
Movements of Hatchery-Reared Dusky Groupers Released in a Northeast Atlantic Coastal Marine Protected Area

 ,
,  , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Tagging and Tracking
2.3. Data Analysis
- The site attachment in the no-take areas, i.e., the selection of an area to settle [50], was assessed by analyzing the presence of each tagged fish in the study areas and how persistent was that presence.
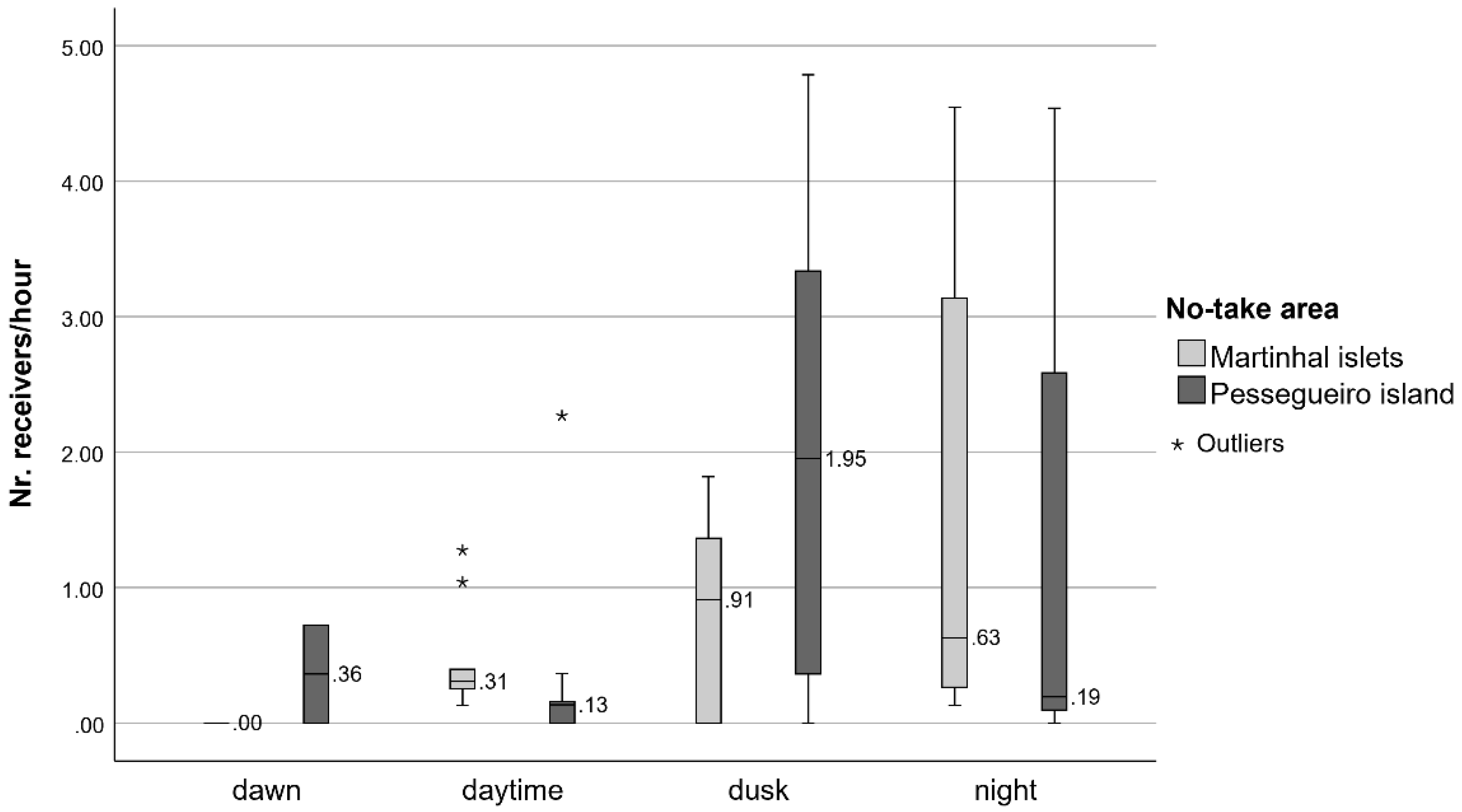
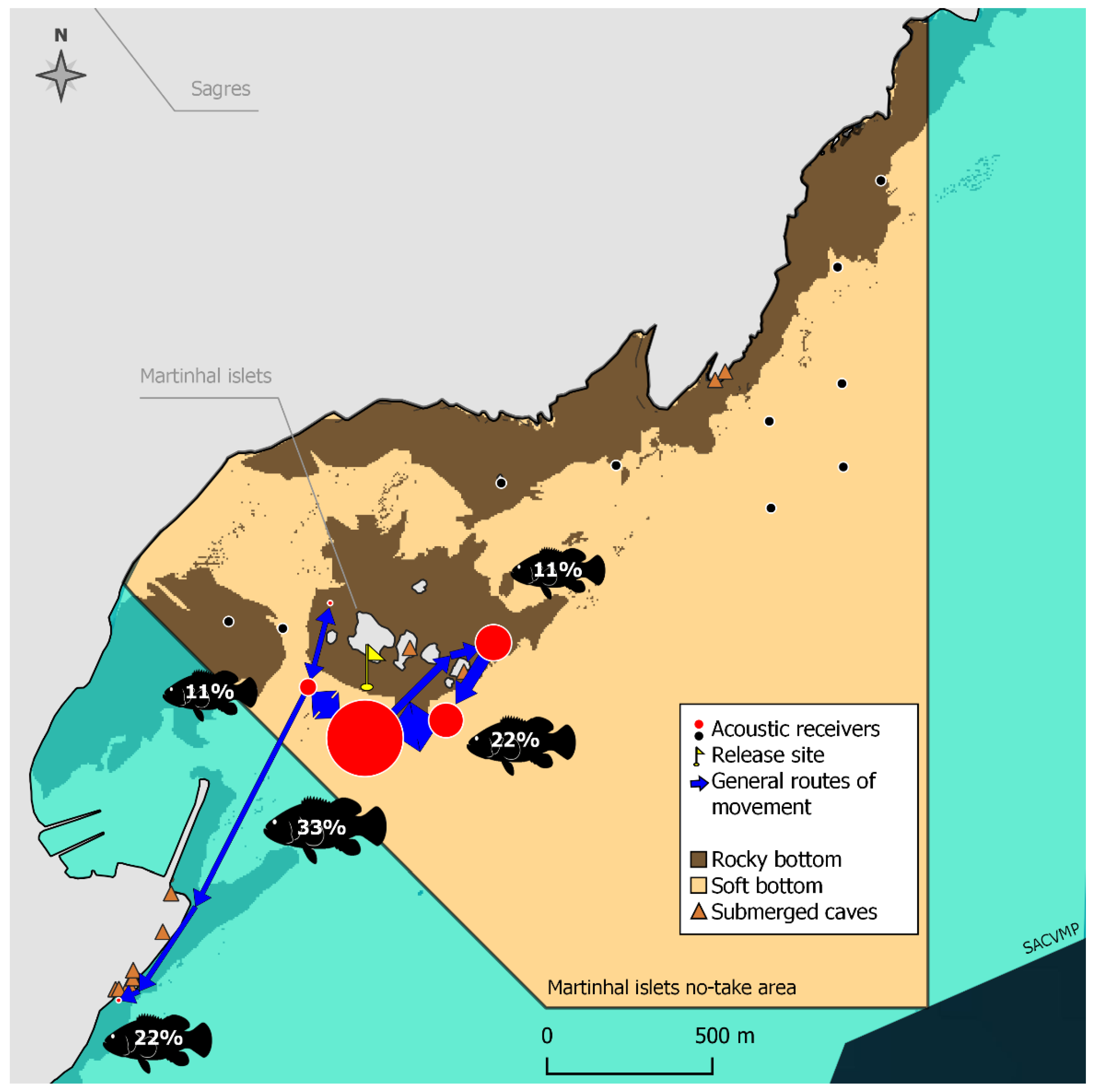
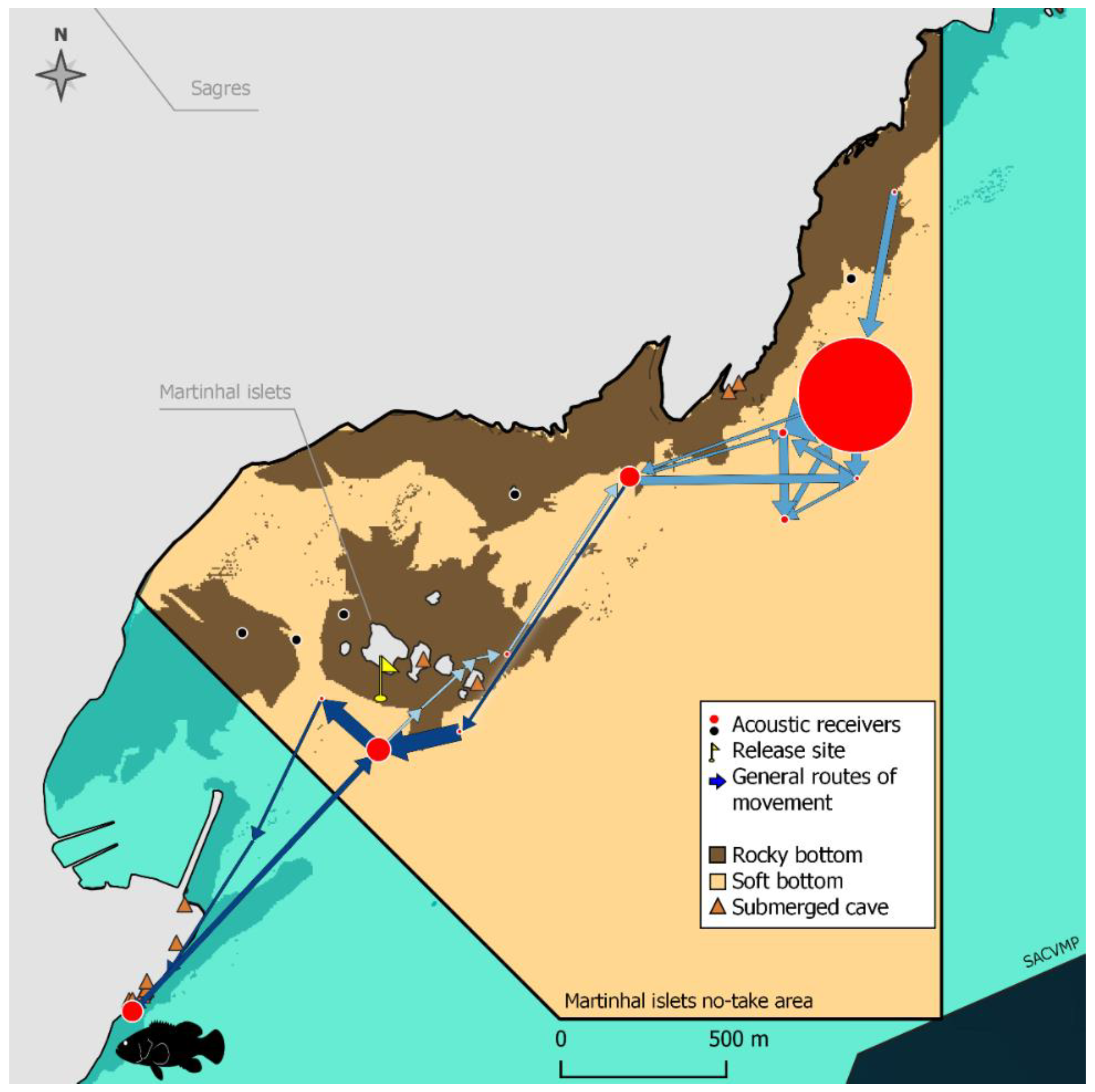
- The activity patterns and the movements of the dusky groupers inside the study areas (short-distance ranging movements) were also analyzed. Each fish was considered to be present in the study area while the interval between consecutive detections was less than 24 h. When the detections ceased permanently after a period of continuous presence, or whenever two consecutive detections were 24 h (or more) apart, it was assumed that the fish left the study area. Thus, for this analysis, only the set of detections between the releasing and the departure of the fish from the study area was considered.The activity and the movement patterns of the dusky groupers were analyzed in terms of the daily variation of the proportion of time in continuous activity and the number of different receivers with detections. The latter was evaluated as a proxy for the type of activity: stationary activity (detections in the same receiver) vs. movements (detections in multiple receivers) and as a proxy for the amplitude of those movements. The more receivers with detections, the wider the movements. Groupers were considered to be continuously active when intervals between consecutive detections were less than 10 min. Above this interval, individuals were assumed to be rarely and shortly active and hidden in refuges [44]. For each day in the study area, an activity index (IA) was calculated for each period of the day—Dawn, Daytime, Dusk, and Night (classified according to the national entity of Astronomy data [51]). The IA was calculated as the proportion of time of each period of the day during which each grouper was continuously active:where are the multiple (n) intervals in continuous activity and is the total duration of each period of the day. The IA values were then statistically compared to detect the daily variation of the fish activity. The number of receivers with detections at the four different periods of the day was weighted by the total duration of each period (NR) and the daily variation of this metric was also tested. All tests were performed through univariate Permutational analysis of variance (PERMANOVA) with Primer 6 and PERMANOVA+ [52], considering the ‘day period’ as a fixed factor with four levels—Dawn, Daytime, Dusk, and Night. Boxplots were made in IBM SPSS Statistics 27.The general routes used by the dusky groupers within the no-take areas and when exiting them were also tracked. These were computed through Refined Shortest Paths (RSP), an R package of tools that estimates the shortest distances traveled by monitored animals between pairs of detections using a least-cost path analysis by accounting for the presence of topographic features and constraints [53]. For consecutive detections in different