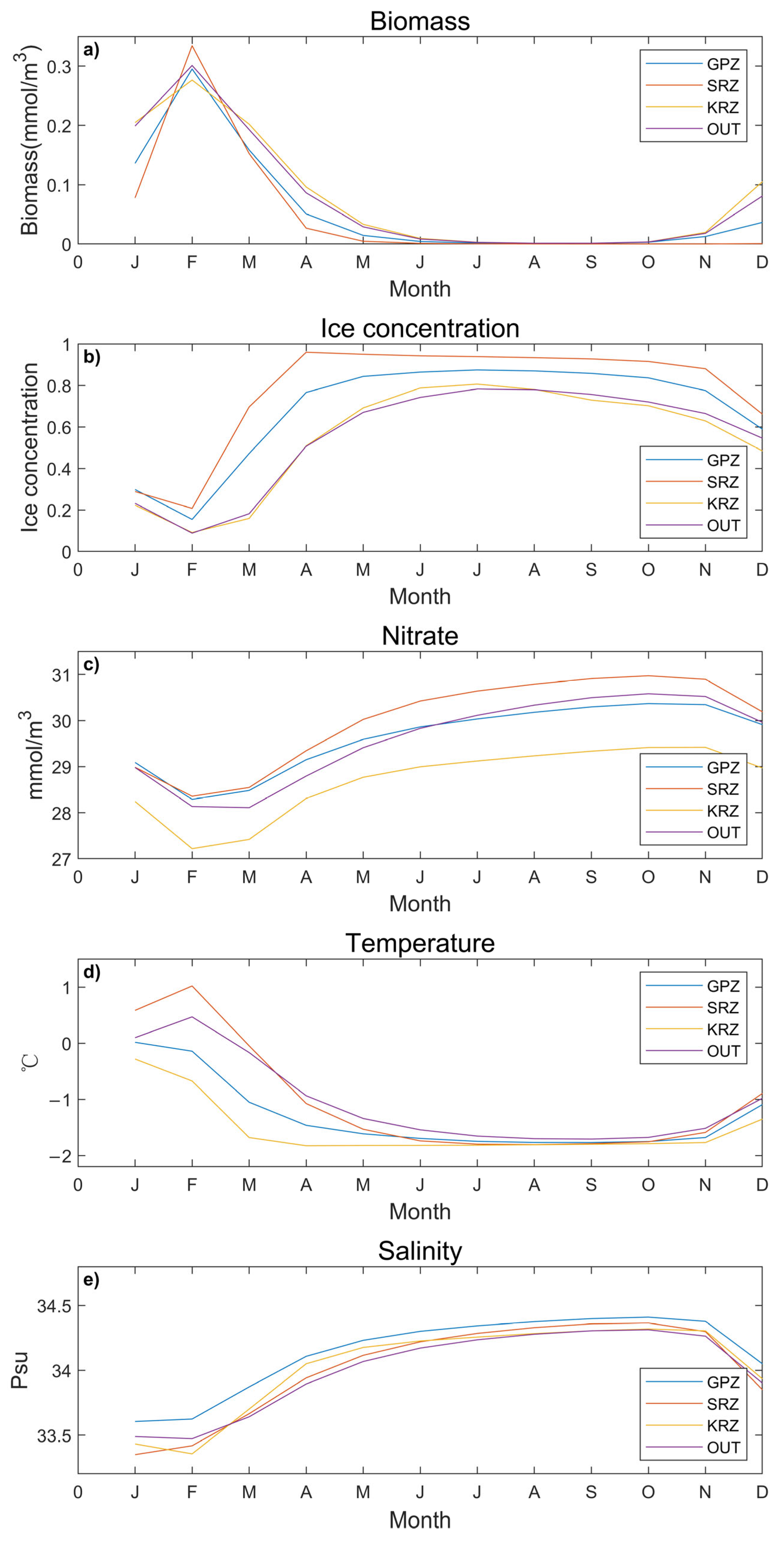
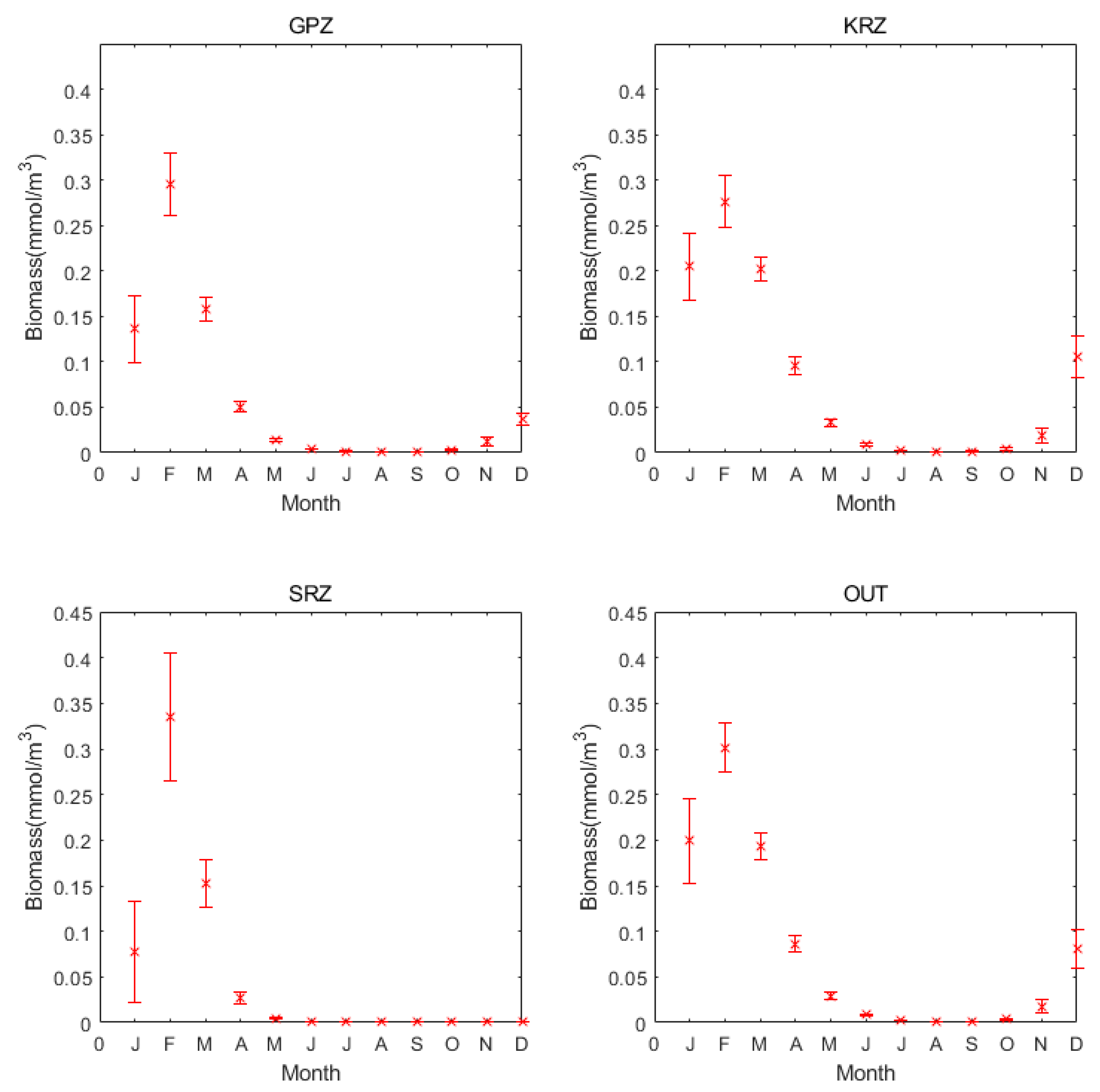
3.1. Annual Cycle Trends of Different MPA in the Ross Sea
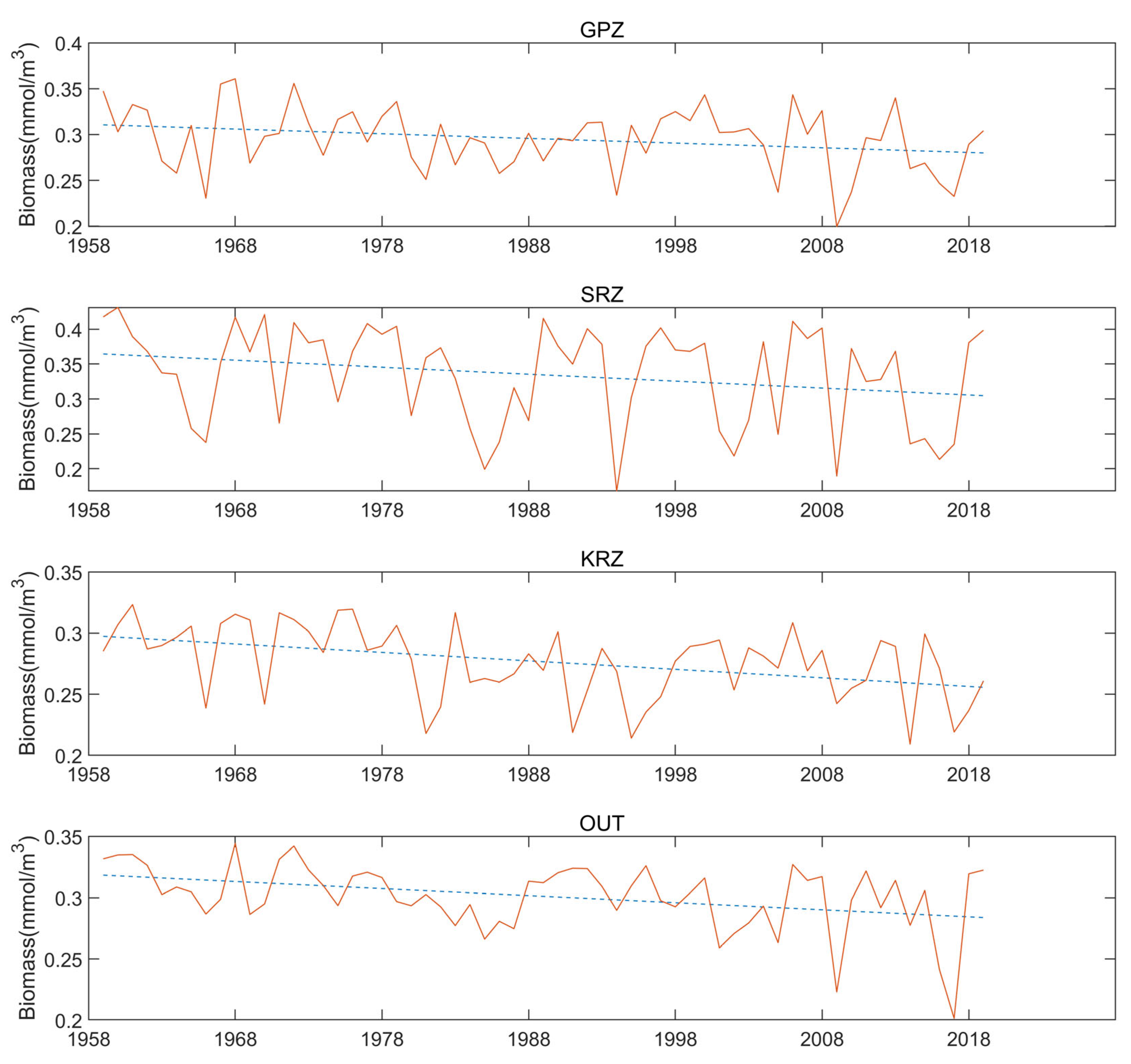
3.2. The Interannual Variation Trend in February
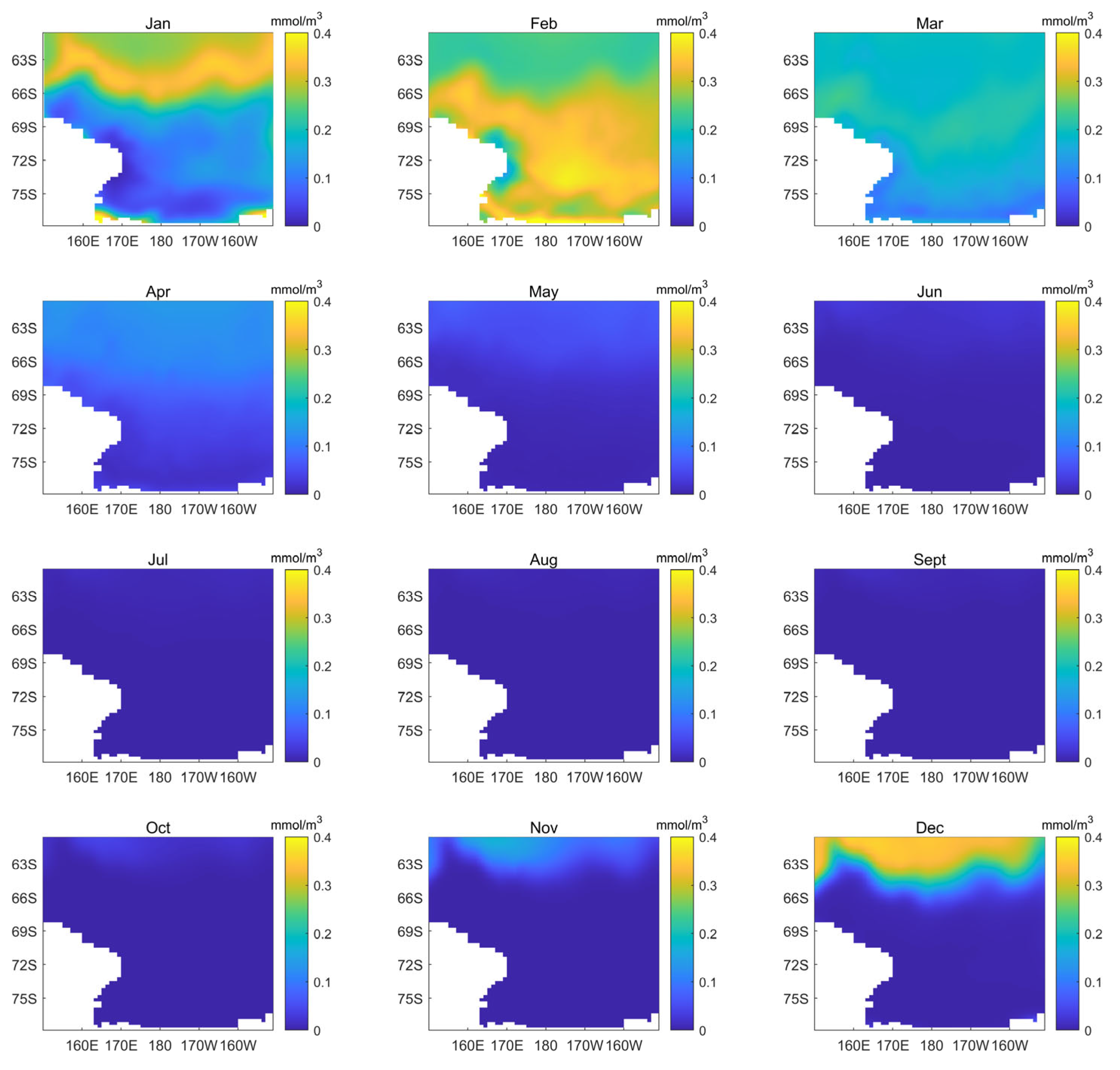
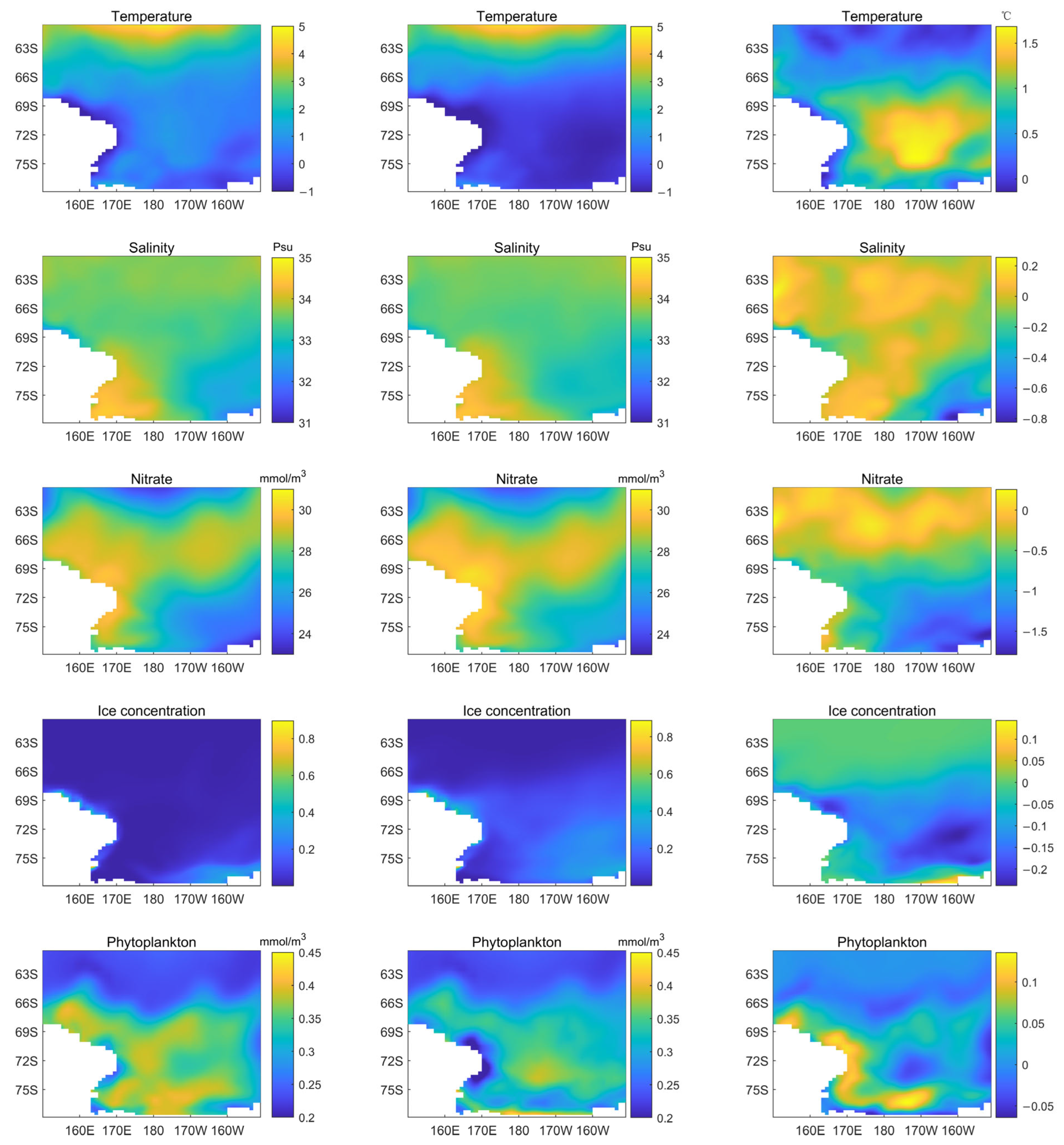
3.3. Spatial Variability in Phytoplankton Biomass and Other Environmental Variables in the Ross Sea
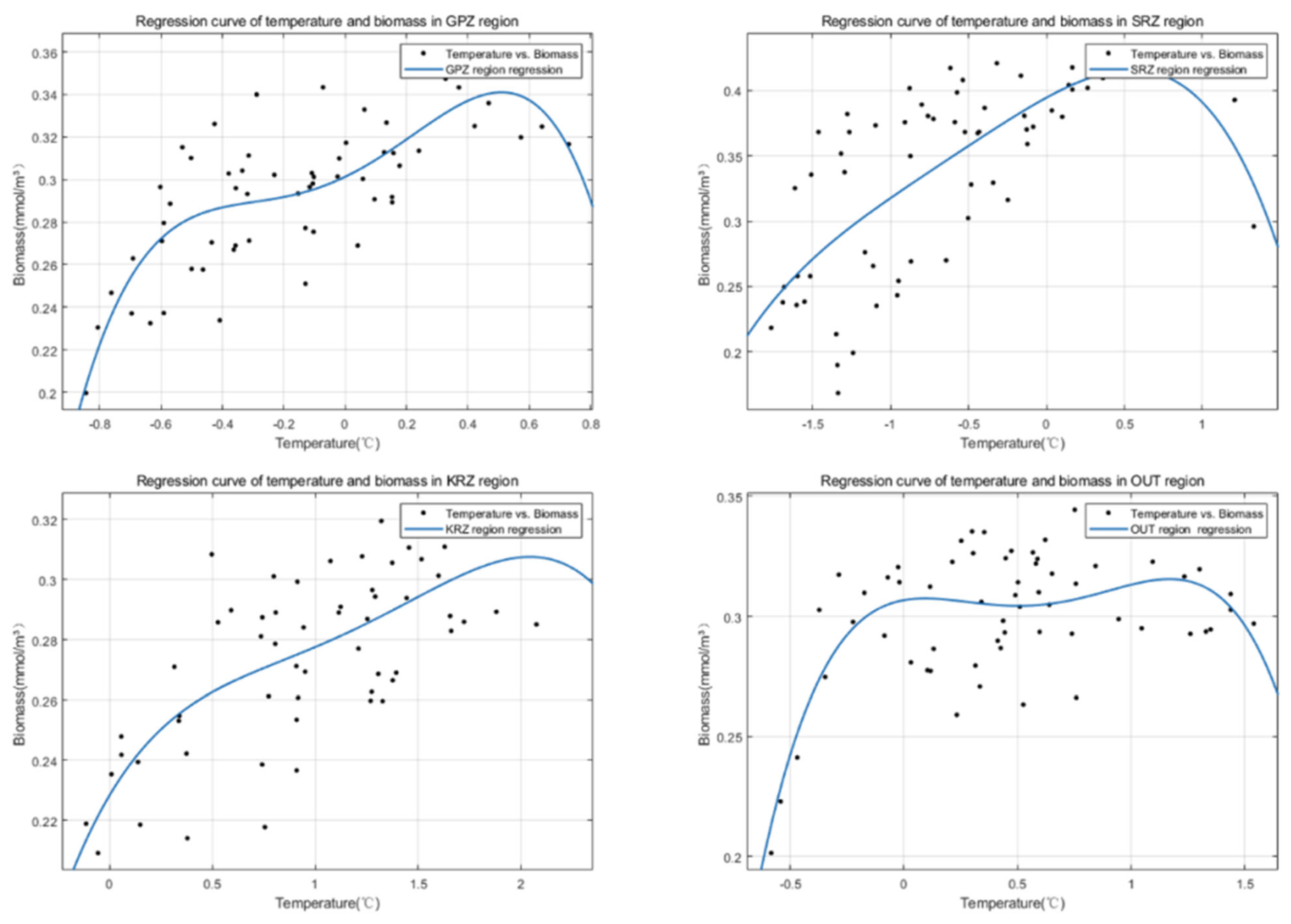
3.4. Lag Correlation Analysis
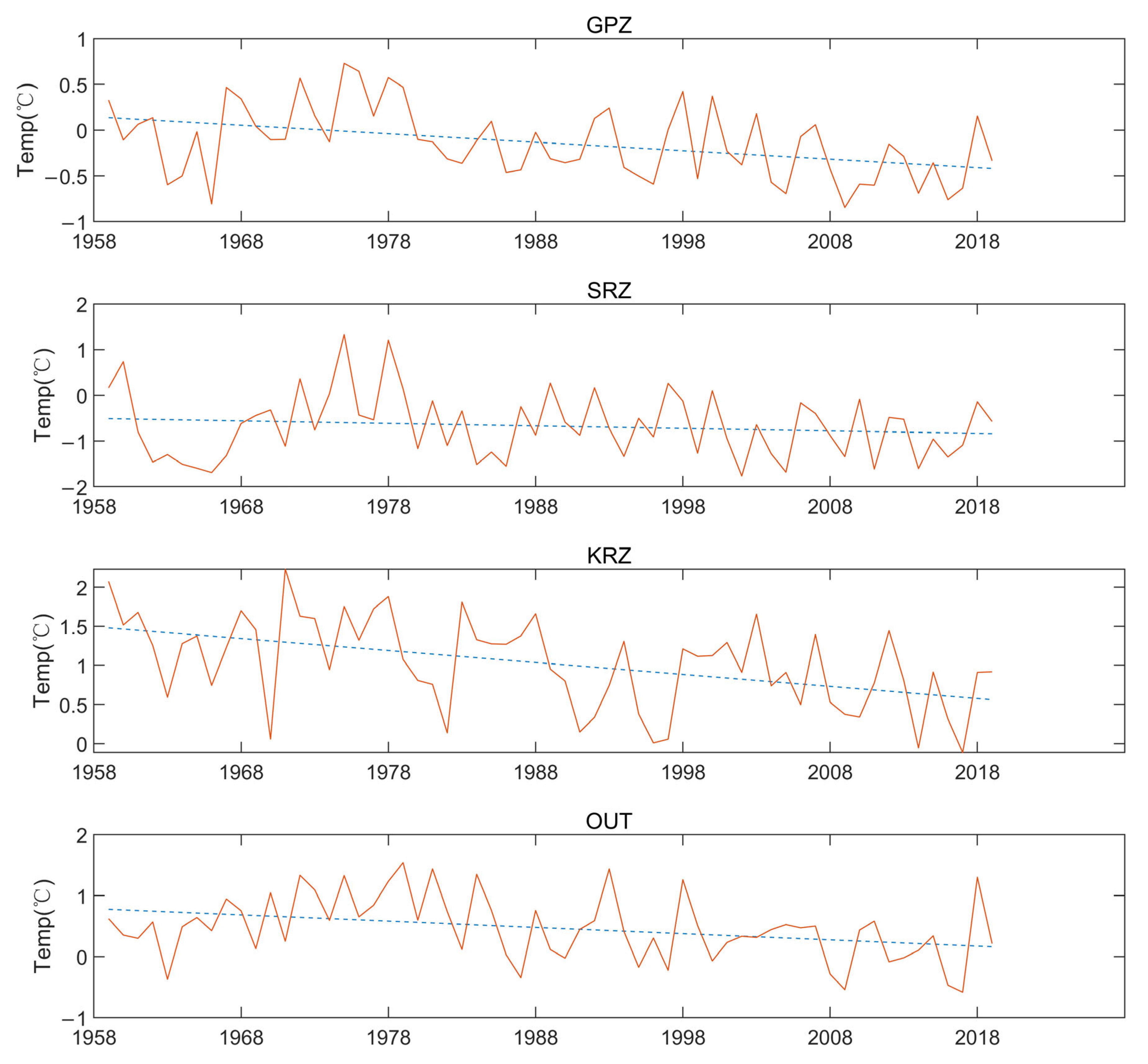
3.5. Years with Unusually High Temperatures
4. Discussion
4.1. Limitation and Plan
4.2. Lack of Perceived Temperature Change in the Ross Sea
4.3. Changes in Sea Ice in the Southern Ocean and Ecological Impacts, Interactions in the Ecosystem of the Ross Sea Shelf
4.4. The Future of the Ross Sea MPA Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Waterhouse, E.J. (Ed.) Ross Sea Region 2001: A State of the Environment Report for the Ross Sea Region of Antarctica; New Zealand Antarctic Inst.: Christchurch, New Zealand, 2001. [Google Scholar]
- Beer, K.; Brears, R.; Briars, L.; Roldan, G. A Pragmatic Utopia: Should the Ross Sea be Designated a Marine Protected Area? University of Canterbury: Christchurch, New Zealand, 2011. [Google Scholar]
- Silvano, A.; Foppert, A.; Rintoul, S.R.; Holland, P.R.; Tamura, T.; Kimura, N.; Macdonald, A.M. Recent recovery of Antarctic Bottom Water formation in the Ross Sea driven by climate anomalies. Nat. Geosci. 2020, 13, 780–786. [Google Scholar] [CrossRef]
