
Evolutionary Rates in the Haptophyta: Exploring Molecular and Phenotypic Diversity


Abstract
:1. Introduction


2. Molecular Diversity and Phylogeny of the Haptophyta
2.1. Haptophytes in Brief: Main Phylogenetic Clades
2.2. Molecular Phylogeny: Consistent Groupings and Hidden Surprises
2.3. Molecular Clock Estimates
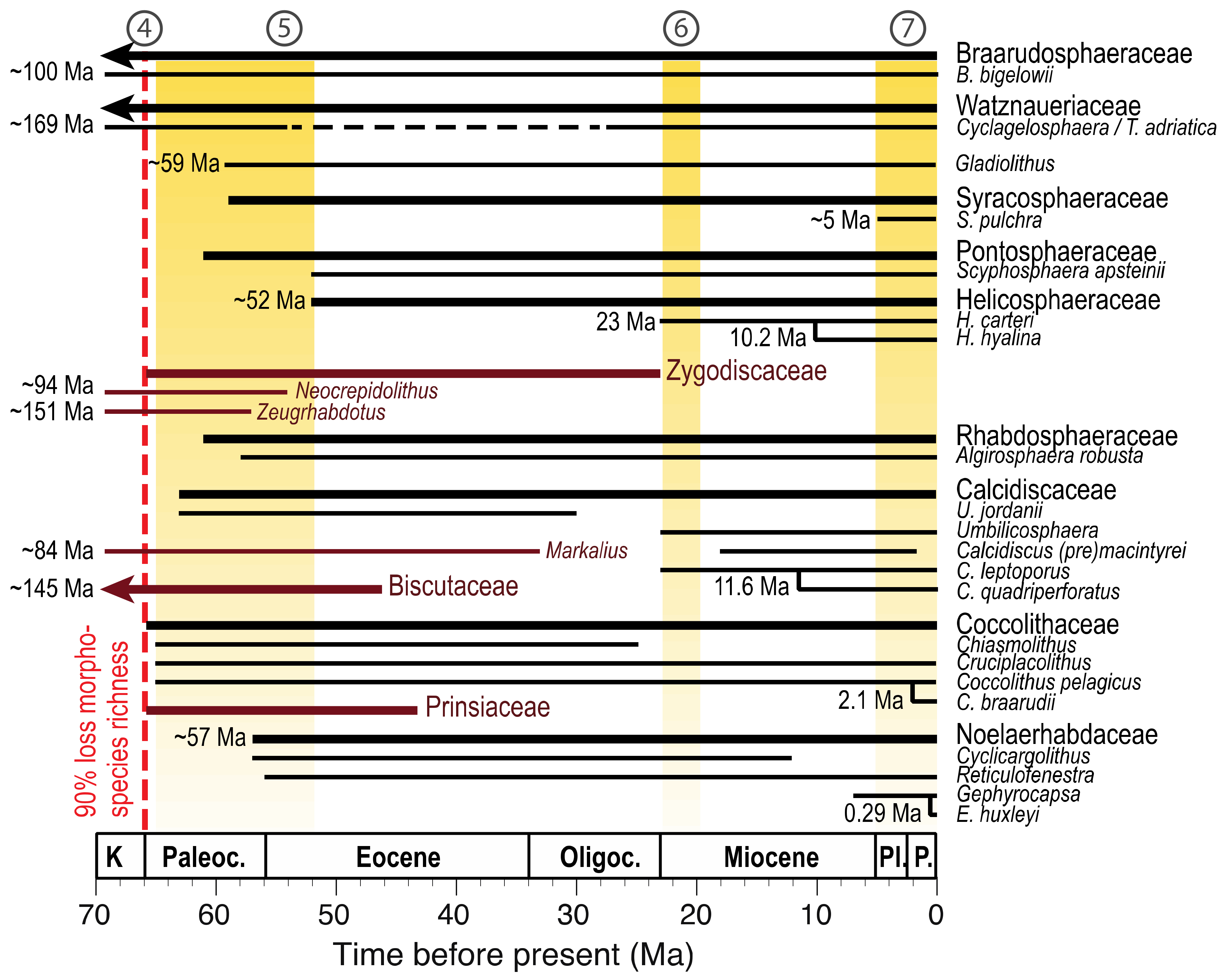
3. The Fossil Record of Coccolithophores
3.1. Telling Time: The Dates of Ancient History
3.2. The Fossil Lineages of Extant Clades
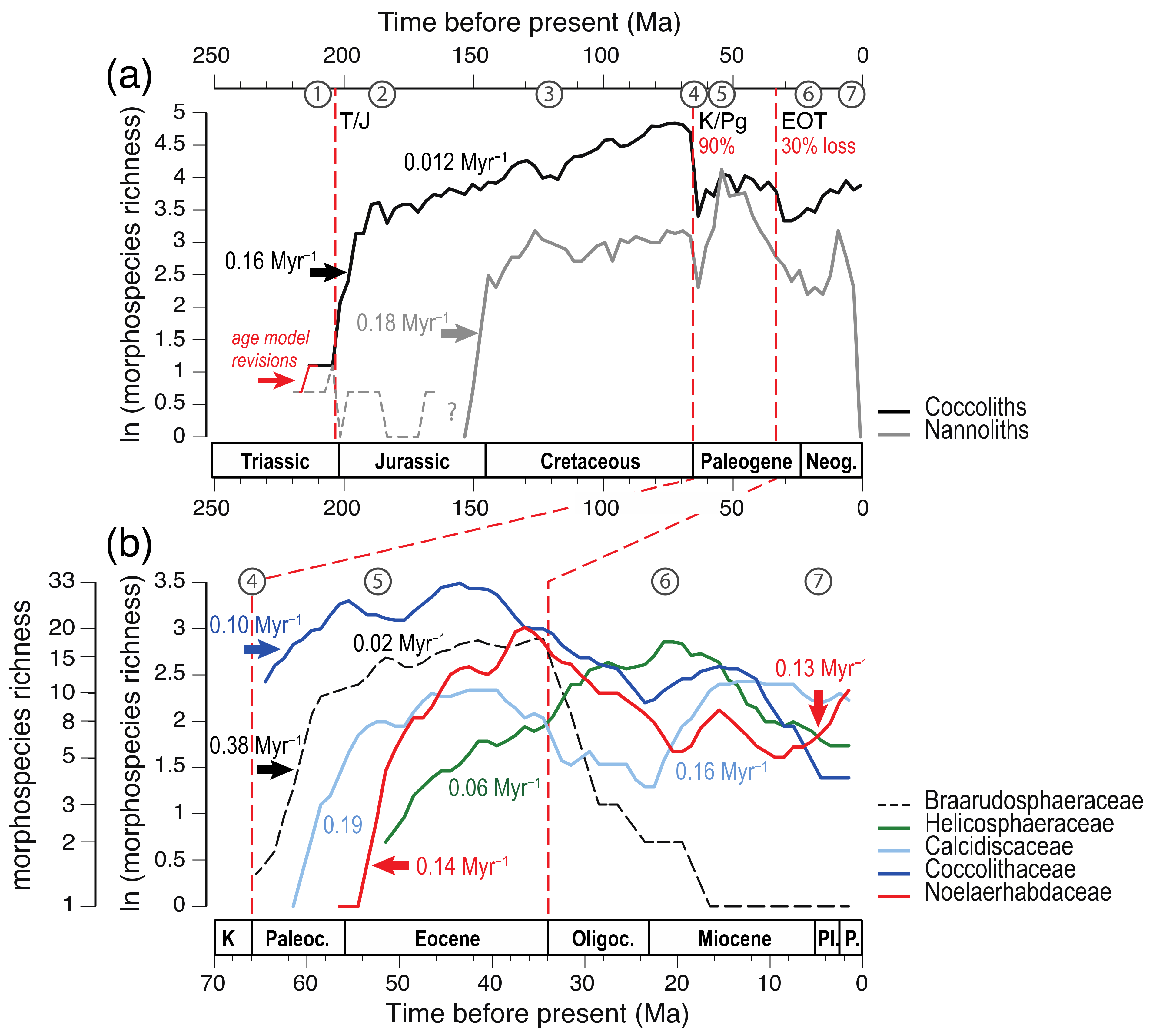
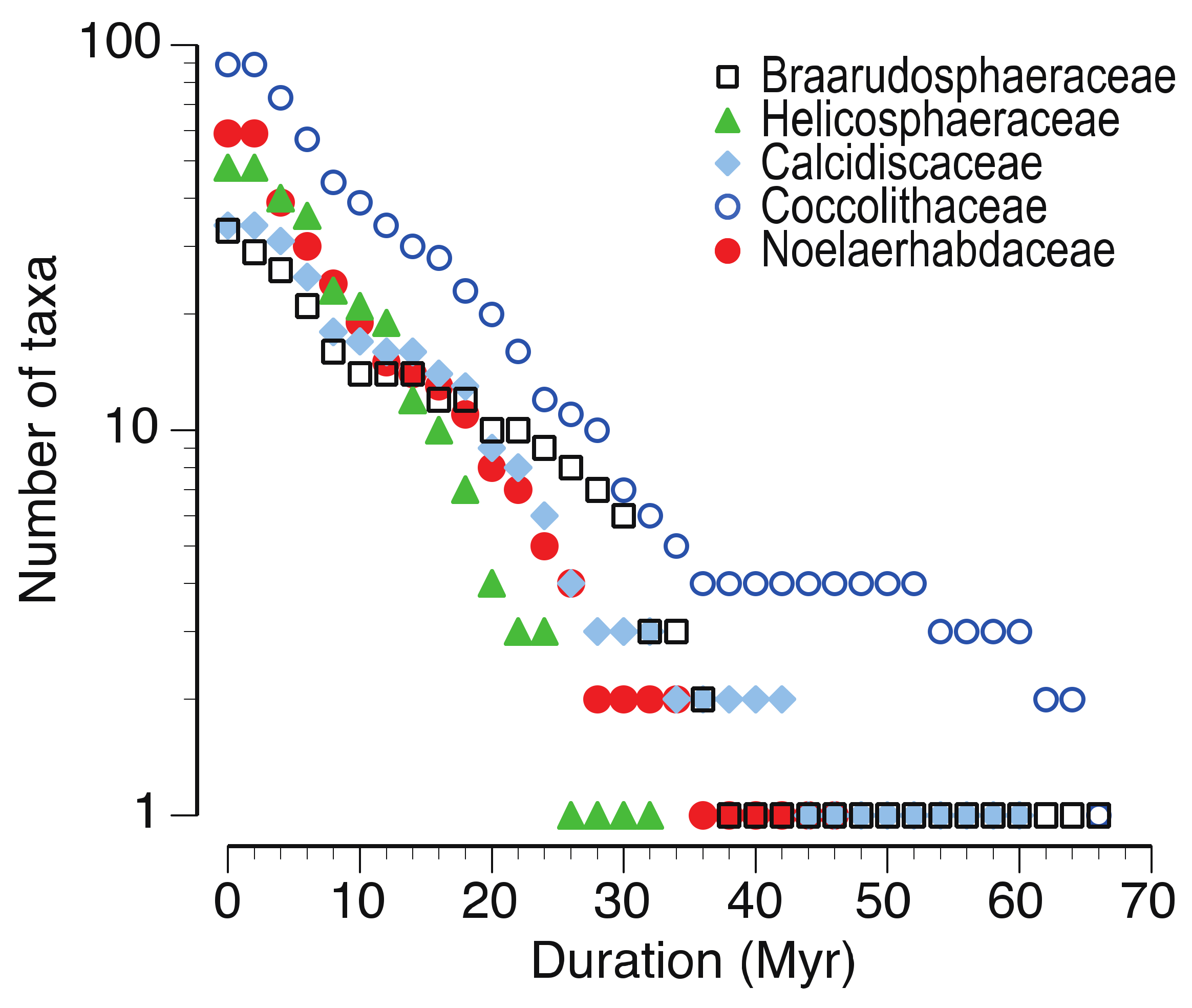
3.3. The Adaptive Radiations That Established Modern Clades
3.4. Modes of Speciation in Coccolithophores
4. Origins and Evolutionary History of Key Haptophyte Traits
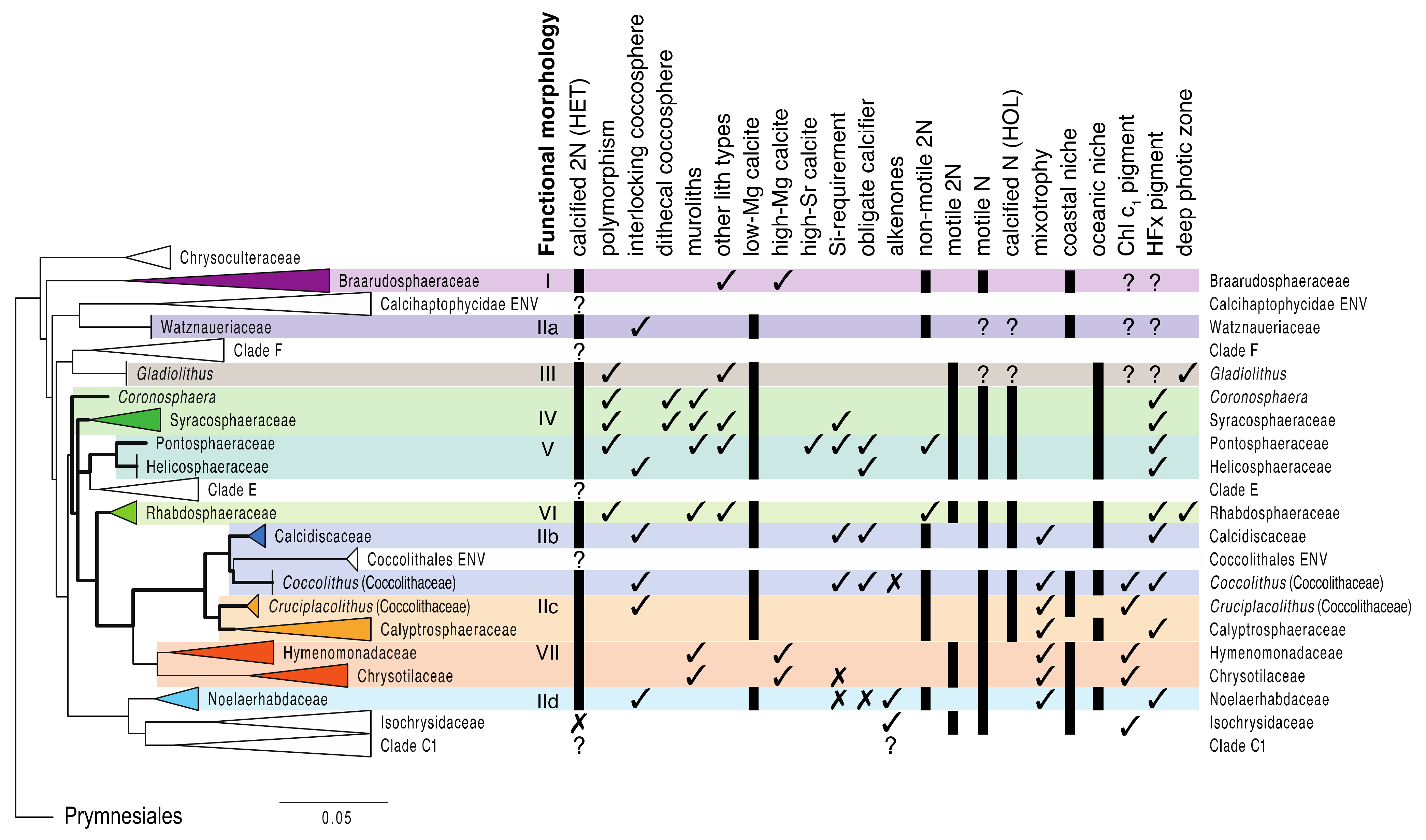