acoustic receivers at least 250 m apart and with a10 min interval (minimum), intermediate positions of the tagged individuals were interpolated by RSP. Compared to the single use of the raw tracking data, more realistic movements of tracked animals were able to be estimated with this analysis tool. Each set of detections before a 24-h interval with no detections was treated as a distinct track, and distances traveled per track were computed in the R environment (RStudio v1.4.1717, RStudio, PBC) [53]. According to these calculations, the minimum distances traveled by each dusky grouper during the short-distance ranging movements in the study areas were estimated. The general routes of the movements were also visually evaluated by mapping the detections based on a Network Analysis, which evaluates the type and degree of interactions (edges) between activity centers (nodes) [54]. In this case, a spatial network, nodes were considered to represent acoustic receivers connected by edges, i.e., fish movements [44,55]. In this visual analysis, nodes were dimensioned according to the number of detections logged in the respective receivers and edge thickness according to the number of movements between two receivers [44,55]. The balance between the number of movements in each direction was represented by arrows. The proportion of fish detected for the last time was calculated and represented at the receivers where those detections occurred. These maps were prepared in QGis v3.16.8-Hannover, QGIS Development Team.Due to the distinct behavior of the grouper G29, i.e., a higher residency period inside the Martinhal islets no-take area, data analysis on this grouper was performed separately and was restricted to a visual graphical analysis (R packages ‘circular’ v0.4_93 and ‘circlize’ v0.4.13, RStudio v1.4.1717, RStudio, PBC) and mapping (QGis 3.16.8-Hannover, QGIS Development Team). Hourly circular graphs were built to visually analyze the daily variation of the total activity time and the number of receivers with detections, which were the same metrics statistically evaluated for the rest of the groupers. As for the other groupers, the general routes of the short-distance ranging movements of this fish were mapped.
- The ranging movements outside the study areas (long-distance ranging movements) were evaluated according to multiple reported events, such as detections in other biotelemetry acoustic receivers’ arrays deployed on the Portuguese coast in the framework of other projects and CoastNet infrastructure [56], recaptures by professional and recreational fishers, and researcher underwater sightings. Based on the date-timestamp of each reported event, and considering continuous and unidirectional movements at constant depths between 15 to 25 m, minimum distances and maximum time traveled were assigned, and ground speeds were estimated whenever possible.
3. Results
3.1. Site Attachment in the No-Take Areas
3.2. Activity Patterns and Short-Distance Ranging Movements (Inside the Study Areas)
3.3. Long-Distance Ranging Movements (Outside the Study Areas)
3.4. Grouper 29
