
MicroRNA-Mediated Responses: Adaptations to Marine Extreme Environments



Abstract
:1. Introduction
1.1. Organism Metabolic Adaptations
1.2. Cellular Adaptations
2. RNA-Mediated Gene Silencing: A Way to Adapt to the Environment
2.1. Gene Silencing: The Role of Non-Coding RNAs
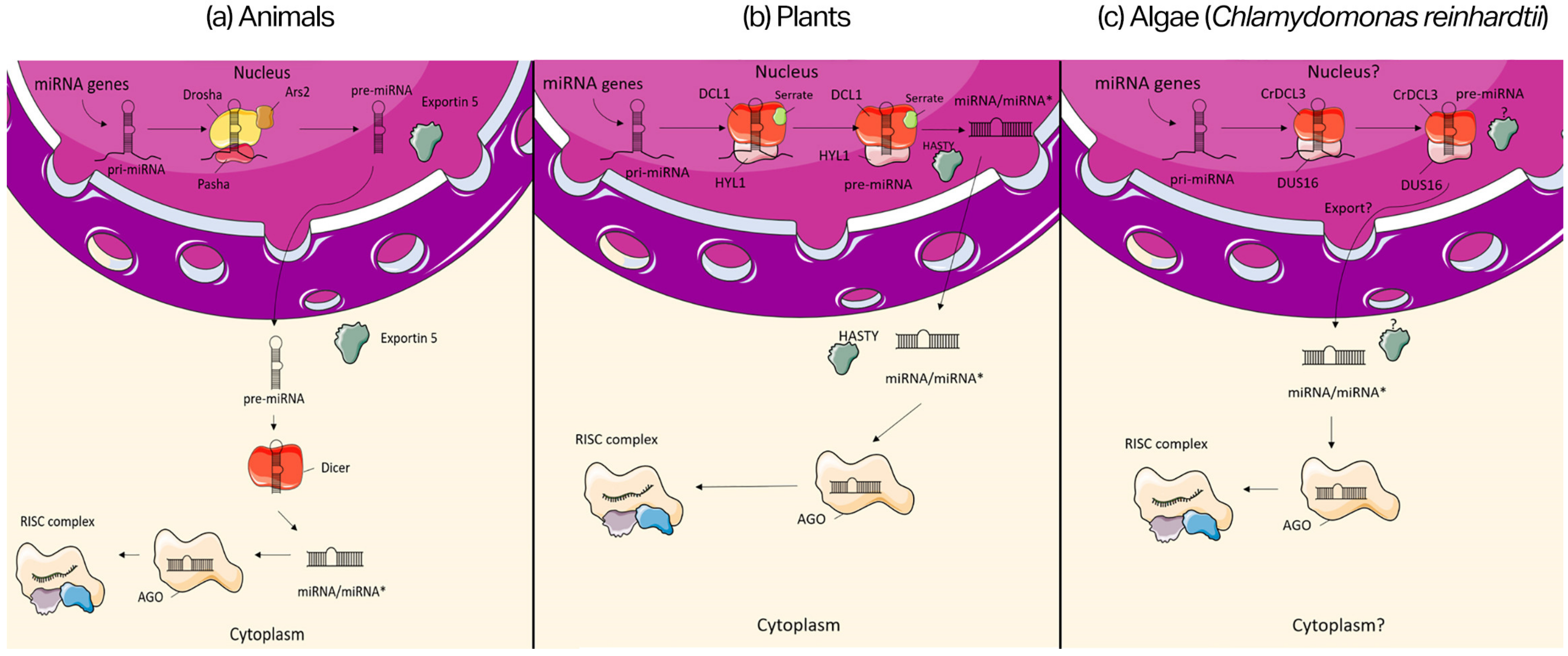
2.2. miRNA Biogenesis
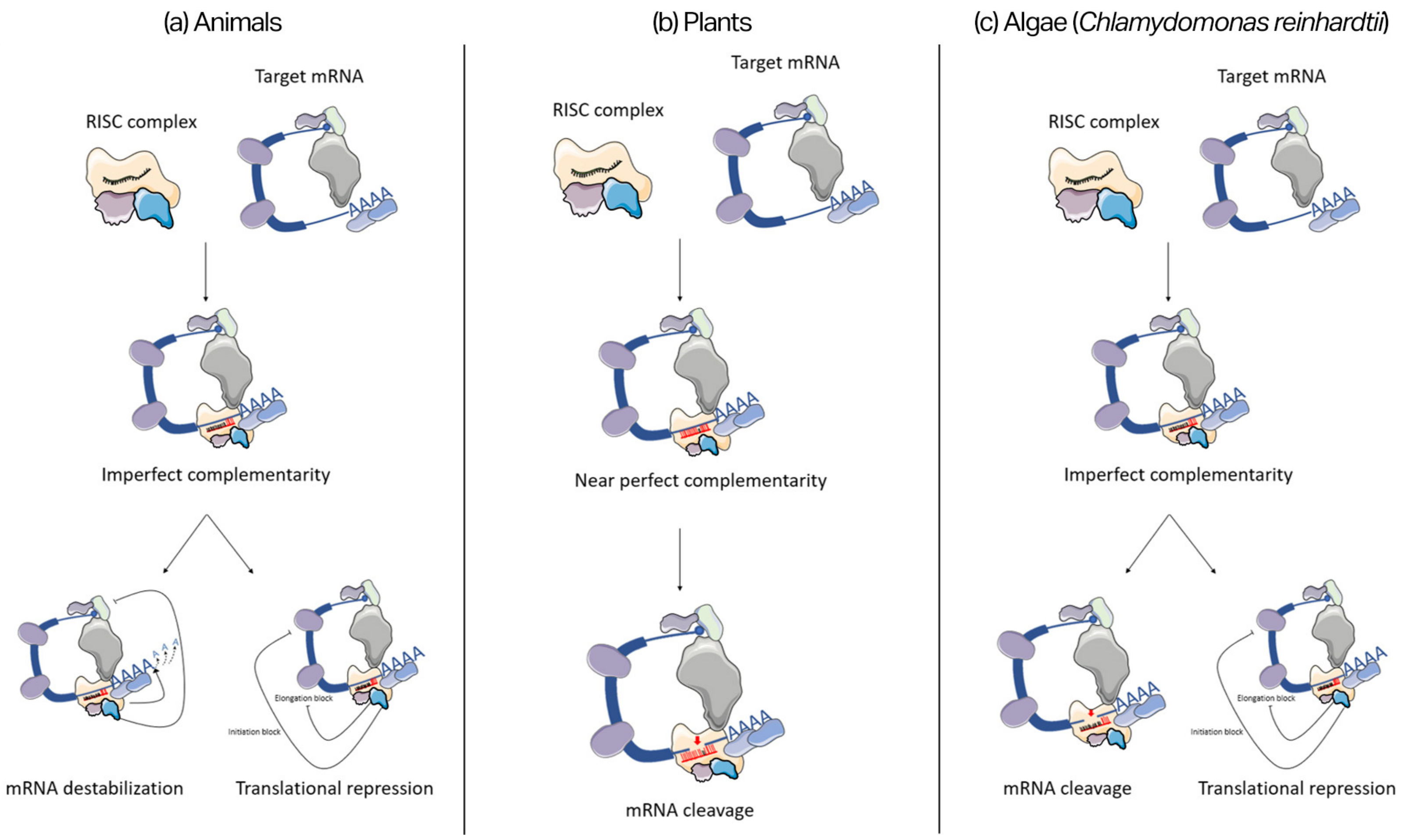
2.3. miRNA Mode of Action
2.4. miRNA Conservation
3. miRNA-Mediated Adaptations to Marine Extreme Conditions
3.1. Organisms Experiencing Extreme Conditions during Their Lifetime

{kind=link}
{kind=link}
| Species | Condition | DE miRNAs | Adaptations | References |
|---|---|---|---|---|
| Dosidicus gigas | Hypoxia and freezing | miR-1175; miR-133, miR-33; miR-67; miR-29; miR-2a; miR-100; miR-12; miR-1985; miR-2001; miR-2722; miR-190; miR-34 | Hypometabolic state; anti-apoptotic responses; reduction of oxidative stress; modulation of cardiac hypertrophy | [77] |
| Hemiscyllium ocellatum | Hypoxia and anoxia | miR-92; miR181a; miR-146b; miR-140; miR-20a; miR-17; miR-138; miR-143 | Metabolic rate depression; anti-ischemic responses; modulation of HIF; anti-apoptotic responses | [88] |
| Apostichopus japonicus | Hypoxia | Aja-miR-2008,; Aja-miR-10-5p; Aja-miR-184; Aja-miR-71b; Aja-miR-125-5p; novel-miR-1; Aja-let-7a-5p; Aja-miR-375-3p; Aja-miR-2013-3p; novel-miR-2; Aja-miR-2835; Aja-miR-1; Aja-miR-71-5p; Aja-miR-200-3p; Aja-miR-2011-3p; Aja-miR-2478a; Aja-miR-31-5p; Aja-miR-7977; Aja-miR-71a; Aja-miR-29b-3p; Aja-miR-2478b; Aja-miR-2008-5p; Aja-miR-1a-3p; novel-miR-3; Aja-miR-153-3p; Aja-miR-153 | Negatively regulate HIF pathway; reduction of cellular autophagy; induction of cell cycle arrest | [94] |
| Littorina littorea | Anoxia and freezing | miR-1a; miR-210; miR-34a; miR-133a; miR-125b; miR-29b; miR-2a | Hypometabolic state; anti- apoptotic responses; reduction of protein synthesis; activation of oxidative stress response pathways | [98] |
| Trematomus bernacchi | Heat stress | tbe-miR-22a; tbe-let-7; tbe-miR-21; tbe-let-7a; tbe-miR26a; tbe-miR30a; tbe-miR-146a; tbe-miR-203b; tbe-miR-200a; tbe-miR-725 | Regulation of: FoxO signaling cascade, TLR pathway, PI3KT-AKT signaling pathway. Anti-apoptotic responses | [106] |
| Dunaliella salina | High salinity and light intensity | miR-482; miR-162; miR-3630; miR-166; miR-858; novel-m0038-5p; novel-m0783-5p; novel-m1007-3p; novel-m0533-3p | Accumulation of antioxidants (β-carotene) | [113] |
3.2. Identification of miRNAs through Stress-Response Laboratory Experiments
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindgren, A.R.; Buckley, B.A.; Eppley, S.M.; Reysenbach, A.-L.; Stedman, K.M.; Wagner, J.T. Life on the Edge—The Biology of Organisms Inhabiting Extreme Environments: An Introduction to the Symposium. Integr. Comp. Biol. 2016, 56, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, L.J.; Mancinelli, R.L. Life in Extreme Environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Bijma, J.; Pörtner, H.-O.; Yesson, C.; Rogers, A.D. Climate Change and the Oceans—What Does the Future Hold? Mar. Pollut. Bull. 2013, 74, 495–505. [Google Scholar] [CrossRef]
- Schmidtko, S.; Stramma, L.; Visbeck, M. Decline in Global Oceanic Oxygen Content during the Past Five Decades. Nature 2017, 542, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Camp, E.F.; Schoepf, V.; Mumby, P.J.; Hardtke, L.A.; Rodolfo-Metalpa, R.; Smith, D.J.; Suggett, D.J. The Future of Coral Reefs Subject to Rapid Climate Change: Lessons from Natural Extreme Environments. Front. Mar. Sci. 2018, 5, 4. [Google Scholar] [CrossRef]
- Macelroy, R.D. Some Comments on the Evolution of Extremophiles. Biosystems 1974, 6, 74–75. [Google Scholar] [CrossRef]
- Morita, R.Y. Extremes of Biodiversity. BioScience 1999, 49, 245–248. [Google Scholar] [CrossRef]
- Lévêque, E.; Janeček, Š.; Haye, B.; Belarbi, A. Thermophilic Archaeal Amylolytic Enzymes. Enzyme Microb. Technol. 2000, 26, 3–14. [Google Scholar] [CrossRef]