4.1. The Origin and Evolution of Calcification in Haptophytes
4.2. Functional Morphology: Exploring the Traitscape of Coccolithophores
5. Evolutionary Benefits of the Haplo-Diplontic Life Cycle
5.1. What Is the Haplo-Diplontic Life Cycle Good for?
5.2. How Do Coccolithophores Live Their Haplo-Diplontic Lifestyle?
5.3. Sexual vs. Asexual Reproduction
6. Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karlson, B.; Andersen, P.; Arneborg, L.; Cembella, A.; Eikrem, W.; John, U.; West, J.J.; Klemm, K.; Kobos, J.; Lehtinen, S.; et al. Harmful algal blooms and their effects in coastal seas of Northern Europe. Harmful Algae 2021, 102, 101989. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.W.; Yoder, J.A. Coccolithophorid blooms in the global ocean. J. Geophys. Res. 1994, 99, 7467–7482. [Google Scholar] [CrossRef]
- Rousseaux, S.C.; Gregg, W.W. Interannual variation in phytoplankton primary production at a global scale. Remote Sens. 2014, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Poulton, A.J.; Adey, T.R.; Balch, W.M.; Holligan, P.M. Relating coccolithophore calcification rates to phytoplankton community dynamics: Regional differences and implications for carbon export. Deep.-Sea Res. II 2007, 54, 538–557. [Google Scholar] [CrossRef]
- Frada, M.J.; Bendif, E.M.; Keuter, S.; Probert, I. The private life of coccolithophores. Perspect. Phycol. 2018, 6, 11–30. [Google Scholar] [CrossRef]
- Eikrem, W.; Medlin, L.K.; Henderiks, J.; Rokitta, S.; Rost, B.; Probert, I.; Throndsen, J.; Edvardsen, B. Haptophyta. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–61. [Google Scholar]