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelleher, G.; Kenchington, R. Guidelines for Establishing Marine Protected Areas: A marine Conservation and Development Report; IUCN: Gland, Switzerland, 1992; pp. i–vii + 1–79; Available online: https://portals.iucn.org/library/sites/library/files/documents/MRN-001.pdf (accessed on 6 April 2022).
- Lausche, B.J. Guidelines for Protected Areas Legislation; 81; IUCN: Gland, Switzerland, 2011; pp. i–xxvi + 1–370; Available online: https://portals.iucn.org/library/efiles/documents/eplp-081.pdf (accessed on 6 April 2022).
- Day, J.C. Zoning—lessons from the Great Barrier Reef Marine Park. Ocean Coast. Manag. 2002, 45, 139–156. [Google Scholar] [CrossRef] [Green Version]
- Kramer, D.L.; Chapman, M.R. Implications of fish home range size and relocation for marine reserve function. Environ. Biol. Fishes 1999, 55, 65–79. [Google Scholar] [CrossRef]
- Di Franco, A.; Plass-Johnson, J.G.; Di Lorenzo, M.; Meola, B.; Claudet, J.; Gaines, S.D.; García-Charton, J.A.; Giakoumia, S.; Grorud-Colvert, K.; Hackradt, C.W.; et al. Linking home ranges to protected area size: The case study of the Mediterranean Sea. Biol. Conserv. 2018, 221, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Kellner, J.B.; Nisbet, R.M.; Gaines, S.D. Spillover from marine reserves related to mechanisms of population regulation. Theor. Ecol. 2008, 1, 117–127. [Google Scholar] [CrossRef]
- Halpern, B.S.; Warner, R.R. Matching marine reserve design to reserve objectives. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 1871–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, D.A.; Afonso, P.; Bertoncini, A.A.; Fennessy, S.; Francour, P.; Barreiros, J. Epinephelus marginatus. The IUCN Red List of Threatened Species 2018: E.T7859A100467602; IUCN: Gland, Switzerland, 2018. [Google Scholar]
- Harmelin-Vivien, M.; Craig, M.T. Epinephelus marginatus. The IUCN Red List of Threatened Species 2015: E.T7859A44904558; IUCN: Gland, Switzerland, 2015. [Google Scholar]
- Cornish, A.; Harmelin-Vivien, M. Epinephelus marginatus. The IUCN Red List of Threatened Species 2011: E.T7859A12856576; IUCN: Gland, Switzerland, 2011. [Google Scholar]
- Nieto, A.; Ralph, G.M.; Comeros-Raynal, M.T.; Kemp, J.; Criado, M.G.; Allen, D.J.; Dulvy, N.K.; Walls, R.H.L.; Russell, B.; Pollard, D.; et al. European Red List of Marine Fishes.; IUCN: Luxembourg, 2015; pp. i–iv + 1–81. [Google Scholar] [CrossRef]
- Assis, J.; Gonçalves, J.M.S.; Veiga, P.; Pita, P. Spearfishing in Portugal: A baseline study on spearfishers’ profiles, habits and perceptions towards management measures. Fish Manag. Ecol. 2018, 25, 417–428. [Google Scholar] [CrossRef]
- Condini, M.V.; García-Charton, J.A.; Garcia, A.M. A review of the biology, ecology, behavior and conservation status of the dusky grouper, Epinephelus marginatus (Lowe 1834). Rev. Fish Biol. Fish. 2018, 28, 301–330. [Google Scholar] [CrossRef]
- Marino, G.; Azzurro, E.; Massari, A.; Finoia, M.; Mandich, A. Reproduction in the dusky grouper from the southern Mediterranean. J. Fish Biol. 2001, 58, 909–927. [Google Scholar] [CrossRef]
- Condini, M.V.; Seyboth, E.; Vieira, J.P.; Barreiros, J.P.; Favaro, L.F.; Garcia, A.M. First record of the dusky grouper Epinephelus marginatus (Actinopterygii: Epinephelidae) undergoing sexual transition in the South Western Atlantic, Brazil. Hidrobiológica 2013, 23, 446–449. [Google Scholar]