- Driedzic, W.R.; Clow, K.A.; Short, C.E.; Ewart, K.V. Glycerol Production in Rainbow Smelt (Osmerus mordax) May Be Triggered by Low Temperature Alone and Is Associated with the Activation of Glycerol-3-Phosphate Dehydrogenase and Glycerol-3-Phosphatase. J. Exp. Biol. 2006, 209, 1016–1023. [Google Scholar] [CrossRef]
- Seckbach, J. Enigmatic Microorganisms and Life in Extreme Environments; Springer: Dordrecht, The Netherlands, 2013; ISBN 978-94-011-4838-2. [Google Scholar]
- Chilukuri, L.N.; Bartlett, D.H. Isolation and Characterization of the Gene Encoding Single-Stranded-DNA-Binding Protein (SSB) from Four Marine Shewanella Strains That Differ in Their Temperature and Pressure Optima for Growth. Microbiology 1997, 143, 1163–1174. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, R.G. Limnology: Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA, 2001; ISBN 978-0-12-744760-5. [Google Scholar]
- Storey, K.B.; Storey, J.M. Biochemical Adaptation to Extreme Environments. In Integrative Physiology in the Proteomics and Post-Genomics Age; Walz, W., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 169–200. ISBN 978-1-58829-315-2. [Google Scholar]
- Storey, K.B.; Storey, J.M. Tribute to P. L. Lutz: Putting Life on ‘pause’—Molecular Regulation of Hypometabolism. J. Exp. Biol. 2007, 210, 1700–1714. [Google Scholar] [CrossRef]
- Heldmaier, G.; Ortmann, S.; Elvert, R. Natural Hypometabolism during Hibernation and Daily Torpor in Mammals. Resp. Physiol. Neurobiol. 2004, 141, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution; Oxford University Press: New York, NY, USA, 2002; ISBN 978-0-19-511702-8. [Google Scholar]
- Storey, K.B. Life in the Slow Lane: Molecular Mechanisms of Estivation. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 133, 733–754. [Google Scholar] [CrossRef] [PubMed]
- Lutz, P.L.; Nilsson, G.E. Vertebrate Brains at the Pilot Light. Resp. Physiol. Neurobiol. 2004, 141, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, R.; Varacallo, M. Biochemistry, Glycolysis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Akram, M. Mini-Review on Glycolysis and Cancer. J. Cancer Educ. 2013, 28, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory Metabolism: Glycolysis, the TCA Cycle and Mitochondrial Electron Transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef]
- Choi, I.; Son, H.; Baek, J.-H. Tricarboxylic Acid (TCA) Cycle Intermediates: Regulators of Immune Responses. Life 2021, 11, 69. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Regulation in Photosynthetic Organisms: Signaling, Acclimation, and Practical Implications. Antioxid. Redox Signal 2009, 11, 861–905. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B. How to Characterize an Antioxidant: An Update. Biochem. Soc. Symp. 1995, 61, 73–101. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Baky, H.H.; El Baz, F.K.; El-Baroty, G.S. Enhancement of Antioxidant Production in Spirulina platensis under Oxidative Stress. Acta Physiol. Plant. 2009, 31, 623–631. [Google Scholar] [CrossRef]
- Akhtar, T.A.; Lees, H.A.; Lampi, M.A.; Enstone, D.; Brain, R.A.; Greenberg, B.M. Photosynthetic Redox Imbalance Influences Flavonoid Biosynthesis in Lemna gibba. Plant Cell Environ. 2010, 33, 1205–1219. [Google Scholar] [CrossRef]
- Agati, G.; Tattini, M. Multiple Functional Roles of Flavonoids in Photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Matteini, P.; Goti, A.; Tattini, M. Chloroplast-located Flavonoids Can Scavenge Singlet Oxygen. New Phytol. 2007, 174, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Gould, K.S.; McKelvie, J.; Markham, K.R. Do Anthocyanins Function as Antioxidants in Leaves? Imaging of H2O2 in Red and Green Leaves after Mechanical Injury: H2O2 Scavenging by Anthocyanins. Plant Cell Environ. 2002, 25, 1261–1269. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as Natural Functional Pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef]
- Remias, D.; Karsten, U.; Lütz, C.; Leya, T. Physiological and Morphological Processes in the Alpine Snow Alga Chloromonas nivalis (Chlorophyceae) during Cyst Formation. Protoplasma 2010, 243, 73–86. [Google Scholar] [CrossRef]