- Castagno, P.; Capozzi, V.; DiTullio, G.R.; Falco, P.; Fusco, G.; Rintoul, S.R.; Budillon, G. Rebound of shelf water salinity in the Ross Sea. Nat. Commun. 2019, 10, 5441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, C.M.; Bloom, E.; Kavanagh, A.; Nocito, E.S.; Watters, G.M.; Weller, J. The Ross Sea, Antarctica: A highly protected MPA in international waters. Mar. Policy 2021, 134, 104795. [Google Scholar] [CrossRef]
- Eastman, J.T. The nature of the diversity of Antarctic fishes. Polar Biol. 2005, 28, 93–107. [Google Scholar] [CrossRef]
- Smith, W.O., Jr.; Ainley, D.G.; Cattaneo-Vietti, R. Trophic interactions within the Ross Sea continental shelf ecosystem. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Schine, C.M.; van Dijken, G.; Arrigo, K.R. Spatial analysis of trends in primary production and relationship with large-scale climate variability in the Ross Sea, Antarctica (1997–2013). J. Geophys. Res. Ocean. 2016, 121, 368–386. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, C.L.; Cavalieri, D.J. Antarctic sea ice variability and trends, 1979–2010. Cryosphere 2012, 6, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Cavalieri, D.J.; Parkinson, C.L.; Gloersen, P.; Comiso, J.C.; Zwally, H.J. Deriving long-term time series of sea ice cover from satellite passive-microwave multisensor data sets. J. Geophys. Res. Ocean. 1999, 104, 15803–15814. [Google Scholar] [CrossRef]
- Hobbs, W.R.; Massom, R.; Stammerjohn, S.; Reid, P.; Williams, G.; Meier, W. A review of recent changes in Southern Ocean sea ice, their drivers and forcings. Glob. Planet. Chang. 2016, 143, 228–250. [Google Scholar] [CrossRef] [Green Version]
- Deppeler, S.L.; Davidson, A.T. Southern Ocean phytoplankton in a changing climate. Front. Mar. Sci. 2017, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Hückstädt, L.A.; Piñones, A.; Palacios, D.M.; McDonald, B.I.; Dinniman, M.S.; Hofmann, E.E.; Costa, D.P. Projected shifts in the foraging habitat of crabeater seals along the Antarctic Peninsula. Nat. Clim. Chang. 2020, 10, 472–477. [Google Scholar] [CrossRef]