- Zabala, M.; Louisy, P.; García-Rubies, A.; Gràcia, V. Socio-behavioural context of reproduction in the Mediterranean dusky grouper Epinephelus marginatus (Lowe, 1834) (Pisces, Serranidae) in the Medes Islands Marine Reserve (NW Mediterranean, Spain). Sci. Mar. 1997, 61, 79–89. [Google Scholar]
- Machado, L.F.; Bertoncini, Á.A.; Hostim-Silva, M.; Barreiros, J.P. Habitat use by the juvenile dusky grouper Epinephelus marginatus and its relative abundance, in Santa Catarina, Brazil. Aqua J. Ichthyol. Aquat. Biol. 2003, 6, 133–138. [Google Scholar]
- Hereu, B.; Diaz, D.; Pasqual, J.; Zabala, M.; Sala, E. Temporal patterns of spawning of the dusky grouper Epinephelus marginatus in relation to environmental factors. Mar. Ecol. Prog. Ser. 2006, 325, 187–194. [Google Scholar] [CrossRef]
- Reñones, O.; Grau, A.; Mas, X.; Riera, F.; Saborido-Rey, F. Reproductive pattern of an exploited dusky grouper Epinephelus marginatus (Lowe 1834)(Pisces: Serranidae) population in the western Mediterranean. Sci. Mar. 2010, 74, 523–537. [Google Scholar] [CrossRef] [Green Version]
- Begossi, A.; Salyvonchyk, S.; Glamuzina, B.; de Souza, S.P.; Lopes, P.F.M.; Priolli, R.H.G.; do Prado, D.O.; Ramires, M.; Clauzet, M.; Zapelini, C.; et al. Fishers and groupers (Epinephelus marginatus and E. morio) in the coast of Brazil: Integrating information for conservation. J. Ethnobiol. Ethnomed. 2019, 15, 53. [Google Scholar] [CrossRef] [PubMed]
- La Mesa, G.; Vacchi, M. An Analysis of the Coastal Fish Assemblage of the Ustica Island Marine Reserve (Mediterranean Sea). Mar. Ecol. 1999, 20, 147–165. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.; García-Charton, J.A.; Bayle-Sempere, J.; Charbonnel, E.; Le Diréach, L.; Ody, D.; Pérez-Ruzafa, A.; Reñones, O.; Sánchez-Jerez, P.; Valle, C. Importance of marine reserves for the population dynamics of groupers (Epinephelinae) in the Western Mediterranean. In Proceedings of the 2nd Symposium on Mediterranean Groupers, Nice, France, 10–13 May 2007; pp. 91–93. [Google Scholar]
- Di Franco, A.; Bussotti, S.; Navone, A.; Panzalis, P.; Guidetti, P. Evaluating effects of total and partial restrictions to fishing on Mediterranean rocky-reef fish assemblages. Mar. Ecol. Prog. Ser. 2009, 387, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Harmelin, J.-G.; Ruitton, S. GEM 2010—Statut du mérou brun (Epinephelus marginatus) dans le Parc national de Port-Cros (France, Méditerranée): État 2008 et évolution en 15 ans. Sci. Rep. Port-Cros Natl. Park 2010, 24, 147–159. [Google Scholar]
- García-Rubies, A.; Hereu, B.; Zabala, M. Long-Term Recovery Patterns and Limited Spillover of Large Predatory Fish in a Mediterranean MPA. PLoS ONE 2013, 8, e73922. [Google Scholar] [CrossRef] [Green Version]
- Reñones, O.; Goñi, R.; Pozo, M.; Deudero, S.; Moranta, J. Effects of protection on the demographic structure and abundance of Epinephelus marginatus (Lowe, 1834). Evidence from Cabrera Archipelago National Park (West-central Mediterranean). Mar. Life 1999, 9, 45–53. [Google Scholar]
- Direção-Geral de Recursos Naturais, Segurança e Serviços Marítimos; Lisbon, Portugal. Portuguese Fisheries Statistics, 1997–2017. Unpublished work.
- Silva, A.F.; MARE; Lisbon, Portugal. Professional Fishing Surveys in ‘Sudoeste Alentejano’ and ‘Costa Vicentina’ Marine Park (MARSW Project), 2019. Unpublished work.
