
Searching for the Origin and the Differentiation of Haemocytes before and after Larval Settlement of the Colonial Ascidian Botryllus schlosseri: An Ultrastructural Viewpoint


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Transmission Electron Microscopy
2.3. Histoenzymatic Assays
2.3.1. Assay for Acid Phosphatase
2.3.2. Assay for Phenoloxidase
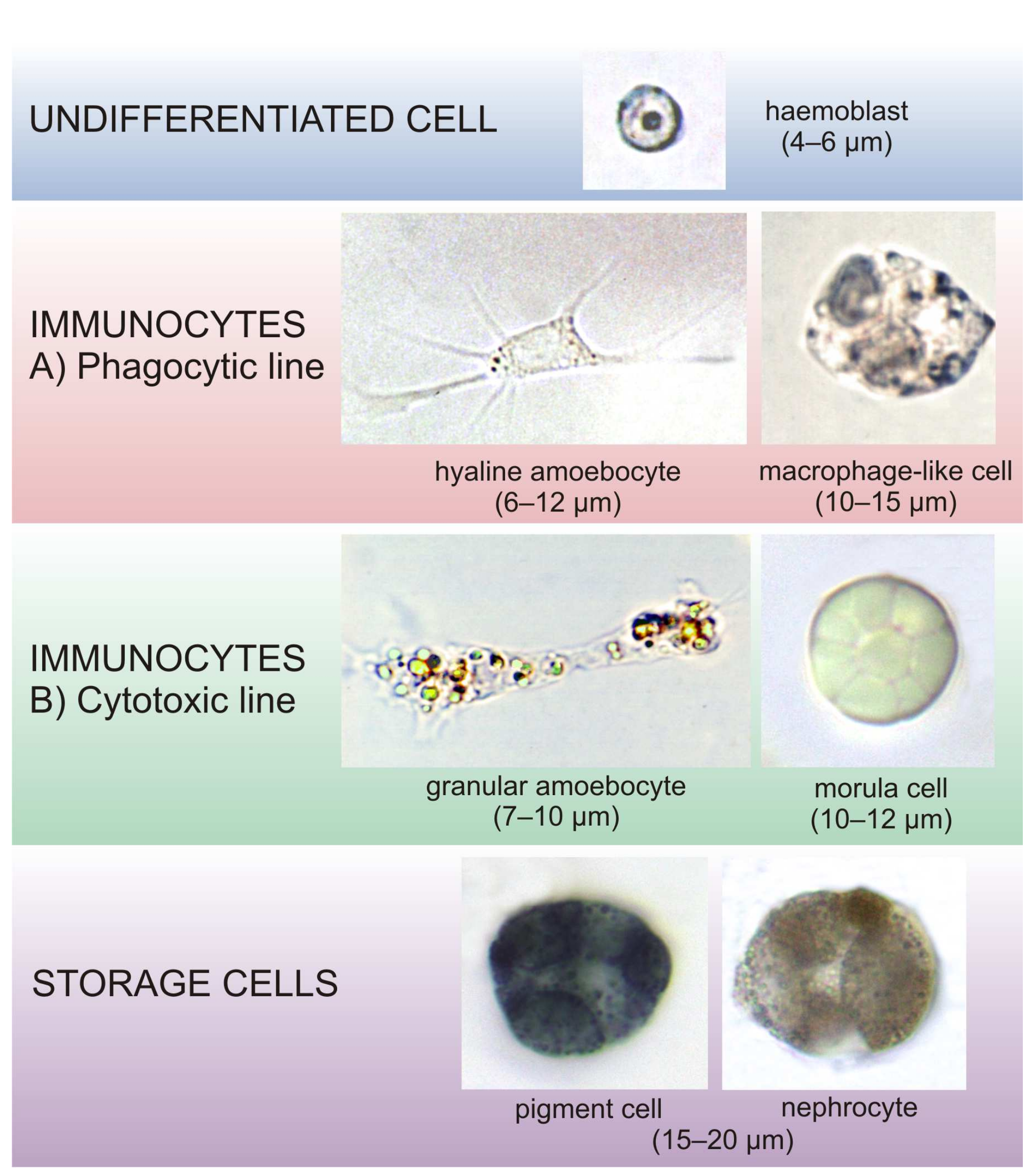
3. Results
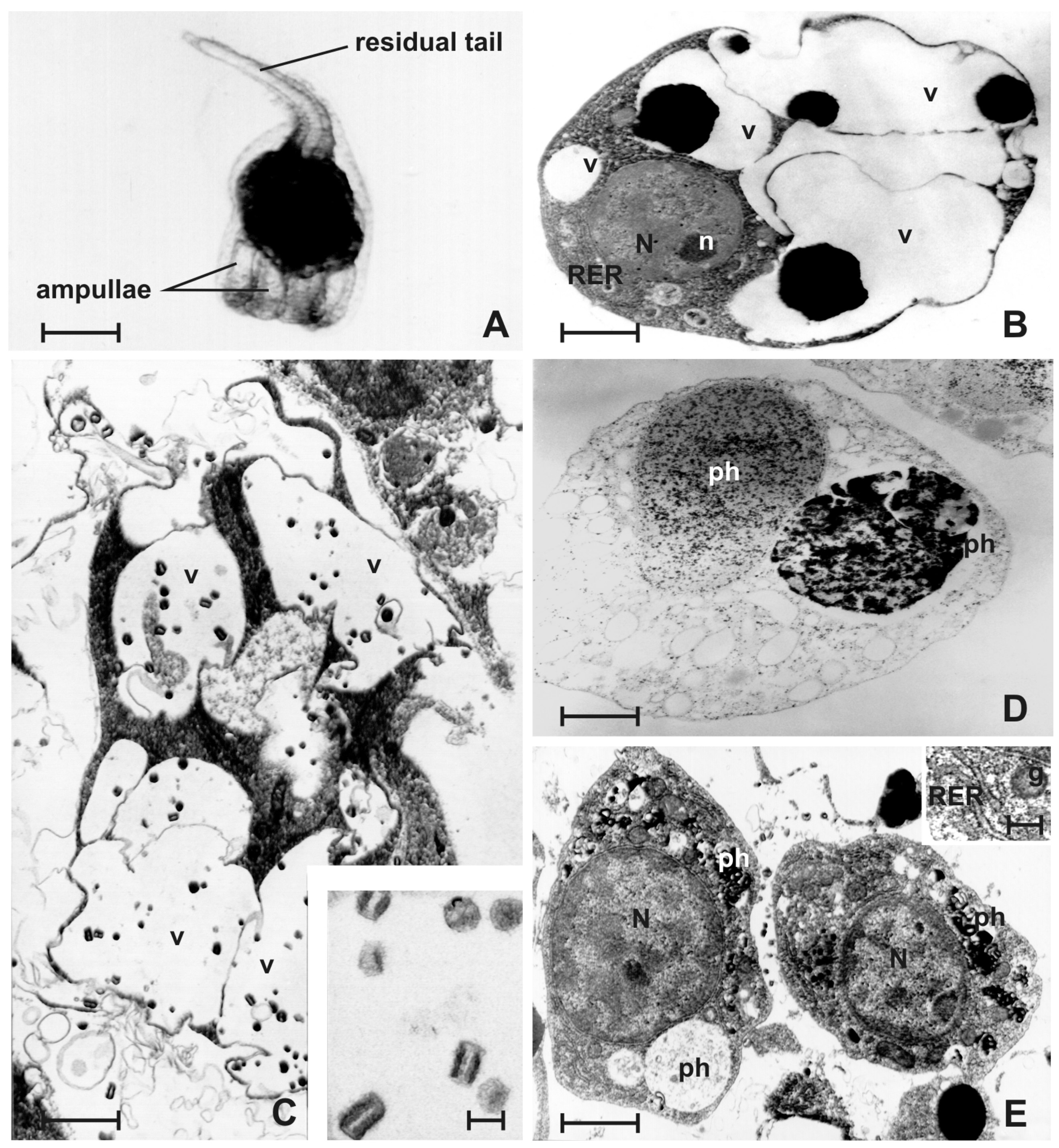
3.1. Early Tailbud Embryo
3.2. Late Tailbud Embryo
3.3. Newly Hatched Tadpole Larva (Early Metamorphosis)
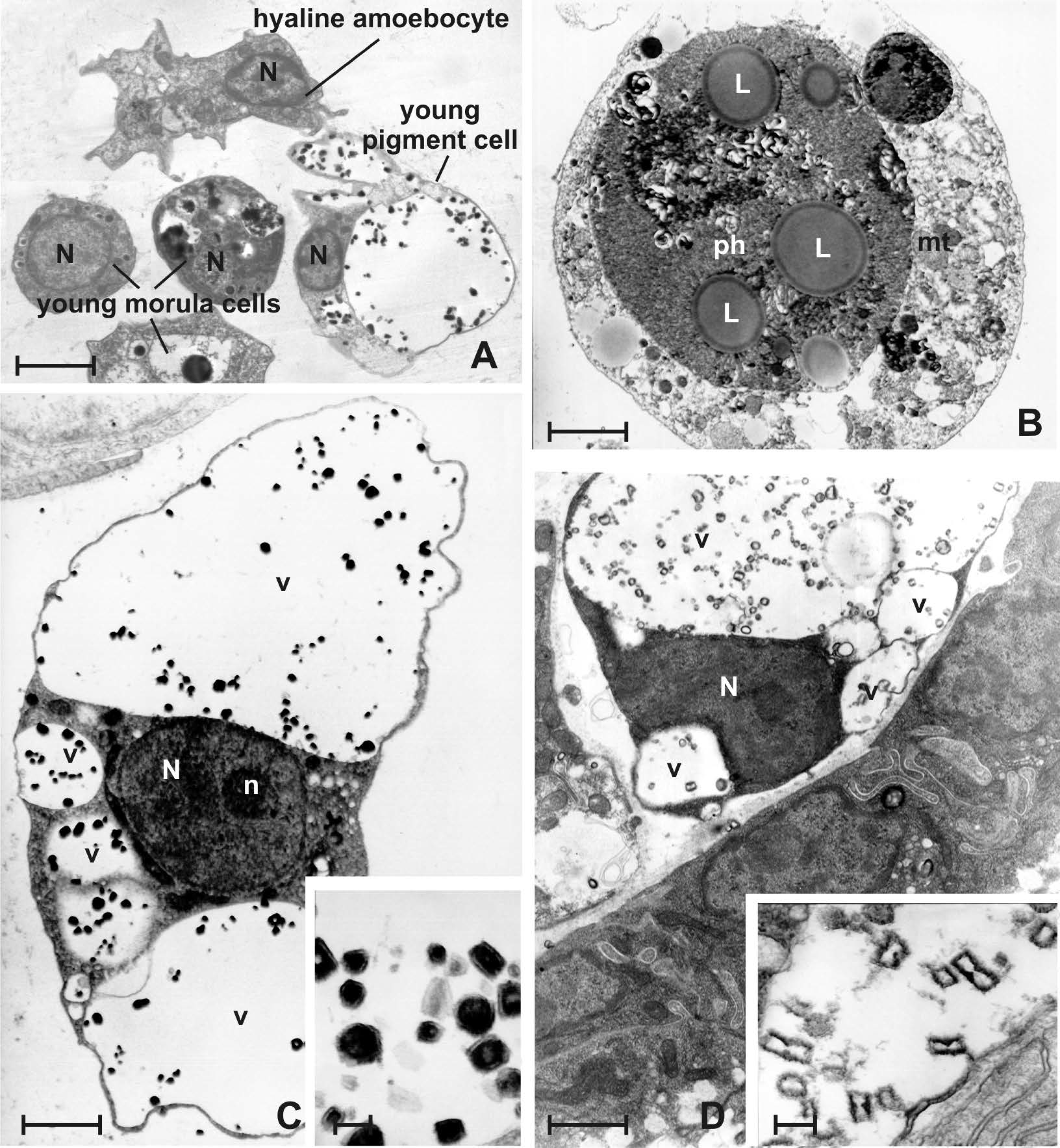
3.4. Late Larval Metamorphosis
3.5. Oozoid
4. Discussion
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Cima, F.; Ballarin, L.; Caicci, F.; Franchi, N.; Gasparini, F.; Rigon, F.; Schiavon, F.; Manni, L. Life history and ecological genetics of the colonial ascidian Botryllus schlosseri. Zool. Anz. 2015, 257, 54–70. [Google Scholar] [CrossRef]
- Reem, E.; Douek, J.; Rinkevich, B. A critical deliberation of the ‘species complex’ status of the globally spread colonial ascidian Botryllus schlosseri. J. Mar. Biol. Assoc. UK 2021, 101, 1047–1060. [Google Scholar] [CrossRef]
- Rosengarten, R.D.; Nicotra, M.L. Model systems of invertebrate allorecognition. Curr. Biol. 2011, 21, R82–R92. [Google Scholar] [CrossRef] [PubMed]
- Ballarin, L.; Cima, F.; Franchi, N. Origin and functions of tunicate hemocytes. In The Evolution of the Immune System: Conservation and Diversification, 1st ed.; Malagoli, D., Ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2016; Chapter 2; pp. 29–49. [Google Scholar]
- Delsuc, F.; Tsagkogeorga, G.; Lartillot, N.; Philippe, H. Additional molecular support for the new chordate phylogeny. Genesis 2008, 46, 592–604. [Google Scholar] [CrossRef]
- Putnam, N.; Butts, T.; Ferrier, D.; Furlong, R.F.; Hellsten, U.; Kawashima, T.; Robinson-Rechavi, M.; Shoguchi, E.; Terry, A.; Dubchak, I.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 2008, 453, 1064–1071. [Google Scholar] [CrossRef]
- Fujimoto, H.; Watanabe, H. The characterization of granular amebocytes and their possible roles in the asexual reproduction of the plystyelid ascidian, Polyzoa vesiculiphora. J. Morphol. 1976, 150, 623–638. [Google Scholar] [CrossRef]
- Goodbody, I. The physiology of ascidians. Adv. Mar. Biol. 1974, 12, 1–149. [Google Scholar]
- Oka, H.; Watanabe, H. Vascular budding, a new type of budding in Botryllus. Biol. Bull. 1957, 112, 225–240. [Google Scholar] [CrossRef]
- Freeman, G. The role of blood cells in the process of asexual reproduction in the tunicate Perophora viridis. J. Exp. Zool. 1964, 156, 157–183. [Google Scholar] [CrossRef]
- Sabbadin, A.; Zaniolo, G. Sexual differentiation and germ cell transfer in the colonial ascidian Botryllus schlosseri. J. Exp. Zool. 1979, 207, 289–304. [Google Scholar] [CrossRef]
- Kassmer, S.H.; Langenbacher, A.D.; De Tomaso, A.W. Integrin-alpha-6+ candidate stem cells are responsible for whole body regeneration in the invertebrate chordate Botrylloides diegensis. Nat. Commun. 2020, 11, 4435. [Google Scholar] [CrossRef] [PubMed]