- Doppler, P.; Kornpointner, C.; Halbwirth, H.; Remias, D.; Spadiut, O. Tetraedron Minimum, First Reported Member of Hydrodictyaceae to Accumulate Secondary Carotenoids. Life 2021, 11, 107. [Google Scholar] [CrossRef]
- Han, R.-M.; Zhang, J.-P.; Skibsted, L.H. Reaction Dynamics of Flavonoids and Carotenoids as Antioxidants. Molecules 2012, 17, 2140–2160. [Google Scholar] [CrossRef] [Green Version]
- Molino, A.; Iovine, A.; Casella, P.; Mehariya, S.; Chianese, S.; Cerbone, A.; Rimauro, J.; Musmarra, D. Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals. Int. J. Environ. Res. Public Health 2018, 15, 2436. [Google Scholar] [CrossRef]
- Yilancioglu, K.; Cokol, M.; Pastirmaci, I.; Erman, B.; Cetiner, S. Oxidative Stress Is a Mediator for Increased Lipid Accumulation in a Newly Isolated Dunaliella salina Strain. PLoS ONE 2014, 9, e91957. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z. Lipid Peroxidation. In Encyclopedia of Toxicology; Elsevier: Amsterdam, The Netherlands, 2005; pp. 730–734. ISBN 978-0-12-369400-3. [Google Scholar]
- Hartwell, L.H.; Weinert, T.A. Checkpoints: Controls That Ensure the Order of Cell Cycle Events. Science 1989, 246, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Pietenpol, J.A.; Stewart, Z.A. Cell Cycle Checkpoint Signaling. Toxicology 2002, 181–182, 475–481. [Google Scholar] [CrossRef]
- Joerger, A.C.; Fersht, A.R. The P53 Pathway: Origins, Inactivation in Cancer, and Emerging Therapeutic Approaches. Annu. Rev. Biochem. 2016, 85, 375–404. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and Regulation of Akt/PKB by the Rictor-MTOR Complex. Science 2005, 307, 1098–1101. [Google Scholar] [CrossRef]
- Mocellin, S.; Provenzano, M.M. RNA Interference: Learning Gene Knock-down from Cell Physiology. J. Transl. Med. 2004, 2, 39. [Google Scholar] [CrossRef]
- El-Sappah, A.H.; Yan, K.; Huang, Q.; Islam, M.M.; Li, Q.; Wang, Y.; Khan, M.S.; Zhao, X.; Mir, R.R.; Li, J.; et al. Comprehensive Mechanism of Gene Silencing and Its Role in Plant Growth and Development. Front. Plant Sci. 2021, 12, 705249. [Google Scholar] [CrossRef]
- Shabalina, S.; Koonin, E. Origins and Evolution of Eukaryotic RNA Interference. Trends Ecol. Evol. 2008, 23, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Higgs, P.G.; Lehman, N. The RNA World: Molecular Cooperation at the Origins of Life. Nat. Rev. Genet. 2015, 16, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional Regulation of the Heterochronic Gene Lin-14 by Lin-4 Mediates Temporal Pattern Formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Lou, S.; Sun, T.; Li, H.; Hu, Z. Mechanisms of MicroRNA-Mediated Gene Regulation in Unicellular Model Alga Chlamydomonas reinhardtii. Biotechnol. Biofuels 2018, 11, 244. [Google Scholar] [CrossRef] [PubMed]
- Bonin, C.A.; van Wijnen, A.J.; Lewallen, E.A. MicroRNA Applications in Marine Biology. Curr. Mol. Biol. Rep. 2019, 5, 167–175. [Google Scholar] [CrossRef]
- Wilson, M.D.; Wang, D.; Wagner, R.; Breyssens, H.; Gertsenstein, M.; Lobe, C.; Lu, X.; Nagy, A.; Burke, R.D.; Koop, B.F.; et al. ARS2 Is a Conserved Eukaryotic Gene Essential for Early Mammalian Development. Mol. Cell Biol. 2008, 28, 1503–1514. [Google Scholar] [CrossRef]
- Han, J.; Lee, Y.; Yeom, K.-H.; Kim, Y.-K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 Complex in Primary MicroRNA Processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef]
- Du, T.; Zamore, P.D. Beginning to Understand MicroRNA Function. Cell Res. 2007, 17, 661–663. [Google Scholar] [CrossRef]
- Park, M.Y.; Wu, G.; Gonzalez-Sulser, A.; Vaucheret, H.; Poethig, R.S. Nuclear Processing and Export of MicroRNAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 3691–3696. [Google Scholar] [CrossRef]
- Bollman, K.M.; Aukerman, M.J.; Park, M.-Y.; Hunter, C.; Berardini, T.Z.; Poethig, R.S. HASTY, the Arabidopsis ortholog of Exportin 5/MSN5, Regulates Phase Change and Morphogenesis. Development 2003, 130, 1493–1504. [Google Scholar] [CrossRef]