- Lembo, G.; Fleming, I.A.; Økland, F.; Carbonara, P.; Spedicato, M.T. Site fidelity of the dusky grouper Epinephelus marginatus (Lowe, 1834) studied by acoustic telemetry. Mar. Life 1999, 9, 37–43. [Google Scholar]
- Lembo, G.; Spedicato, M.T.; Økland, F.; Carbonara, P.; Fleming, I.A.; McKinley, R.S.; Thorstad, E.B.; Sisak, M.; Ragonese, S. A wireless communication system for determining site fidelity of juvenile dusky groupers Epinephelus marginatus (Lowe, 1834) using coded acoustic transmitters. Hydrobiologia 2002, 483, 249–257. [Google Scholar] [CrossRef]
- Pastor, J.; Verdoit-Jarraya, M.; Astruch, P.; Dalias, N.; Nelva Pasqual, J.S.; Saragoni, G.; Lenfant, P. Acoustic telemetry survey of the dusky grouper (Epinephelus marginatus) in the Marine Reserve of Cerbère-Banyuls: Informations on the territoriality of this emblematic species. Comptes Rendus. Biol. 2009, 332, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Afonso, P.; Abecasis, D.; Santos, R.S.; Fontes, J. Contrasting movements and residency of two serranids in a small Macaronesian MPA. Fish. Res. 2016, 177, 59–70. [Google Scholar] [CrossRef]
- Desiderà, E.; Trainito, E.; Navone, A.; Blandin, R.; Magnani, L.; Panzalis, P.; Mazzoldi, C.; Guidetti, P. Using complementary visual approaches to investigate residency, site fidelity and movement patterns of the dusky grouper (Epinephelus marginatus) in a Mediterranean marine protected area. Mar. Biol. 2021, 168, 111. [Google Scholar] [CrossRef]
- Spedicato, M.T.; Carbonara, P.; Lembo, G. Insight into the homing behaviour of the dusky grouper (Epinephelus marginatus Lowe, 1834) around the island of Ustica, Italy. In Proceedings of the 5th Conference on Fish Telemetry held in Europe, Aquatic Telemetry: Advances and Applications, Ustica, Italy, 9–13 June 2003; pp. 103–109. [Google Scholar]
- La Mesa, G.; Longobardi, A.; Sacco, F.; Marino, G. First release of hatchery juveniles of the dusky grouper Epinephelus marginatus (Lowe, 1834) (Serranidae: Teleostei) at artificial reefs in the Mediterranean: Results from a pilot study. Sci. Mar. 2008, 72, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Gallego, M.Á.B.; Reyes, O.M.; Rodríguez, A.I.M. Repoblación y seguimiento con telemetría acústica del mero (Epinephelus marginatus) en el Paraje Natural de Acantilados de Maro-Cerro Gordo. Chron. Nat. 2013, 3, 83–91. [Google Scholar]
- Horta e Costa, B.; Sousa, I.; Belackova, A.; Henriques, N.S.; Rangel, M.; Oliveira, F.; Afonso, C.M.L.; Bentes, L.; Monteiro, P.; Quintella, B.R.; et al. Relatório de Síntese da Biodiversidade Marinha da Área Marinha do Parque Natural do Sudoeste Alentejano e Costa Vicentina; Technical Report nr. 1, MARSW Project; CCMAR, UAlgarve, MARE, Ulisboa; UÉvora: Faro, Portugal, 2018; pp. i–vii + 1–74 + Appendices; Available online: https://www.marsw.pt/downloads/repo/materiais_divulgacao/Relatorio_de_Sintese_da_Biodiversidade_Marinha_da_area_marinha_do_Parque_Natural_do_Sudoeste_Alentejano_e_Costa_Vicentina_Relatorio_tecnico_do_Projeto_MARSW-1_compressed.pdf (accessed on 6 April 2022).
- López, V.G.; Orvay, F.C. Preliminary data on the culture of juveniles of the dusky grouper, Epinephelus marginatus (Lowe, 1834). Hidrobiológica 2003, 13, 321–327. [Google Scholar]
- Pierre, S.; Gaillard, S.; Prevot-D’Alvise, N.; Aubert, J.; Rostaing-Capaillon, O.L.-T.; Daniel, L.-T.; Grillaska, J.-P. Grouper Aquaculture: Taiwan 5—Mediterranean 0. In Proceedings of the 2nd International Symposium on the Mediterranean Groupers, Nice, France, 10–13 May 2007; pp. 135–138. [Google Scholar]
- Mahé, K.; CoChard, M.-L.; Quéro, J.-C.; Sevin, K.; Bailly, N.; Tetard, A. First record of Epinephelus marginatus (Serranidae: Epinephelinae) in the eastern English Channel. Cybium 2012, 36, 485–486. [Google Scholar] [CrossRef]
- Gonçalves, J.M.S.; Oliveira, F.; Monteiro, P.; Bentes, L.; Andrade, M.; Belackova, A.; Afonso, C.M.L.; Sousa, I.; Henriques, N.S.; Rangel, M. Mapeamento de Habitats e de Espécies na Área Marinha do PNSACV; Technical Report nr. 6, MARSW Project; CCMAR, UAlgarve, MARE, Ulisboa; UÉvora: Faro, Portugal, 2021; pp. i–xiii + 1–288 + Appendices; Available online: https://www.marsw.pt/downloads/repo/materiais_divulgacao/h._Mapeamento.pdf (accessed on 6 April 2022).
- Fallet, J. Evaluate the protection effect of a no-take zone with experimental fishing in the ‘Sudoeste Alentejano’ and ‘Costa Vicentina’ Natural Park. Master’s Thesis, Aix-Marseille University, Marseille, France, 2021. [Google Scholar]
- Pereira, T.; Manique, J.; Quintella, B.; Castro, N.; Almeida, P.; Costa, J. Changes in fish assemblage structure after implementation of Marine Protected Areas in the south western coast of Portugal. Ocean Coast. Manag. 2017, 135, 103–112. [Google Scholar] [CrossRef]
- Pereira, T.J.; Silva, A.F.; de Almeida, P.R.; Belo, A.F.; Costa, J.L.; Castro, N.; Quintella, B.R. Assessing the size adequacy of a small no-take marine protected area (MPA) for Mediterranean moray and European conger. Mar. Ecol. Prog. Ser. 2017, 584, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Belackova, A. Evaluation of the reserve effect in a Marine Protected Area in Sagres (PNSACV). Master’s Dissertation, Algarve University, Faro, Portugal, 2019. [Google Scholar]
- Cabrita, E.; Engrola, S.; Conceição, L.E.C.; Pousão-Ferreira, P.; Dinis, M.T. Successful cryopreservation of sperm from sex-reversed dusky grouper, Epinephelus marginatus. Aquaculture 2009, 287, 152–157. [Google Scholar] [CrossRef]
- Cunha, M.E.; Quental, H.; Barradas, A.; Pousão-Ferreira, P.; Cabrita, E.; Engrola, S. Rearing larvae of dusky grouper, Epinephelus marginatus (Lowe, 1834), (Pisces: Serranidae) in a semi-extensive mesocosm. Sci. Mar. 2009, 73, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Dingle, H. Migration: The Biology of Life on the Move, 2nd ed.; Oxford University Press: New York, NY, USA, 2014; Chapter 1; pp. 1–12. [Google Scholar] [CrossRef]
- Burt, W.H. Territoriality and Home Range Concepts as Applied to Mammals. J. Mammal. 1943, 24, 346–352. [Google Scholar] [CrossRef]
- Ketterson, E.D.; Nolan, V., Jr. Site Attachment and Site Fidelity in Migratory Birds: Experimental Evidence from the Field and Analogies from Neurobiology. In Bird Migration, 1st ed.; Gwinner, E., Ed.; Springer: Berlin/Heidelberg, Germany, 1990; pp. 117–129. ISBN 978-3-642-74542-3. [Google Scholar]
- OAL. Crepúsculos, Duração Do Dia, Alturas e Azimutes Do Sol (Lisboa). Available online: http://oal.ul.pt/documentos/2018/12/creplis2019.pdf/ (accessed on 28 January 2022).