- Manni, L.; Anselmi, C.; Cima, F.; Gasparini, F.; Voskoboynik, A.; Martini, M.; Peronato, A.; Burighel, P.; Zaniolo, G.; Ballarin, L. Sixty years of experimental studies on the blastogenesis of the colonial tunicate Botryllus schlosseri. Dev. Biol. 2019, 448, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Hirose, E.; Taneda, Y.; Ishii, T. Two modes of tunic cuticle formation in a colonial ascidian Aplidium yamazii, responding to wounding. Dev. Comp. Immunol. 1997, 21, 25–34. [Google Scholar] [CrossRef]
- Franchi, N.; Ballarin, L. Immunity in protochordates: The tunicate perspective. Front. Immunol. 2017, 8, 674. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.S. Cellular responses to foreign bodies in the tunicate Molgula manhattensis (DeKay). Biol. Bull. 1971, 141, 91–98. [Google Scholar] [CrossRef]
- Wright, R.K.; Cooper, E.L. Immunological maturation in the tunicate Ciona intestinalis. Am. Zool. 1975, 15, 21–27. [Google Scholar] [CrossRef]
- Wright, R.K.; Cooper, E.L. Inflammatory reactions of protochordata. Am. Zool. 1983, 23, 205–211. [Google Scholar] [CrossRef]
- Ballarin, L.; Cima, F.; Sabbadin, A. Phagocytosis in the colonial ascidian Botryllus schlosseri. Dev. Comp. Immunol. 1994, 18, 467–481. [Google Scholar] [CrossRef]
- Cima, F.; Ballarin, L.; Sabbadin, A. New data on phagocytes and phagocytosis in the compound ascidian Botryllus schlosseri (Tunicata: Ascidiacea). Ital. J. Zool. 1996, 63, 357–364. [Google Scholar] [CrossRef]
- Kelly, K.L.; Cooper, E.L.; Raftos, D.A. Purification and characterization of a humoral opsonin from the solitary urochordate Styela clava. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1992, 103, 749–753. [Google Scholar] [CrossRef]
- Raftos, D.A.; Cooper, E.L.; Habicht, G.S.; Beck, G. Invertebrate cytokines: Tunicate cell proliferation stimulated by an interleukin-1-like molecule. Proc. Natl. Acad. Sci. USA 1991, 88, 9518–9522. [Google Scholar] [CrossRef] [PubMed]
- Menin, A.; Ballarin, L. Immunomodulatory molecules in the compound ascidian Botryllus schlosseri: Evidence from conditioned media. J. Invertebr. Pathol. 2008, 99, 275–280. [Google Scholar] [CrossRef]
- Melillo, D.; Marino, R.; Della Camera, G.; Italiani, P.; Boraschi, D. Assessing immunological memory in the solitary ascidian Ciona robusta. Front. Immunol. 2019, 10, 1977. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, F.; Franchi, N.; Spolaore, B.; Ballarin, L. Novel rhamnose-binding lectins from the colonial ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2008, 32, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Franchi, N.; Schiavon, F.; Carletto, M.; Gasparini, F.; Bertoloni, G.; Tosatto, S.C.E.; Ballarin, L. Immune roles of a rhamnose-binding lectin in the colonial ascidian Botryllus schlosseri. Immunobiology 2011, 216, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Franchi, N.; Ballarin, L. Preliminary characterization of complement in a colonial tunicate: C3, Bf and inhibition of C3 opsonic activity by compstatin. Dev. Comp. Immunol. 2014, 46, 430–438. [Google Scholar] [CrossRef]