- Mitchell, B.G.; Brody, E.A.; Holm-Hansen, O.; McClain, C.; Bishop, J. Light limitation of phytoplankton biomass and macronutrient utilization in the Southern Ocean. Limnol. Oceanogr. 1991, 36, 1662–1677. [Google Scholar] [CrossRef]
- Martin, J.H.; Fitzwater, S.E.; Gordon, R.M. Iron deficiency limits phytoplankton growth in Antarctic waters. Glob. Biogeochem. Cycles 1990, 4, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Charlson, R.J.; Lovelock, J.E.; Andreae, M.O.; Warren, S.G. Oceanic phytoplankton, atmospheric sulphur, cloud albedo and climate. Nature 1987, 326, 655–661. [Google Scholar] [CrossRef]
- Meiners, K.M.; Vancoppenolle, M.; Thanassekos, S.; Dieckmann, G.S.; Thomas, D.N.; Tison, J.L.; Raymond, B. Chlorophyll a in Antarctic sea ice from historical ice core data. Geophys. Res. Lett. 2012, 39, L21602. [Google Scholar] [CrossRef]
- Miller, C.B. Biological Oceanography; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Smith, W.O.; Nelson, D.M. Phytoplankton bloom produced by a receding ice edge in the Ross Sea: Spatial coherence with the density field. Science 1985, 227, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Lizotte, M.P. The contributions of sea ice algae to Antarctic marine primary production. Am. Zool. 2001, 41, 57–73. [Google Scholar] [CrossRef]
- Smith, W.O., Jr.; Nelson, D.M.; DiTullio, G.R.; Leventer, A.R. Temporal and spatial patterns in the Ross Sea: Phytoplankton biomass, elemental composition, productivity and growth rates. J. Geophys. Res. Ocean. 1996, 101, 18455–18465. [Google Scholar] [CrossRef]