- Yamasaki, T.; Onishi, M.; Kim, E.-J.; Cerutti, H.; Ohama, T. RNA-Binding Protein DUS16 Plays an Essential Role in Primary MiRNA Processing in the Unicellular Alga Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2016, 113, 10720–10725. [Google Scholar] [CrossRef] [Green Version]
- Höck, J.; Meister, G. The Argonaute Protein Family. Genome Biol. 2008, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Rashid, U.J.; Paterok, D.; Koglin, A.; Gohlke, H.; Piehler, J.; Chen, J.C.-H. Structure of Aquifex Aeolicus Argonaute Highlights Conformational Flexibility of the PAZ Domain as a Potential Regulator of RNA-Induced Silencing Complex Function. J. Biol. Chem. 2007, 282, 13824–13832. [Google Scholar] [CrossRef] [PubMed]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of MicroRNA–Target Recognition. PLoS Biol. 2005, 3, e85. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, F.; Hentze, M.W. Molecular Mechanisms of Translational Control. Nat. Rev. Mol. Cell Biol. 2004, 5, 827–835. [Google Scholar] [CrossRef]
- Till, S.; Lejeune, E.; Thermann, R.; Bortfeld, M.; Hothorn, M.; Enderle, D.; Heinrich, C.; Hentze, M.W.; Ladurner, A.G. A Conserved Motif in Argonaute-Interacting Proteins Mediates Functional Interactions through the Argonaute PIWI Domain. Nat. Struct. Mol. Biol. 2007, 14, 897–903. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of Post-Transcriptional Regulation by MicroRNAs: Are the Answers in Sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Chendrimada, T.P.; Finn, K.J.; Ji, X.; Baillat, D.; Gregory, R.I.; Liebhaber, S.A.; Pasquinelli, A.E.; Shiekhattar, R. MicroRNA Silencing through RISC Recruitment of EIF6. Nature 2007, 447, 823–828. [Google Scholar] [CrossRef]
- Basu, U.; Si, K.; Warner, J.R.; Maitra, U. The Saccharomyces cerevisiae TIF6 Gene Encoding Translation Initiation Factor 6 Is Required for 60S Ribosomal Subunit Biogenesis. Mol. Cell. Biol. 2001, 21, 1453–1462. [Google Scholar] [CrossRef]
- Kim, J.; Krichevsky, A.; Grad, Y.; Hayes, G.D.; Kosik, K.S.; Church, G.M.; Ruvkun, G. Identification of Many MicroRNAs That Copurify with Polyribosomes in Mammalian Neurons. Proc. Natl. Acad. Sci. USA 2004, 101, 360–365. [Google Scholar] [CrossRef]
- Park, J.H.; Shin, C. MicroRNA-Directed Cleavage of Targets: Mechanism and Experimental Approaches. BMB Rep. 2014, 47, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Chung, B.Y.-W.; Deery, M.J.; Groen, A.J.; Howard, J.; Baulcombe, D.C. Endogenous MiRNA in the Green Alga Chlamydomonas Regulates Gene Expression through CDS-Targeting. Nat. Plants 2017, 3, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Dexheimer, P.J.; Cochella, L. MicroRNAs: From Mechanism to Organism. Front. Cell Dev. Biol. 2020, 8, 409. [Google Scholar] [CrossRef]
- Giraldez, A.J.; Cinalli, R.M.; Glasner, M.E.; Enright, A.J.; Thomson, J.M.; Baskerville, S.; Hammond, S.M.; Bartel, D.P.; Schier, A.F. MicroRNAs Regulate Brain Morphogenesis in Zebrafish. Science 2005, 308, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Vasquez-Rifo, A.; Jannot, G.; Armisen, J.; Labouesse, M.; Bukhari, S.I.A.; Rondeau, E.L.; Miska, E.A.; Simard, M.J. Developmental Characterization of the MicroRNA-Specific, C. elegans Argonautes Alg-1 and Alg-2. PLoS ONE 2012, 7, e33750. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, M.; Fujimi, M.; Iwamoto, C.; Onizuka, K.; Fukuda, N.; Ikeo, K.; Gojobori, T. Evolutionary Transitions of MicroRNA-Target Pairs. Genome Biol. Evol. 2016, 8, 1621–1633. [Google Scholar] [CrossRef]
- Coolen, M.; Katz, S.; Bally-Cuif, L. MiR-9: A Versatile Regulator of Neurogenesis. Front. Cell Neurosci. 2013, 7, 220. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, F.; Lee, J.-A.; Gao, F.-B. MicroRNA-9a Ensures the Precise Specification of Sensory Organ Precursors in Drosophila. Genes Dev. 2006, 20, 2793–2805. [Google Scholar] [CrossRef]