- Peronato, A.; Drago, L.; Rothbächer, U.; Macor, P.; Ballarin, L.; Franchi, N. Complement system and phagocytosis in a colonial protochordate. Dev. Comp. Immunol. 2020, 103, 103530. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Lee, I.H.; Menzel, L.; Waring, A.; Zhao, C. Clavanins and styelins, α-helical antimicrobial peptides from the hemocytes of Styela clava. In Phylogenetic Perspectives on the Vertebrate Immune System. Advances in Experimental Medicine and Biology; Beck, G., Sugumaran, M., Cooper, E.L., Eds.; Springer: Boston, MA, USA, 2001; Volume 484, pp. 71–76. [Google Scholar]
- Parrinello, N.; Cammarata, M.; Parrinello, D. The inflammatory response of Urochordata: The basic process of the ascidians’ innate immunity. In Advances in Comparative Immunology; Cooper, E.L., Ed.; Springer: Cham, Switzerland, 2018; pp. 521–590. [Google Scholar]
- Sabbadin, A.; Zaniolo, G.; Ballarin, L. Genetic and cytological aspects of histocompatibility in ascidians. Ital. J. Zool. 1992, 59, 167–173. [Google Scholar] [CrossRef]
- Cima, F.; Sabbadin, A.; Ballarin, L. Cellular aspects of allorecognition in the compound ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2004, 28, 881–889. [Google Scholar] [CrossRef]
- De Tomaso, A.W.; Nyholm, S.V.; Palmeri, K.J.; Ishizuka, K.J.; Ludington, W.B.; Mitchel, K.; Weissman, I.L. Isolation and characterization of a protochordate histocompatibility locus. Nature 2005, 438, 454–459. [Google Scholar] [CrossRef]
- Nyholm, S.V.; Passegue, E.; Ludington, W.B.; Voskoboynik, A.; Mitchel, K.; Weissman, I.L.; De Tomaso, A.W. fester, a candidate allorecognition receptor from a primitive chordate. Immunity 2006, 25, 163–173. [Google Scholar] [CrossRef] [PubMed]
- McKitrick, T.R.; Muscat, C.C.; Pierce, J.D.; Bhattacharya, D.; De Tomaso, A.W. Allorecognition in a basal chordate consists of independent activating and inhibitory pathways. Immunity 2011, 34, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Milanesi, C.; Burighel, P. Blood cell ultrastructure of the ascidian Botryllus schlosseri. I. Hemoblast, granulocytes, macrophage, morula cell and nephrocyte. Acta Zool. 1978, 59, 135–147. [Google Scholar] [CrossRef]
- Burighel, P.; Milanesi, C.; Sabbadin, A. Blood cell ultrastructure of the ascidian Botryllus schlosseri. II. Pigment cells. Acta Zool. 1983, 64, 15–23. [Google Scholar] [CrossRef]
- Ballarin, L.; Cima, F. Cytochemical properties of Botryllus schlosseri haemocytes: Indications for morpho-functional characterisation. Eur. J. Histochem. 2005, 49, 255–264. [Google Scholar] [PubMed]
- Wright, R.K. Urochordates. In Invertebrate Blood Cells; Ratcliffe, N.A., Rowley, A.F., Eds.; Academic Press: New York, NY, USA, 1981; Volume 2, pp. 565–626. [Google Scholar]
- De Leo, G. Ascidian hemocytes and their involvement in defense reactions. Boll. Zool. 1992, 59, 195–213. [Google Scholar] [CrossRef]
- Burighel, P.; Cloney, R.A. Urochordata: Ascidiacea. In Microscopic Anatomy of Invertebrates: Hemichordata, Chaetognatha and the Invertebrate Chordates; Harrison, F.W., Ruppert, E.E., Eds.; Wiley-Liss, Inc.: New York, NY, USA, 1997; Volume 15, pp. 221–347. [Google Scholar]
- Jiménez-Merino, J.; de Abreu, I.S.; Hiebert, L.S.; Allodi, S.; Tiozzo, S.; De Barros, C.M.; Brown, F.D. Putative stem cells in the hemolymph and in the intestinal submucosa of the solitary ascidian Styela plicata. Evo Devo 2019, 10, 31. [Google Scholar] [CrossRef]