- Boyd, P.W.; Arrigo, K.R.; Strzepek, R.; Van Dijken, G.L. Mapping phytoplankton iron utilization: Insights into Southern Ocean supply mechanisms. J. Geophys. Res. Ocean. 2012, 117, C06009. [Google Scholar] [CrossRef] [Green Version]
- Strzepek, R.F.; Maldonado, M.T.; Hunter, K.A.; Frew, R.D.; Boyd, P.W. Adaptive strategies by Southern Ocean phytoplankton to lessen iron limitation: Uptake of organically complexed iron and reduced cellular iron requirements. Limnol. Oceanogr. 2011, 56, 1983–2002. [Google Scholar] [CrossRef] [Green Version]
- De Baar, H.J.; de Jong, J.T.; Bakker, D.C.; Löscher, B.M.; Veth, C.; Bathmann, U.; Smetacek, V. Importance of iron for plankton blooms and carbon dioxide drawdown in the Southern Ocean. Nature 1995, 373, 412–415. [Google Scholar] [CrossRef]
- Sohrin, Y.; Iwamoto, S.I.; Matsui, M.; Obata, H.; Nakayama, E.; Suzuki, K.; Ishii, M. The distribution of Fe in the Australian sector of the Southern Ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2000, 47, 55–84. [Google Scholar] [CrossRef]
- Kiss, A.E.; Hogg, A.M.; Hannah, N.; Boeira Dias, F.; Brassington, G.B.; Chamberlain, M.A.; Chapman, C.; Dobrohotoff, P.; Domingues, C.M.; Duran, E.R.; et al. ACCESS-OM2 v1.0: A global ocean–sea ice model at three resolutions. Geosci. Model Dev. 2020, 13, 401–442. [Google Scholar]
- Schiller, A.; Oke, P.R. BLUElink> Development of operational oceanography and servicing in Australia. J. Res. Pract. Inf. Technol. 2007, 39, 151–164. [Google Scholar]
- Kidston, M.; Matear, R.; Baird, M.E. Parameter optimisation of a marine ecosystem model at two contrasting stations in the Sub-Antarctic Zone. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 2301–2315. [Google Scholar] [CrossRef]