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. Guide to Software and Statistical Methods PRIMER-E, 1st ed.; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Niella, Y.; Flávio, H.; Smoothey, A.F.; Aarestrup, K.; Taylor, M.D.; Peddemors, V.M.; Harcourt, R. Refined Shortest Paths (RSP): Incorporation of topography in space use estimation from node-based telemetry data. Methods Ecol. Evol. 2020, 11, 1733–1742. [Google Scholar] [CrossRef]
- Jacoby, D.M.P.; Brooks, E.J.; Croft, D.P.; Sims, D.W. Developing a deeper understanding of animal movements and spatial dynamics through novel application of network analyses. Methods Ecol. Evol. 2012, 3, 574–583. [Google Scholar] [CrossRef]
- Belo, A.F.; Pereira, T.J.; Quintella, B.R.; Castro, N.; Costa, J.L.; de Almeida, P.R. Movements of Diplodus sargus (Sparidae) within a Portuguese coastal Marine Protected Area: Are they really protected? Mar. Environ. Res. 2016, 114, 80–94. [Google Scholar] [CrossRef]
- CoastNet Project. CoastNet Project Geoportal. Available online: http://geoportal.coastnet.pt/ (accessed on 15 August 2019).
- Gallego, M.; Reyes, O.M. La población de meros Epinephelus marginatus en el litoral andaluz (España). Chron. Nat. 2015, 5, 68–80. [Google Scholar]
- Bodilis, P.; Ganteaume, A.; Francour, P. Recruitment of the dusky grouper (Epinephelus marginatus) in the north-western Mediterranean Sea. Cybium 2003, 27, 123–129. [Google Scholar] [CrossRef]
- Maggio, T.; Andaloro, F.; Arculeo, M. Genetic population structure of Epinephelus marginatus (Pisces, Serranidae) revealed by two molecular markers. Ital. J. Zool. 2006, 73, 275–283. [Google Scholar] [CrossRef]
- Chauvet, C.; Francour, P. Les mérous du parc national de Port-Cros: Aspects socio-démographiques. Bull. Soc. Zool. Fr. 1990, 114, 5–13. [Google Scholar]
- Derbal, F.; Kara, M.H. Habitat et comportement du mérou Epinephelus marginatus dans la région d’Annaba (Algérie). Cah. Biol. Mar. 1995, 36, 29–32. [Google Scholar]
- Harmelin, J.-G.; Harmelin-Vivien, M. A review on habitat, diet and growth of the dusky grouper Epinephelus marginatus (Lowe, 1834). Mar. Life 1999, 9, 11–20. [Google Scholar]
- La Mesa, G.; Louisy, P.; Vacchi, M. Assessment of microhabitat preferences in juvenile dusky grouper (Epinephelus marginatus) by visual sampling. Mar. Biol. 2002, 140, 175–185. [Google Scholar] [CrossRef]
- Sousa, I.I. Aspectos da Variabilidade na Abundância de Juvenis de Mero (Epinephelus marginatus) (Lowe, 1834) (Pisces, Serranidae) na Costa Sul da Ilha Terceira, Açores. Bachelor’s Thesis, Universidade do Algarve, Faro, Portugal, 2005. [Google Scholar]
- Gibran, F. Activity, habitat use, feeding behavior, and diet of four sympatric species of Serranidae (Actinopterygii: Perciformes) in southeastern Brazil. Neotrop. Ichthyol. 2007, 5, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Berastegui, D.; Coll, J.; Rueda, L.; Stobart, B.; Morey, G.; Navarro, O.; Aparicio-González, A.; Grau, A.M.; Reñones, O. Multiscale seascape habitat of necto-benthic littoral species, application to the study of the dusky grouper habitat shift throughout ontogeny. Mar. Environ. Res. 2018, 142, 21–31. [Google Scholar] [CrossRef]
- Derbal, F.; Kara, M. Alimentation estivale du mérou, Epinephelus marginatus (Serranidae), des côtes est algériennes. Cybium 1996, 20, 295–301. [Google Scholar]
- Barreiros, J.; Santos, R. Notes on the food habits and predatory behaviour of the dusky grouper, Epinephelus marginatus (Lowe, 1834) (Pisces: Serranidae) in the Azores. Arquipelago (Cienc. Biol. Mar./Life Mar. Sci.) 1998, 16A, 29–35. [Google Scholar]
- Reñones, O.; Polunin, N.V.C.; Goni, R. Size related dietary shifts of Epinephelus marginatus in a western Mediterranean littoral ecosystem: An isotope and stomach content analysis. J. Fish Biol. 2002, 61, 122–137. [Google Scholar] [CrossRef]
- Linde, M.; Grau, A.; Riera, F.; Enric, M.-P. Analysis of trophic ontogeny in Epinephelus marginatus (Serranidae). Cybium 2004, 28, 27–35. [Google Scholar] [CrossRef]
- López, V.; Castelló, F. Food habits of groupers Epinephelus marginatus (Lowe, 1834) and Epinephelus costae (Steindachner, 1878) in the Mediterranean Coast of Spain. Hidrobiológica 2005, 15, 27–34. [Google Scholar]
- Lelong, P. Identification individuelle du mérou brun, Epinephelus marginatus (Lowe, 1834) par les taches céphaliques. Mar. Life 1999, 9, 29–35. [Google Scholar]
- Gerhardinger, L.; Hostim-Silva, M.; Samagaia, R.; Barreiros, J. A following association between juvenile Epinephelus marginatus (Serranidae) and Myrichthys ocellatus (Ophichthidae). Cybium 2006, 30, 82–84. [Google Scholar]
- Azzurro, E.; Pais, A.; Consoli, P.; Andaloro, F. Evaluating day–night changes in shallow Mediterranean rocky reef fish assemblages by visual census. Mar. Biol. 2007, 151, 2245–2253. [Google Scholar] [CrossRef]