- Hirano, T.; Nishida, H. Developmental fates of larval tissues after metamorphosis in ascidian Halocynthia roretzi. I. Origin of mesodermal tissues of the juvenile. Dev. Biol. 1997, 192, 199–210. [Google Scholar] [CrossRef]
- Cowden, R.R. The embryonic origin of blood cells in the tunicate Clavelina. Trans. Am. Microsc. Sci. 1968, 87, 521–524. [Google Scholar] [CrossRef]
- Andrew, W. Phase microscope studies of living blood cells of the tunicates under normal and experimental conditions with a description of a new type of motile cell appendage. Q. J. Microsc. Sci. 1961, 102, 89–105. [Google Scholar] [CrossRef]
- Davidson, B.; Swalla, B.J. A molecular analysis of ascidian metamorphosis reveals activation of an innate immune response. Development 2002, 129, 4739–4751. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.; Davidson, B.; MacMaster, G.; Lockhart, V.; Ma, E.; Wallace, S.S.; Swalla, B.J. A complement response may activate metamorphosis in the ascidian Boltenia villosa. Dev. Genes Evol. 2007, 217, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, S.; Todd, C. Inhibition and facilitation of bryozoan and ascidian settlement by natural multi-species biofilms: Effects of film age and the roles of active and passive larval attachment. Mar. Biol. 1997, 128, 463–473. [Google Scholar] [CrossRef]
- Chase, A.L.; Dijkstra, J.A.; Harris, L.G. The influence of substrate material on ascidian larval settlement. Mar. Pollut. Bull. 2016, 106, 35–42. [Google Scholar] [CrossRef]
- Freckelton, M.; Nedved, B.; Hadfield, M. Induction of invertebrate larval settlement; different bacteria, different mechanisms? Sci. Rep. 2017, 7, 42557. [Google Scholar] [CrossRef]
- Dobretsov, S.; Rittschof, D. Love at first taste: Induction of larval settlement by marine microbes. Int. J. Mol. Sci. 2020, 21, 731. [Google Scholar] [CrossRef]
- Ermak, T.H. An autoradiographic demonstration of blood cell renewal in Styela clava (Urochordata: Ascidiacea). Experientia 1975, 31, 837–838. [Google Scholar] [CrossRef]
- Ermak, T.H. The hematogenic tissues of tunicates. In Phylogeny of Thymus and Bone Marrow-Bursa Cells; Wright, R.K., Cooper, E.L., Eds.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1976; pp. 45–56. [Google Scholar]
- Wright, R.K.; Ermak, T.H. Cellular defense systems of the protochordata. In Phylogeny and Ontogeny; Cohen, N., Sigel, M.M., Eds.; Springer: Boston, MA, USA, 1982; pp. 283–320. [Google Scholar]
- Zaniolo, G.; Burighel, P.; Martinucci, G.B. Ovulation and placentation in Botryllus schlosseri (Ascidiacea): An ultrastructural study. Can. J. Zool. 1987, 65, 1181–1190. [Google Scholar] [CrossRef]
- Manni, L.; Lane, N.J.; Sorrentino, M.; Zaniolo, G.; Burighel, P. Mechanism of neurogenesis during the embryonic development of a tunicate. J. Comp. Neurol. 1999, 412, 527–541. [Google Scholar] [CrossRef]
- Manni, L.; Burighel, P. Common and divergent pathways in alternative developmental processes of ascidians. Bioessays 2006, 28, 902–912. [Google Scholar] [CrossRef]
- Cima, F.; Varello, R. Effects of exposure to trade antifouling paints and biocides on larval settlement and metamorphosis of the compound ascidian Botryllus schlosseri. J. Mar. Sci. Eng. 2022, 10, 123. [Google Scholar] [CrossRef]