- Oke, P.R.; Griffin, D.A.; Schiller, A.; Matear, R.J.; Fiedler, R.; Mansbridge, J.; Ridgway, K. Evaluation of a near-global eddy-resolving ocean model. Geosci. Model Dev. 2013, 6, 591–615. [Google Scholar] [CrossRef] [Green Version]
- Heinle, A.; Slawig, T. Internal dynamics of NPZD type ecosystem models. Ecol. Model. 2013, 254, 33–42. [Google Scholar] [CrossRef]
- Xue, P.; Schwab, D.J.; Zhou, X.; Huang, C.; Kibler, R.; Ye, X. A hybrid lagrangian–eulerian particle model for ecosystem simulation. J. Mar. Sci. Eng. 2018, 6, 109. [Google Scholar] [CrossRef] [Green Version]
- Oschlies, A.; Garçon, V. An eddy-permitting coupled physical-biological model of the North Atlantic: 1. Sensitivity to advection numerics and mixed layer physics. Glob. Biogeochem. Cycles 1999, 13, 135–160. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.O., Jr.; Marra, J.; Hiscock, M.R.; Barber, R.T. The seasonal cycle of phytoplankton biomass and primary productivity in the Ross Sea, Antarctica. Deep Sea Res. Part II Top. Stud. Oceanogr. 2000, 47, 3119–3140. [Google Scholar] [CrossRef]
- Griffies, S.M.; Winton, M.; Anderson, W.G.; Benson, R.; Delworth, T.L.; Dufour, C.O.; Zhang, R. Impacts on ocean heat from transient mesoscale eddies in a hierarchy of climate models. J. Clim. 2015, 28, 952–977. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, H.T.; Roberts, M.J.; Hyder, P.; Graham, T.; Rae, J.; Belcher, S.E.; Wood, R.A. The impact of resolving the Rossby radius at mid-latitudes in the ocean: Results from a high-resolution version of the Met Office GC2 coupled model. Geosci. Model Dev. 2016, 9, 3655–3670. [Google Scholar] [CrossRef] [Green Version]
- Bishop, S.P.; Gent, P.R.; Bryan, F.O.; Thompson, A.F.; Long, M.C.; Abernathey, R. Southern Ocean overturning compensation in an eddy-resolving climate simulation. J. Phys. Oceanogr. 2016, 46, 1575–1592. [Google Scholar] [CrossRef]