- Olla, B.L.; Davis, M.W.; Ryer, C.H. Understanding how the Hatchery Environment Represses or Promotes the Development of Behavioral Survival Skills. Bull. Mar. Sci. 1998, 62, 531–550. [Google Scholar]
- Henderson, H.F. Behavioral adjustment of fishes to release into a new habitat. In Proceedings of the Conference of International Center for Living Aquatic Resources Management on the Physiological and Behavioral Manipulation of Food Fish as Production and Management Tools, Bellagio, Italy, 3–8 November 1977; pp. 331–341. [Google Scholar]
- De Oliveira, N.Y.; de Sousa, O.M.; Kuhnen, V.V.; Sanches, E.G. Shelters influence in dusky grouper culture. Bol. Inst. Pesca 2019, 45, e473. [Google Scholar] [CrossRef]
- Caddy, J.F. Marine Habitat and Cover: Their Importance for Productive Coastal Fishery Resources; UNESCO Publishing: Paris, France, 2007; ISBN 978-92-3-104035-1. [Google Scholar]
- Caddy, J.F. Why do assessments of demersal stocks largely ignore habitat? ICES J. Mar. Sci. 2014, 71, 2114–2126. [Google Scholar] [CrossRef] [Green Version]
- Relini, G.; Relini, M.; Torchia, G.; Palandri, G. Ten years of censuses of fish fauna on the Loano artificial reef. ICES J. Mar. Sci. 2002, 59, S132–S137. [Google Scholar] [CrossRef] [Green Version]
- Selfati, M.; El Ouamari, N.; Lenfant, P.; Fontcuberta, A.; Lecaillon, G.; Mesfioui, A.; Boissery, P.; Bazairi, H. Promoting restoration of fish communities using artificial habitats in coastal marinas. Biol. Conserv. 2018, 219, 89–95. [Google Scholar] [CrossRef]
- Androulakis, D.N.; Dounas, C.G.; Banks, A.C.; Magoulas, A.N.; Margaris, D.P. An Assessment of Computational Fluid Dynamics as a Tool to Aid the Design of the HCMR-Artificial-ReefsTM Diving Oasis in the Underwater Biotechnological Park of Crete. Sustainability 2020, 12, 4847. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish ID | Total Length (mm) | Weight (g) | Transmitter ID/Model 1 | TBWR (%) | Release Site | Release Date/Time (UTC) |
|---|---|---|---|---|---|---|
| G1 | 542 | 2995 | 8041/V16-6H | 0.5 | R2 | 22 May 2019 12:00 |
| G2 | 538 | 2802 | 8040/V16-6H | 0.5 | R2 | 22 May 2019 12:00 |
| G3 | 515 | 2325 | 8039/V16-6H | 0.6 | R2 | 22 May 2019 12:00 |
| G4 | 508 | 2814 | 8038/V16-6H | 0.5 | R2 | 22 May 2019 12:00 |
| G5 | 500 | 2480 | 8037/V16-6H | 0.6 | R1 | 22 May 2019 14:00 |
| G6 | 518 | 2481 | 8036/V16-6H | 0.6 | R1 | 22 May 2019 14:00 |
| G7 | 495 | 2217 | 8035/V16-6H | 0.7 | R2 | 22 May 2019 12:00 |
| G8 | 517 | 2434 | 8034/V16-6H | 0.6 | R1 | 22 May 2019 14:00 |
| G9 | 517 | 2666 | 8042/V16-6H | 0.6 | R2 | 22 May 2019 12:00 |
| G10 | 488 | 2362 | 8043/V16-6H | 0.6 | R1 | 22 May 2019 14:00 |
| G11 | 515 | 2609 | 22984/V13-1L | 0.2 | R2 | 22 May 2019 12:00 |
| G12 | 489 | 1925 | 22983/V13-1L | 0.3 | R1 | 22 May 2019 14:00 |
| G13 | 502 | 2481 | 22982/V13-1L | 0.3 | R1 | 22 May 2019 14:00 |
| G14 | 500 | 2470 | 22976/V13-1L | 0.3 | R1 | 22 May 2019 14:00 |
| G15 | 530 | 2700 | 22977/V13-1L | 0.2 | R1 | 22 May 2019 14:00 |
| G16 | 513 | 2513 | 22978/V13-1L | 0.3 | R1 | 22 May 2019 14:00 |
| G17 | 486 | 2083 | 22979/V13-1L | 0.3 | R2 | 22 May 2019 12:00 |
| G18 | 512 | 2678 | 22980/V13-1L | 0.2 | R2 | 22 May 2019 12:00 |
| G19 | 486 | 2137 | 22970/V13-1L | 0.3 | R2 | 22 May 2019 12:00 |
| G20 | 495 | 2078 | 22981/V13-1L | 0.3 | R2 | 22 May 2019 12:00 |
| G21 | 650 | 5200 | 11418/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G22 | 650 | 5150 | 11429/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G23 | 620 | 4953 | 11428/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G24 | 615 | 5232 | 11430/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G25 | 650 | 5738 | 11431/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G26 | 640 | 6480 | 11432/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G27 | 610 | 5220 | 11433/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G28 | 560 | 3430 | 11424/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G29 | 590 | 3690 | 11425/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| G30 | 580 | 4250 | 11426/V9-2L | <0.1 | R3 | 19 April 2021 12:00 |
| Variable | Factor | Df | SS | MS | Pseudo-F | p | Perm |
|---|---|---|---|---|---|---|---|
| IA | Day period | 3 | 22,362 | 7454 | 3.0708 | 0.010 | 997 |
| NR | 3 | 35,331 | 11,777 | 5.7491 | 0.001 | 997 |
| Day Period | Total nr. Detections | Time per Day (min) 1 | IA (%) 2 | NR 2 |
|---|---|---|---|---|
| Dawn | 26 | 75 | 0 | 0 |
| Daytime | 799 | 900 | 2.90 | 0.14 |
| Dusk | 258 | 75 | 22.59 | 1.45 |
| Night | 298 | 390 | 5.20 | 0.56 |
| Variable | Factor Levels | Dawn | Daytime | Dusk | Night |
|---|---|---|---|---|---|
| IA | Dawn | - | 0.010 | 0.024 | 0.211 |
| Daytime | - | - | 0.026 | 0.151 | |
| Dusk | - | - | - | 0.509 | |
| Night | - | - | - | - | |