- Müller, W.C.; Greenwood, A.D. The ultrastructure of phenolic-storing cells fixed with caffeine. J. Exp. Bot. 1978, 29, 757–764. [Google Scholar] [CrossRef]
- Cima, F. Enzyme histochemistry for functional histology in invertebrates. In Single Molecule Histochemistry: Methods and Protocols; Pellicciari, C., Biggiogera, M., Eds.; Methods in Molecular Biology; Springer Science: New York, NY, USA, 2017; Volume 1560, pp. 69–90. [Google Scholar]
- Barka, T.; Anderson, P.J. Histochemical methods for acid phosphatase using hexazonium pararosanilin as coupler. J. Histochem. Cytochem. 1962, 10, 741–753. [Google Scholar] [CrossRef]
- Hose, J.E.; Martin, G.G.; Nguyen, V.A.; Lucas, J.; Rosenstein, A.T. Cytochemical features of shrimp hemocytes. Biol. Bull. 1987, 173, 178–187. [Google Scholar] [CrossRef]
- Kowarsky, M.; Anselmi, C.; Hotta, K.; Burighel, P.; Zaniolo, G.; Caicci, F.; Rosental, B.; Neff, N.F.; Ishizuka, K.J.; Palmeri, K.J.; et al. Sexual and asexual development: Two distinct programs producing the same tunicate. Cell Rep. 2021, 34, 108681. [Google Scholar] [CrossRef]
- Burighel, P.; Brunetti, R. The circulatory system in the blastozooid of the colonial ascidian Botryllus schlosseri (Pallas). Boll. Zool. 1971, 38, 273–289. [Google Scholar] [CrossRef]
- Katow, H.; Watanabe, H. Fine structure and possible role of ampullae on tunic supply and attachment in a compound ascidian, Botryllus primigenus Oka. J. Ultrastruct. Res. 1978, 64, 23–34. [Google Scholar] [CrossRef]
- Rodriguez, D.; Nourizadeh, S.; De Tomaso, A.W. The biology of the extracorporeal vasculature of Botryllus schlosseri. Dev. Biol. 2019, 448, 309–319. [Google Scholar] [CrossRef]
- Schiaffino, S.; Burighel, P.; Nunzi, M.G. Involution of the caudal musculature during metamorphosis in the ascidian, Botryllus schlosseri. Cell Tissue Res. 1974, 153, 293–305. [Google Scholar] [CrossRef]
- Manni, L.; Zaniolo, G.; Burighel, P. 1993 Egg envelope cytodifferentiation in the colonial ascidian Botryllus schlosseri (Tunicata). Acta Zool. 1993, 74, 103–113. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Voskoboynik, A.; Rosner, A.; Rabinowitz, C.; Paz, G.; Oren, M.; Douek, J.; Alfassi, G.; Moiseeva, E.; Ishizuka, K.J.; et al. Repeated, long-term cycling of putative stem cells between niches in a basal chordate. Dev. Cell. 2013, 24, 76–88. [Google Scholar] [CrossRef]
- Rosental, B.; Kowarsky, M.; Seita, J.; Corey, D.M.; Ishizuka, J.K.; Palmeri, K.J.; Chen, S.Y.; Sinha, R.; Okamoto, J.; Mantalas, G.; et al. Complex mammalian-like haematopoietic system found in a colonial chordate. Nature 2018, 564, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Ballarin, L.; Karahan, A.; Salvetti, A.; Rossi, L.; Manni, L.; Rinkevich, B.; Rosner, A.; Voskoboynik, A.; Rosental, B.; Canesi, L.; et al. Stem cells and innate immunity in aquatic invertebrates: Bridging two seemingly disparate disciplines for new discoveries in biology. Front. Immunol. 2021, 12, 688106. [Google Scholar] [CrossRef] [PubMed]
- Fontane, A.R.; Lambert, P. The fine structure of the leucocytes of the holothurian Cucumaria miniata. Can. J. Zool. 1977, 55, 1530–1544. [Google Scholar] [CrossRef] [PubMed]