- Bagga, S.; Bracht, J.; Hunter, S.; Massirer, K.; Holtz, J.; Eachus, R.; Pasquinelli, A.E. Regulation by Let-7 and Lin-4 MiRNAs Results in Target MRNA Degradation. Cell 2005, 122, 553–563. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Müller, P.; et al. Conservation of the Sequence and Temporal Expression of Let-7 Heterochronic Regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Gilly, W.F.; Zeidberg, L.D.; Booth, J.A.T.; Stewart, J.S.; Marshall, G.; Abernathy, K.; Bell, L.E. Locomotion and Behavior of Humboldt Squid, Dosidicus gigas, in Relation to Natural Hypoxia in the Gulf of California, Mexico. J. Exp. Biol. 2012, 215, 3175–3190. [Google Scholar] [CrossRef] [Green Version]
- Hadj-Moussa, H.; Logan, S.M.; Seibel, B.A.; Storey, K.B. Potential Role for MicroRNA in Regulating Hypoxia-Induced Metabolic Suppression in Jumbo Squids. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiao, F.-Y.; Shan, P.-R.; Su, L.; Chen, D.-L.; Ding, J.-Y.; Wang, Z.-Q. Overexpression of MicroRNA-133a Inhibits Ischemia-Reperfusion-Induced Cardiomyocyte Apoptosis by Targeting DAPK2. J. Hum. Genet. 2015, 60, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Ber, Y.; Shiloh, R.; Gilad, Y.; Degani, N.; Bialik, S.; Kimchi, A. DAPK2 Is a Novel Regulator of MTORC1 Activity and Autophagy. Cell Death Differ. 2015, 22, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Näär, A.M. MiR-33: A Metabolic Conundrum. Trends Endocrinol. Metab. 2018, 29, 667–668. [Google Scholar] [CrossRef]
- Dávalos, A.; Goedeke, L.; Smibert, P.; Ramírez, C.M.; Warrier, N.P.; Andreo, U.; Cirera-Salinas, D.; Rayner, K.; Suresh, U.; Pastor-Pareja, J.C.; et al. MiR-33a/b Contribute to the Regulation of Fatty Acid Metabolism and Insulin Signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 9232–9237. [Google Scholar] [CrossRef]
- Ramírez, C.M.; Goedeke, L.; Rotllan, N.; Yoon, J.-H.; Cirera-Salinas, D.; Mattison, J.A.; Suárez, Y.; de Cabo, R.; Gorospe, M.; Fernández-Hernando, C. MicroRNA 33 Regulates Glucose Metabolism. Mol. Cell Biol. 2013, 33, 2891–2902. [Google Scholar] [CrossRef]
- Kriegel, A.J.; Baker, M.A.; Liu, Y.; Liu, P.; Cowley, A.W.; Liang, M. Endogenous MicroRNAs in Human Microvascular Endothelial Cells Regulate MRNAs Encoded by Hypertension-Related Genes. Hypertension 2015, 66, 793–799. [Google Scholar] [CrossRef]
- van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of MicroRNAs after Myocardial Infarction Reveals a Role of MiR-29 in Cardiac Fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef]
- Dong, D.; Yang, B. Role of MicroRNAs in Cardiac Hypertrophy, Myocardial Fibrosis and Heart Failure. Acta Pharm. Sin. 2011, 1, 1–7. [Google Scholar] [CrossRef]
- Dowd, W.W.; Renshaw, G.M.C.; Cech, J.J.; Kültz, D. Compensatory Proteome Adjustments Imply Tissue-Specific Structural and Metabolic Reorganization Following Episodic Hypoxia or Anoxia in the Epaulette Shark (Hemiscyllium ocellatum). Physiol. Genomics 2010, 42, 93–114. [Google Scholar] [CrossRef]
- Hadj-Moussa, H.; Storey, K.B. The OxymiR Response to Oxygen Limitation: A Comparative MicroRNA Perspective. J. Exp. Biol. 2020, 223, jeb204594. [Google Scholar] [CrossRef] [PubMed]
- Riggs, C.L.; Summers, A.; Warren, D.E.; Nilsson, G.E.; Lefevre, S.; Dowd, W.W.; Milton, S.; Podrabsky, J.E. Small Non-Coding RNA Expression and Vertebrate Anoxia Tolerance. Front. Genet. 2018, 9, 230. [Google Scholar] [CrossRef]
- Valera, V.A.; Walter, B.A.; Linehan, W.M.; Merino, M.J. Regulatory Effects of MicroRNA-92 (MiR-92) on VHL Gene Expression and the Hypoxic Activation of MiR-210 in Clear Cell Renal Cell Carcinoma. J. Cancer 2011, 2, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Greijer, A.E. The Role of Hypoxia Inducible Factor 1 (HIF-1) in Hypoxia Induced Apoptosis. J. Clin. Pathol. 2004, 57, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Di, Y.-F.; Li, D.-C.; Shen, Y.-Q.; Wang, C.-L.; Zhang, D.-Y.; Shang, A.-Q.; Hu, T. MiR-146b Protects Cardiomyocytes Injury in Myocardial Ischemia/Reperfusion by Targeting Smad4. Am. J. Transl. Res. 2017, 9, 656–663. [Google Scholar] [PubMed]
- Li, J.-W.; He, S.-Y.; Feng, Z.-Z.; Zhao, L.; Jia, W.-K.; Liu, P.; Zhu, Y.; Jian, Z.; Xiao, Y.-B. MicroRNA-146b Inhibition Augments Hypoxia-Induced Cardiomyocyte Apoptosis. Mol. Med. Rep. 2015, 12, 6903–6910. [Google Scholar] [CrossRef]