- Eppley, R.W. Temperature and phytoplankton growth in the sea. Fish. Bull 1972, 70, 1063–1085. [Google Scholar]
- Monod, J. The growth of bacterial cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef] [Green Version]
- Hayashida, H.; Matear, R.J.; Strutton, P.G. Background nutrient concentration determines phytoplankton bloom response to marine heatwaves. Glob. Chang. Biol. 2020, 26, 4800–4811. [Google Scholar] [CrossRef]
- Armour, K.C.; Marshall, J.; Scott, J.R.; Donohoe, A.; Newsom, E.R. Southern Ocean warming delayed by circumpolar upwelling and equatorward transport. Nat. Geosci. 2016, 9, 549–554. [Google Scholar] [CrossRef]
- Convey, P.; Bindschadler, R.; di Prisco, G.; Fahrbach, E.; Gutt, J.; Hodgson, D.; Mayewski, P.; Summerhayes, C.; Turner, J. Antarctic climate change and the environment. Antarct. Sci. 2009, 21, 541–563. [Google Scholar] [CrossRef] [Green Version]
- Holland, P.R.; Jenkins, A.; Holland, D.M. Ice and ocean processes in the Bellingshausen Sea, Antarctica. J. Geophys. Res. Ocean. 2010, 115, C05020. [Google Scholar] [CrossRef] [Green Version]
- St-Laurent, P.; Klinck, J.M.; Dinniman, M.S. Impact of local winter cooling on the melt of Pine Island Glacier, Antarctica. J. Geophys. Res. Ocean. 2015, 120, 6718–6732. [Google Scholar] [CrossRef] [Green Version]
- Dutrieux, P.; De Rydt, J.; Jenkins, A.; Holland, P.R.; Ha, H.K.; Lee, S.H.; Schröder, M. Strong sensitivity of Pine Island ice-shelf melting to climatic variability. Science 2014, 343, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Agosta, C.; Fettweis, X.; Datta, R. Evaluation of the CMIP5 models with the aim of regional modelling of the Antarctic surface mass balance. Cryosphere 2015, 9, 2311–2321. [Google Scholar] [CrossRef] [Green Version]