- Smith, U.Z. The echinoderms. In Invertebrate Blood Cells; Ratcliffe, N.A., Rowley, A.F., Eds.; Academic Press: New York, NY, USA, 1981; Volume 2, pp. 513–562. [Google Scholar]
- Azéma, M. Le sang des Botrylles. Comptes Rendus Séances Soc. Biol. Paris 1929, 192, 823–825. [Google Scholar]
- Azéma, M. Recherches sur le sang et l’excrétion chez les ascidies. Ann. Inst. Océan. Monaco 1937, 17, 1–150. [Google Scholar]
- Endean, R. The blood cells of the ascidian Phallusia mammillata. Q. J. Microsc. Sci. 1960, 101, 177–197. [Google Scholar]
- George, W.C. A comparative study of the blood of the tunicates. Q. J. Microsc. Sci. 1939, 81, 391–428. [Google Scholar]
- Mackie, G.O.; Singla, C.L. Coordination of compound ascidians by epithelial conduction in the colonial blood vessels. Biol. Bull. 1983, 165, 209–220. [Google Scholar] [CrossRef]
- Voskoboynik, A.; Soen, Y.; Rinkevich, Y.; Rosner, A.; Ueno, H.; Reshef, R.; Ishizuka, K.J.; Palmeri, K.J.; Moiseeva, E.; Rinkevich, B.; et al. Identification of the endostyle as a stem cell niche in a colonial chordate. Cell Stem Cell 2008, 3, 456–464. [Google Scholar] [CrossRef]
- Matozzo, V.; Marin, M.G.; Cima, F.; Ballarin, L. First evidence of cell division in circulating haemocytes from the Manila clam Tapes philippinarum. Cell Biol. Int. 2008, 32, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Cima, F.; Matozzo, V. Proliferation and differentiation of circulating haemocytes of Ruditapes philippinarum as a response to bacterial challenge. Fish Shellfish Immunol. 2018, 81, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Cerenius, L.; Söderhäll, K. Immune properties of invertebrate phenoloxidases. Dev. Comp. Immunol. 2021, 122, 104098. [Google Scholar] [CrossRef] [PubMed]
- Chaga, O.Y. Ortho-diphenoloxidase system of ascidians. Tsitologia 1980, 22, 619–625. [Google Scholar]
- Smith, V.J.; Söderhäll, K. A comparison of phenoloxidase activity in the blood of marine invertebrates. Dev. Comp. Immunol. 1991, 15, 251–262. [Google Scholar] [CrossRef]
- Ballarin, L.; Cima, F.; Sabbadin, A. Phenoloxidase and cytotoxicity in the compound ascidian Botryllus schlosseri. Dev. Comp. Immunol. 1998, 22, 479–492. [Google Scholar] [CrossRef]
- Ballarin, L.; Franchini, A.; Ottaviani, E.; Sabbadin, A. Morula cells as the major immunomodulatory hemocytes in ascidians: Evidences from the colonial species Botryllus schlosseri. Biol. Bull. 2001, 201, 59–64. [Google Scholar] [CrossRef]
- Ballarin, L.; Cima, F.; Sabbadin, A. Morula cells and histocompatibility in the colonial ascidian Botryllus schlosseri. Zool. Sci. 2005, 12, 757–764. [Google Scholar] [CrossRef]
- Ballarin, L.; Franchi, N.; Schiavon, F.; Tosatto, S.C.E.; Mičetić, I.; Kawamura, K. Looking for putative phenoloxidases of compound ascidians: Haemocyanin-like proteins in Polyandrocarpa misakiensis and Botryllus schlosseri. Dev. Comp. Immunol. 2012, 38, 232–242. [Google Scholar] [CrossRef]
- Taketa, D.A.; De Tomaso, A.W. Botryllus schlosseri allorecognition: Tackling the enigma. Dev. Comp. Immunol. 2014, 48, 254–265. [Google Scholar] [CrossRef]