| NR | Dawn | - | 0.004 | 0.020 | 0.013 |
| Daytime | - | - | 0.001 | 0.038 | |
| Dusk | - | - | - | 0.197 | |
| Night | - | - | - | - |
| Study Area | Grouper 1 | Distance Traveled (m) 2 | Time Traveled (hh:mm) 3 | Ranging Speed (m.h−1) |
|---|---|---|---|---|
| Pessegueiro island | G1 | 4379.77 | 3:56 | 1113.48 |
| G2 | 911.87 | 1:12 | 755.52 | |
| G3 | 5340.70 | 20:55 | 255.33 | |
| G4 | - | - | - | |
| G5 | 8233.89 | 6:09 | 1338.84 | |
| G6 | 734.91 | 3:24 | 216.15 | |
| G7 | - | - | - | |
| G8 | 734.91 | 30:31 | 24.08 | |
| G9 | - | - | - | |
| G10 | 3744.90 | 8:05 | 463.29 | |
| G11 | 741.61 | 5:59 | 123.95 | |
| G12 | 1158.21 | 16:52 | 69.13 | |
| G13 | 734.91 | 3:29 | 210.98 | |
| G14 | 747.60 | 0:35 | 1281.60 | |
| G15 | 747.60 | 25:49 | 694.99 | |
| G16 | 734.91 | 0:51 | 864.60 | |
| G17 | 1823.74 | 0:38 | 2879.59 | |
| G18 | 911.87 | 0:39 | 1402.88 | |
| G19 | 2631.52 | 2:45 | 956.92 | |
| G20 | 734.91 | 26:10 | 28.09 | |
| Martinhal islets | G21 | 3421.87 | 29:37 | 115.54 |
| G22 | 3344.15 | 32:55 | 101.59 | |
| G23 | 7594.19 | 16:06 | 471.69 | |
| G24 | 4883.05 | 13:46 | 354.33 | |
| G25 | 2011.72 | 11:11 | 179.69 | |
| G26 | 1473.46 | 2:52 | 513.90 | |
| G27 | 1183.33 | 7:27 | 158.74 | |
| G28 | - | - | - | |
| G30 | 6035.60 | 33:53 | 178.07 | |
| Mean | 2547.60 | 12:14 | 590.12 |
| Grouper. | Movement 1 (A → B) | Type of Reported Event 2,3 (A—B) | Date Time of Reported Event 4 (A–B) | Minimum Distance Traveled (km) | Maximum Time Traveled (Hours) 5 | Ground Speed (m.h−1) |
|---|---|---|---|---|---|---|
| G1 | 1 → 2 | detection—recapture | 23 May 2019 21:19–30 May 2019 | 23.3 | - | - |
| G3 | 3 → 4 | detection—recapture | 23 May 2019 20:38–4 June 2019 | 3.5 | - | - |
| G5 | 3 → 5 | detection—detection | 22 May 2019 21:08–26 May 2019 21:13 | 70.0 | 96.07 | 730 |
| 5 → 6 | detection—detection | 26 May 2019 21:13–28 May 2019 21:36 | 17.4 | 48.38 | 360 | |
| 87.4 | 144.45 | - | ||||
| G6 | 3 → 7 | detection—recapture | 22 May 2019 20:43–17 June 2019 | 25.0 | - | - |
| G7 | 3 → 8 | detection—recapture 6 | 28 May 2019 21:48–24 June 2019 | 16.0 | - | - |
| G8 | 3 → 9 | detection—detection | 23 May 19 20:26–27 May 2019 01:32 | 51.5 | 77.08 | 670 |
| G10 | 3 → 9 | detection—detection | 22 May 2019 22:06–26 May 2019 20:22 | 49.0 | 94.25 | 520 |
| 9 → 3 | detection—detection | 27 May 2019 01:32–29 May 2019 21:33 | 52.0 | 68.00 | 760 | |
| 3 → 8 | detection—detection 7 | 29 May 2019 22:07–7 September 2019; 21 September 2019; 6 January 2020 | 16.0 | - | - | |
| 117.0 | - | - | ||||
| G17 | 3 → 5 | detection—detection | 22 May 2019 15:52–26 May 2019 21:18 | 70.0 | 101.42 | 690 |
| 5 → 6 | detection—detection | 26 May 2019 21:18–28 May 2019 01:36 | 17.4 | 28.30 | 620 | |
| 6 → 10 | detection—detection | 28 May 2019 01:36–31 May 2019 22:56 | 28.6 | 93.32 | 310 | |
| 10 → 11 | detection—recapture | 2 June 2019 02:01–29 July 2019 | 52.0 | - | - | |
| 168.0 | - | - | ||||
| G18 | 3 → 5 | detection—detection | 22 May 2019 21:31–1 June 2019 02:44 | 70.0 | 221.20 | 320 |
| 5 → 6 | detection—detection | 1 June 2019 02:44–2 June 2019 21:28 | 17.4 | 42.70 | 410 | |
| 87.4 | 263.90 | - | ||||
| G22 | 12 → 13 | detection—recapture | 20 April 2021 21:15–June 2021 | 21.0 | - | - |
| G23 | 14 → 15 | detection—detection | 22 April 2021 22:20–15 September 2021 19:55 | 113.0 | 3501.58 | 30 |
| G26 | 12 → 16 | detection—detection | 19 April 2021 14:51–28 May 2021 01:27 | 30.8 | 922.60 | 30 |
| 16 → 12 | detection—detection | 28 May 2021 01:48–29 May 2021 01:09 | 29.0 | 23.33 | 1240 | |
| 59.8 | - | - | ||||
| G30 | 12 → 13 | detection—recapture | 20 April 2021 21:56–June 2021 | 21.0 | - | - |
| Mean | 61.1 | - | 540 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.F.; Horta e Costa, B.; Costa, J.L.; Pereira, E.; Marques, J.P.; Castro, J.J.; Lino, P.G.; Candeias-Mendes, A.; Pousão-Ferreira, P.; Sousa, I.; et al. Movements of Hatchery-Reared Dusky Groupers Released in a Northeast Atlantic Coastal Marine Protected Area. J. Mar. Sci. Eng. 2022, 10, 904. https://doi.org/10.3390/jmse10070904
Silva AF, Horta e Costa B, Costa JL, Pereira E, Marques JP, Castro JJ, Lino PG, Candeias-Mendes A, Pousão-Ferreira P, Sousa I, et al. Movements of Hatchery-Reared Dusky Groupers Released in a Northeast Atlantic Coastal Marine Protected Area. Journal of Marine Science and Engineering. 2022; 10(7):904. https://doi.org/10.3390/jmse10070904
Chicago/Turabian StyleSilva, Ana Filipa, Bárbara Horta e Costa, José Lino Costa, Esmeralda Pereira, João Pedro Marques, João J. Castro, Pedro G. Lino, Ana Candeias-Mendes, Pedro Pousão-Ferreira, Inês Sousa, and et al. 2022. "Movements of Hatchery-Reared Dusky Groupers Released in a Northeast Atlantic Coastal Marine Protected Area" Journal of Marine Science and Engineering 10, no. 7: 904. https://doi.org/10.3390/jmse10070904