- Bracegirdle, T.J.; Stephenson, D.B.; Turner, J.; Phillips, T. The importance of sea ice area biases in 21st century multimodel projections of Antarctic temperature and precipitation. Geophys. Res. Lett. 2015, 42, 10–832. [Google Scholar] [CrossRef] [Green Version]
- Kennicutt, M.C.; Chown, S.L.; Cassano, J.J.; Liggett, D.; Peck, L.S.; Massom, R.; Sutherland, W.J. A roadmap for Antarctic and Southern Ocean science for the next two decades and beyond. Antarct. Sci. 2015, 27, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Mezgec, K.; Stenni, B.; Crosta, X.; Masson-Delmotte, V.; Baroni, C.; Braida, M.; Frezzotti, M. Holocene sea ice variability driven by wind and polynya efficiency in the Ross Sea. Nat. Commun. 2017, 8, 1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massom, R.A.; Stammerjohn, S.E. Antarctic sea ice change and variability–physical and ecological implications. Polar Sci. 2010, 4, 149–186. [Google Scholar] [CrossRef] [Green Version]
- Frazer, T.K.; Quetin, L.B.; Ross, R.M. Energetic demands of larval krill, Euphausia superba, in winter. J. Exp. Mar. Biol. Ecol. 2002, 277, 157–171. [Google Scholar] [CrossRef]
- Quetin, L.B.; Ross, R.M. Life under Antarctic pack ice: A krill perspective. In Smithsonian at the Poles: Contributions to International Polar Year Science; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2009. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Symbol |
|---|---|
| Quadratic mortality rate (Phytoplankton coefficients) | |
| Remineralization rate (Detrital coefficients) | |
| Quadratic mortality rate (Zooplankton coefficients) | |
| Sinking velocity (Detrital coefficients) | |