- Izzard, C.S. Contractile filopodia and in vivo cell movement in the tunic of the ascidian, Botryllus schlosseri. J. Cell Sci. 1974, 15, 513–535. [Google Scholar] [CrossRef]
- Zaniolo, G. Histology of the ascidian Botryllus schlosseri: In particular, the test cells. Boll. Zool. 1981, 48, 169–178. [Google Scholar] [CrossRef]
- Hirose, E. Ascidian tunic cells: Morphology and functional diversity of free cells outside the epidermis. Invertebr. Biol. 2009, 128, 83–96. [Google Scholar] [CrossRef]
- Hirose, E.; Mukai, H. An ultrastructural study on the origin of glomerulocytes, a type of blood cells in a styelid ascidian, Polyandrocarpa misakiensis. J. Morphol. 1992, 211, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Itoh, T. Evidence for the role of glomerulocyte in cellulose synthesis in the tunicate, Metandrocarpa uedai. Protoplasma 1995, 186, 24–33. [Google Scholar] [CrossRef]
- Di Bella, M.A.; Carbone, M.C.; De Leo, G. Aspects of cell proliferation in mantle tissue of Ciona intestinalis L. (Tunicata, Ascidiacea). Micron 2005, 36, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Dias, G.M.; Yokoyama, L.Q. Spatial competition induces the mobilization of morula cells in the colonial ascidian Didemnum perlucidum (Tunicata: Didemnidae). Invertebr. Biol. 2011, 130, 186–192. [Google Scholar] [CrossRef]
- Ballarin, L. Ascidian cytotoxic cells: State of the art and research perspectives. Invertebr. Surv. J. 2012, 9, 1–6. [Google Scholar]
- Melillo, D.; Marino, R.; Italiani, P.; Boraschi, D. Innate immune memory in invertebrate metazoans: A critical appraisal. Front. Immunol. 2018, 9, 1915. [Google Scholar] [CrossRef]
- Franchi, N.; Ballarin, L.; Cima, F. Insights on cytotoxic cells of the colonial ascidian Botryllus schlosseri. Invertebr. Surv. J. 2015, 12, 109–117. [Google Scholar]
- Franchi, N.; Ballarin, L.; Peronato, A.; Cima, F.; Grimaldi, A.; Girardello, R.; de Eguileor, M. Functional amyloidogenesis in immunocytes from the colonial ascidian Botryllus schlosseri: Evolutionary perspective. Dev. Comp. Immunol. 2019, 90, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Taneda, Y.; Watanabe, H. Studies on colony specificity in the compound ascidian, Botryllus primigenus Oka. 1. Blood cells infiltration. Dev. Comp. Immunol. 1982, 6, 43–52. [Google Scholar] [CrossRef]
- Hirose, E.; Saito, Y.; Watanabe, H. Subcuticular rejection: An advanced mode of the allogeneic rejection in the compound ascidians Botrylloides simodensis and B. fuscus. Biol. Bull. 1997, 192, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Ballarin, L.; Cima, F.; Floreani, M.; Sabbadin, A. Oxidative stress induces cytotoxicity during rejection reaction in the compound ascidian Botryllus schlosseri. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 133, 411–418. [Google Scholar] [CrossRef]
- Cima, F.; Sabbadin, A.; Zaniolo, G.; Ballarin, L. Colony specificity and chemotaxis in the compound ascidian Botryllus schlosseri. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 145, 376–382. [Google Scholar] [CrossRef]
- Gasparini, F.; Longo, F.; Manni, L.; Burighel, P.; Zaniolo, G. Tubular sprouting as a mode of vascular formation in a colonial ascidian (Tunicata). Dev. Dyn. 2007, 236, 719–731. [Google Scholar] [CrossRef]