- Curtale, G.; Mirolo, M.; Renzi, T.A.; Rossato, M.; Bazzoni, F.; Locati, M. Negative Regulation of Toll-like Receptor 4 Signaling by IL-10–Dependent MicroRNA-146b. Proc. Natl. Acad. Sci. USA 2013, 110, 11499–11504. [Google Scholar] [CrossRef]
- Huo, D.; Sun, L.; Li, X.; Ru, X.; Liu, S.; Zhang, L.; Xing, L.; Yang, H. Differential Expression of MiRNAs in the Respiratory Tree of the Sea Cucumber Apostichopus japonicus Under Hypoxia Stress. G3 Genes Genomes Genet. 2017, 7, 3681–3692. [Google Scholar] [CrossRef]
- Liu, C.-J.; Tsai, M.-M.; Hung, P.-S.; Kao, S.-Y.; Liu, T.-Y.; Wu, K.-J.; Chiou, S.-H.; Lin, S.-C.; Chang, K.-W. MiR-31 Ablates Expression of the HIF Regulatory Factor FIH to Activate the HIF Pathway in Head and Neck Carcinoma. Cancer Res. 2010, 70, 1635–1644. [Google Scholar] [CrossRef]
- Chang, Y.; Yan, W.; He, X.; Zhang, L.; Li, C.; Huang, H.; Nace, G.; Geller, D.A.; Lin, J.; Tsung, A. MiR-375 Inhibits Autophagy and Reduces Viability of Hepatocellular Carcinoma Cells Under Hypoxic Conditions. Gastroenterology 2012, 143, 177–187.e8. [Google Scholar] [CrossRef]
- Liu, X.; Fu, B.; Chen, D.; Hong, Q.; Cui, J.; Li, J.; Bai, X.; Chen, X. MiR-184 and MiR-150 Promote Renal Glomerular Mesangial Cell Aging by Targeting Rab1a and Rab31. Exp. Cell Res. 2015, 336, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Biggar, K.K.; Kornfeld, S.F.; Maistrovski, Y.; Storey, K.B. MicroRNA Regulation in Extreme Environments: Differential Expression of MicroRNAs in the Intertidal Snail Littorina Littorea During Extended Periods of Freezing and Anoxia. Genom. Proteom. Bioinform. 2012, 10, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Churchill, T.A.; Storey, K.B. Metabolic Responses to Freezing and Anoxia by the Periwinkle Littorina littorea. J. Therm. Biol. 1996, 21, 57–63. [Google Scholar] [CrossRef]
- Devlin, C.; Greco, S.; Martelli, F.; Ivan, M. MiR-210: More than a Silent Player in Hypoxia. IUBMB Life 2011, 63, 94–100. [Google Scholar] [CrossRef]
- Li, N.; Cui, J.; Duan, X.; Chen, H.; Fan, F. Suppression of Type I Collagen Expression by MiR-29b via PI3K, Akt, and Sp1 Pathway in Human Tenon’s Fibroblasts. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1670. [Google Scholar] [CrossRef]
- Faridi, J.; Fawcett, J.; Wang, L.; Roth, R.A. Akt Promotes Increased Mammalian Cell Size by Stimulating Protein Synthesis and Inhibiting Protein Degradation. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E964–E972. [Google Scholar] [CrossRef]
- Ruggero, D.; Sonenberg, N. The Akt of Translational Control. Oncogene 2005, 24, 7426–7434. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, X.; Lim, L.P.; de Stanchina, E.; Xuan, Z.; Liang, Y.; Xue, W.; Zender, L.; Magnus, J.; Ridzon, D.; et al. A MicroRNA Component of the P53 Tumour Suppressor Network. Nature 2007, 447, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Yeung, S.J.; Pan, J.; Lee, M.-H. Roles of P53, Myc and HIF-1 in Regulating Glycolysis—The Seventh Hallmark of Cancer. Cell. Mol. Life Sci. 2008, 65, 3981–3999. [Google Scholar] [CrossRef]
- Vasadia, D.J.; Zippay, M.L.; Place, S.P. Characterization of Thermally Sensitive MiRNAs Reveals a Central Role of the FoxO Signaling Pathway in Regulating the Cellular Stress Response of an Extreme Stenotherm, Trematomus bernacchii. Mar. Genom. 2019, 48, 100698. [Google Scholar] [CrossRef]
- Bi, Y.; Liu, G.; Yang, R. MicroRNAs: Novel Regulators During the Immune Response. J. Cell Physiol. 2009, 218, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.R.; Yang, C.H.; Pfeffer, L.M. The Role of MiR-21 in Cancer: The Role of MiR-21 In Cancer. Drug Dev. Res. 2015, 76, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tang, N.; Hadden, T.J.; Rishi, A.K. Akt, FoxO and Regulation of Apoptosis. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 1978–1986. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Calcium Signalling and Cell Proliferation. Bioessays 1995, 17, 491–500. [Google Scholar] [CrossRef]
- Wei, S.; Bian, Y.; Zhao, Q.; Chen, S.; Mao, J.; Song, C.; Cheng, K.; Xiao, Z.; Zhang, C.; Ma, W.; et al. Salinity-Induced Palmella Formation Mechanism in Halotolerant Algae Dunaliella salina Revealed by Quantitative Proteomics and Phosphoproteomics. Front. Plant Sci. 2017, 8, 810. [Google Scholar] [CrossRef]