| Assimilation efficiency (Zooplankton coefficients) | |
| Excretion rate (Zooplankton coefficients) | |
| The nitrogen concentration below the mixed layer |
| Temperature | Salinity | Ice | Nitrate | |||||
|---|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | r | p-Value | |
| GPZ | 0.723 | 0.000 | −0.217 | 0.093 | −0.534 | 0.000 | −0.553 | 0.000 |
| SRZ | 0.607 | 0.000 | −0.210 | 0.105 | −0.578 | 0.000 | −0.541 | 0.000 |
| KRZ | 0.711 | 0.000 | 0.156 | 0.230 | −0.590 | 0.000 | −0.168 | 0.196 |
| OUT | 0.383 | 0.002 | −0.407 | 0.001 | −0.539 | 0.000 | −0.405 | 0.001 |
| Temperature | Salinity | Ice | Nitrate | |||||
|---|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | r | p-Value | |
| GPZ | 0.639 | 0.000 | −0.439 | 0.000 | −0.586 | 0.000 | −0.332 | 0.009 |
| SRZ | 0.485 | 0.000 | −0.345 | 0.006 | −0.582 | 0.000 | −0.243 | 0.059 |
| KRZ | 0.651 | 0.000 | −0.057 | 0.661 | −0.713 | 0.000 | −0.010 | 0.940 |
| OUT | 0.299 | 0.020 | −0.536 | 0.000 | −0.419 | 0.000 | −0.238 | 0.065 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Lv, X. Study of Phytoplankton Biomass and Environmental Drivers in and around the Ross Sea Marine Protected Area. J. Mar. Sci. Eng. 2023, 11, 747. https://doi.org/10.3390/jmse11040747
Song Y, Lv X. Study of Phytoplankton Biomass and Environmental Drivers in and around the Ross Sea Marine Protected Area. Journal of Marine Science and Engineering. 2023; 11(4):747. https://doi.org/10.3390/jmse11040747
Chicago/Turabian StyleSong, Yangjinan, and Xianqing Lv. 2023. "Study of Phytoplankton Biomass and Environmental Drivers in and around the Ross Sea Marine Protected Area" Journal of Marine Science and Engineering 11, no. 4: 747. https://doi.org/10.3390/jmse11040747