- Grimm, L.M.; Cloney, R.A. Ultrastructural study of ascidian metamorphosis: Blood cell migration across the epidermis. Am. Zool. 1965, 5, 644. [Google Scholar]
- Cloney, R.A. Ascidian larvae and the events of metamorphosis. Am. Zool. 1982, 22, 817–826. [Google Scholar] [CrossRef]
- Burighel, P.; Schiavinato, A. Degenerative regression of the digestive tract in the colonial ascidian Botryllus schlosseri (Pallas). Cell Tissue Res. 1984, 235, 309–318. [Google Scholar] [CrossRef]
- Cima, F.; Ballarin, L. Apoptosis and pattern of Bcl-2 and Bax expression in the alimentary tract during the colonial blastogenetic cycle of Botryllus schlosseri (Urochordata, Ascidiacea). Ital. J. Zool. 2009, 76, 28–42. [Google Scholar] [CrossRef]
- Cima, F.; Basso, G.; Ballarin, L. Apoptosis and phosphatidylserine-mediated recognition during the take-over of the colonial life-cycle in the ascidian Botryllus schlosseri. Cell Tissue Res. 2003, 312, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Cima, F.; Manni, L.; Basso, G.; Fortunato, E.; Accordi, B.; Schiavon, F.; Ballarin, L. Hovering between death and life: Natural apoptosis and phagocytes in the blastogenetic cycle of the colonial ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2010, 34, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Ballarin, L.; Cima, F.; Sabbadin, A. Histoenzymatic staining and characterization of the colonial ascidian Botryllus schlosseri hemocytes. Boll. Zool. 1993, 60, 19–24. [Google Scholar] [CrossRef]
- Sabbadin, A. Formal genetics of ascidians. Am. Zool. 1982, 22, 765–773. [Google Scholar] [CrossRef]
- Mukai, H. Photo-induced accumulation of pigment cells in a compound ascidian, Botryllus primigenus. Ann. Zool. Jpn. 1974, 47, 43–47. [Google Scholar]
- Sabbadin, A.; Tontodonati, A. Nitrogenous excretion in the compound ascidians Botryllus schlosseri (Pallas) and Botrylloides leachi (Savigny). Monit. Zool. Ital. 1967, 1, 185–190. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Early Tailbud Embryo | Late Tailbud Embryo | Early Metamorphosis | Late Metamorphosis | Oozoid | |
|---|---|---|---|---|---|
| Haemoblasts | |||||
| Morula cells | PO | PO | PO | ||
| Hyaline amoebocytes | AP | AP | |||
| Pigment cells | |||||
| Macrophage-like cells | AP | AP | |||
| Nephrocytes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cima, F. Searching for the Origin and the Differentiation of Haemocytes before and after Larval Settlement of the Colonial Ascidian Botryllus schlosseri: An Ultrastructural Viewpoint. J. Mar. Sci. Eng. 2022, 10, 987. https://doi.org/10.3390/jmse10070987
Cima F. Searching for the Origin and the Differentiation of Haemocytes before and after Larval Settlement of the Colonial Ascidian Botryllus schlosseri: An Ultrastructural Viewpoint. Journal of Marine Science and Engineering. 2022; 10(7):987. https://doi.org/10.3390/jmse10070987
Chicago/Turabian StyleCima, Francesca. 2022. "Searching for the Origin and the Differentiation of Haemocytes before and after Larval Settlement of the Colonial Ascidian Botryllus schlosseri: An Ultrastructural Viewpoint" Journal of Marine Science and Engineering 10, no. 7: 987. https://doi.org/10.3390/jmse10070987