- Jeon, H.; Jeong, J.; Baek, K.; McKie-Krisberg, Z.; Polle, J.E.W.; Jin, E. Identification of the Carbonic Anhydrases from the Unicellular Green Alga Dunaliella salina Strain CCAP19/18. Algal Res. 2016, 19, 12–20. [Google Scholar] [CrossRef]
- Lou, S.; Zhu, X.; Zeng, Z.; Wang, H.; Jia, B.; Li, H.; Hu, Z. Identification of MicroRNAs Response to High Light and Salinity That Involved in Beta-Carotene Accumulation in Microalga Dunaliella salina. Algal Res. 2020, 48, 101925. [Google Scholar] [CrossRef]
- Takahashi-Íñiguez, T.; Aburto-Rodríguez, N.; Vilchis-González, A.L.; Flores, M.E. Function, Kinetic Properties, Crystallization, and Regulation of Microbial Malate Dehydrogenase. J. Zhejiang Univ. Sci. B 2016, 17, 247–261. [Google Scholar] [CrossRef]
- Goodwin, T.W. Biosynthesis of Carotenoids. In The Biochemistry of the Carotenoids; Springer: Dordrecht, The Netherlands, 1980; pp. 33–76. ISBN 978-94-009-5862-3. [Google Scholar]
- Xiao, R.; Zhou, H.; Chen, C.-M.; Cheng, H.; Li, H.; Xie, J.; Zhao, H.; Han, Q.; Diao, X. Transcriptional Responses of Acropora Hyacinthus Embryo under the Benzo(a)Pyrene Stress by Deep Sequencing. Chemosphere 2018, 206, 387–397. [Google Scholar] [CrossRef]
- Zhang, Q.-L.; Dong, Z.-X.; Xiong, Y.; Li, H.-W.; Guo, J.; Wang, F.; Deng, X.-Y.; Chen, J.-Y.; Lin, L.-B. Genome-Wide Transcriptional Response of MicroRNAs to the Benzo(a)Pyrene Stress in Amphioxus Branchiostoma belcheri. Chemosphere 2019, 218, 205–210. [Google Scholar] [CrossRef]
- Xiong, H.; Qian, J.; He, T.; Li, F. Independent Transcription of MiR-281 in the Intron of ODA in Drosophila melanogaster. Biochem. Biophys. Res. Commun. 2009, 378, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Raunio, H.; Vähäkangas, K.; Saarni, H.; Pelkonen, O. Effects of Cigarette Smoke on Rat Lung and Liver Ornithine Decarboxylase and Aryl Hydrocarbon Hydroxylase Activities and Lung Benzo(a)Pyrene Metabolism. Acta Pharmacol. Toxicol. 1983, 52, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Liu, X.; Zhang, H.; Mi, X. Identification of Micro-RNAs Responding to ZnSO4 Stress in Gonads. Fresenius Environ. Bull. 2015, 24, 4446–4451. [Google Scholar]
- Pan, Y.; Zhao, Z.; Zhou, Z. Identification of MiRNAs in Sea Urchin Strongylocentrotus purpuratus Larvae Response to PH Stress. Aquac. Res. 2021, 52, 4735–4744. [Google Scholar] [CrossRef]
- Wei, Z.; Range, R.; Angerer, R.; Angerer, L. Axial Patterning Interactions in the Sea Urchin Embryo: Suppression of Nodal by Wnt1 Signaling. Development 2012, 139, 1662–1669. [Google Scholar] [CrossRef]
- Wikramanayake, A.H.; Huang, L.; Klein, W.H. Beta-Catenin Is Essential for Patterning the Maternally Specified Animal-Vegetal Axis in the Sea Urchin Embryo. Proc. Natl. Acad. Sci. USA 1998, 95, 9343–9348. [Google Scholar] [CrossRef]
- Weis, V.M.; Smith, G.J.; Muscatine, L. A “CO2 Supply” Mechanism in Zooxanthellate Cnidarians: Role of Carbonic Anhydrase. Mar. Biol. 1989, 100, 195–202. [Google Scholar] [CrossRef]
- Mitsunaga, K.; Akasaka, K.; Shimada, H.; Fujino, Y.; Yasumasu, I.; Numanoi, H. Carbonic Anhydrase Activity in Developing Sea Urchin Embryos with Special Reference to Calcification of Spicules. Cell Differ. 1986, 18, 257–262. [Google Scholar] [CrossRef]
- Huang, C.-W.; Chen, Y.-W.; Lin, Y.-R.; Chen, P.-H.; Chou, M.-H.; Lee, L.-J.; Wang, P.-Y.; Wu, J.-T.; Tsao, Y.-P.; Chen, S.-L. Conditional Knockout of Breast Carcinoma Amplified Sequence 2 (BCAS2) in Mouse Forebrain Causes Dendritic Malformation via β-Catenin. Sci. Rep. 2016, 6, 34927. [Google Scholar] [CrossRef]
- Yu, D.; Peng, Z.; Wu, H.; Zhang, X.; Ji, C.; Peng, X. Stress Responses in Expressions of MicroRNAs in Mussel Mytilus galloprovincialis Exposed to Cadmium. Ecotoxicol. Environ. Saf. 2021, 212, 111927. [Google Scholar] [CrossRef]
- Chromik, J.; Safferthal, C.; Serve, H.; Fulda, S. Smac Mimetic Primes Apoptosis-Resistant Acute Myeloid Leukaemia Cells for Cytarabine-Induced Cell Death by Triggering Necroptosis. Cancer Lett. 2014, 344, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Sirenko, V.V.; Dobrzhanskaya, A.V.; Shelud’ko, N.S.; Borovikov, Y.S. Calponin-like Protein from Mussel Smooth Muscle Is a Competitive Inhibitor of Actomyosin ATPase. Biochemistry 2016, 81, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.-Y.; Xu, S.-M.; Wang, Y.-Y.; Long, X.-D.; Ma, S.-N.; Zhou, C.-X.; Xu, J.-L.; Yan, X.-J. Integrated Analyses of MiRNome and Transcriptome Reveal the Critical Role of MiRNAs Toward Heat Stress Response in Isochrysis galbana. Mar. Biotechnol. 2022, 24, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Finka, A.; Goloubinoff, P. How Do Plants Feel the Heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lv, X.; Zhang, J.; Chen, D.; Chen, S.; Fan, G.; Ma, C.; Wang, Y. Roles of E3 Ubiquitin Ligases in Plant Responses to Abiotic Stresses. Int. J. Mol. Sci. 2022, 23, 2308. [Google Scholar] [CrossRef]
- Diener, C.; Keller, A.; Meese, E. Emerging Concepts of MiRNA Therapeutics: From Cells to Clinic. Trends Genet. 2022, 38, 613–626. [Google Scholar] [CrossRef]
- Liang, L.; He, X. A Narrative Review of MicroRNA Therapeutics: Understanding the Future of MicroRNA Research. Precis. Cancer Med. 2021, 4, 33. [Google Scholar] [CrossRef]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for MicroRNA Therapeutics and Clinical Research. Front. Genet. 2019, 10, 478. [Google Scholar] [CrossRef]
- John, A.A.; Xie, J.; Yang, Y.-S.; Kim, J.-M.; Lin, C.; Ma, H.; Gao, G.; Shim, J.-H. AAV-Mediated Delivery of Osteoblast/Osteoclast-Regulating MiRNAs for Osteoporosis Therapy. Mol. Ther. Nucleic Acids 2022, 29, 296–311. [Google Scholar] [CrossRef]
- Carrella, S.; Di Guida, M.; Brillante, S.; Piccolo, D.; Ciampi, L.; Guadagnino, I.; Garcia Piqueras, J.; Pizzo, M.; Marrocco, E.; Molinari, M.; et al. miR-181a/b Downregulation: A Mutation-independent Therapeutic Approach for Inherited Retinal Diseases. EMBO Mol. Med. 2022, 14, e15941. [Google Scholar] [CrossRef]
- Wang, C.; Li, Y.; Yi, Y.; Liu, G.; Guo, R.; Wang, L.; Lan, T.; Wang, W.; Chen, X.; Chen, S.; et al. Hippocampal MicroRNA-26a-3p Deficit Contributes to Neuroinflammation and Behavioral Disorders via P38 MAPK Signaling Pathway in Rats. J. Neuroinflamm. 2022, 19, 283. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, X.; Li, H.; Wang, J.; Hu, Z. Artificial MiRNA Inhibition of Phosphoenolpyruvate Carboxylase Increases Fatty Acid Production in a Green Microalga Chlamydomonas reinhardtii. Biotechnol. Biofuels 2017, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Spillane, C. Reduction in Carotenoid Levels in the Marine Diatom Phaeodactylum tricornutum by Artificial MicroRNAs Targeted Against the Endogenous Phytoene synthase Gene. Mar. Biotechnol. 2015, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Kumar, S.; Verma, R.; Lata, C.; Sanyal, I.; Rai, S.P. MicroRNA 166: An Evolutionarily Conserved Stress Biomarker in Land Plants Targeting HD-ZIP Family. Physiol. Mol. Biol. Plants 2021, 27, 2471–2485. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, S.; Singh, M.P.V.V.B.; Singh, V.P.; Prasad, S.M. Micro RNAs and Nitric Oxide Cross Talk in Stress Tolerance in Plants. Plant Growth Regul. 2017, 83, 199–205. [Google Scholar] [CrossRef]
- Abo-Al-Ela, H.G.; Faggio, C. MicroRNA-Mediated Stress Response in Bivalve Species. Ecotoxicol. Environ. Saf. 2021, 208, 111442. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Falco, G.; Lauritano, C.; Carrella, S. MicroRNA-Mediated Responses: Adaptations to Marine Extreme Environments. J. Mar. Sci. Eng. 2023, 11, 361. https://doi.org/10.3390/jmse11020361
De Falco G, Lauritano C, Carrella S. MicroRNA-Mediated Responses: Adaptations to Marine Extreme Environments. Journal of Marine Science and Engineering. 2023; 11(2):361. https://doi.org/10.3390/jmse11020361
Chicago/Turabian StyleDe Falco, Gabriele, Chiara Lauritano, and Sabrina Carrella. 2023. "MicroRNA-Mediated Responses: Adaptations to Marine Extreme Environments" Journal of Marine Science and Engineering 11, no. 2: 361. https://doi.org/10.3390/jmse11020361