- Bendif, E.M.; Probert, I.; Hervé, A.; Billard, C.; Goux, D.; Lelong, C.; Cadoret, J.-P.; Véron, B. Integrative taxonomy of the Pavlovophyceae (Haptophyta): A reassessment. Protist 2011, 162, 738–761. [Google Scholar] [CrossRef]
- De Vries, J.; Monteiro, F.; Wheeler, G.; Poulton, A.; Godrijan, J.; Cerino, F.; Malinverno, E.; Langer, G.; Brownlee, C. Haplo-diplontic life cycle expands coccolithophore niche. Biogeosciences 2021, 18, 1161–1184. [Google Scholar] [CrossRef]
- Medlin, L.K.; Kooistra, W.H.C.F.; Potter, D.; Saanders, G.; Wandersen, R.A. Phylogenetic relationships of the ‘golden algae’ (haptophytes, heterokont chromophytes) and their plastids. In The Origin of the Algae and Their Plastids (Plant Systematics and Evolution Supplement); Bhattacharya, D., Ed.; Springer: New York, NY, USA, 1997; pp. 187–219. [Google Scholar]
- Medlin, L.K.; Sáez, A.G.; Young, J.R. A molecular clock for coccolithophores and implications for selectivity of phytoplankton extinctions across the K/T boundary. Mar. Micropaleontol. 2008, 67, 69–86. [Google Scholar] [CrossRef]
- Liu, H.; Aris-Brosou, S.; Probert, I.; de Vargas, C. A time line of the environmental genetics of the Haptophytes. Mol. Biol. Evol. 2010, 27, 161–176. [Google Scholar] [CrossRef] [Green Version]
- Bown, P.R.; Lees, J.A.; Young, J.R. Calcareous nannoplankton evolution and diversity through time. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 481–508. [Google Scholar]
- Gardin, S.; Krystyn, L.; Richoz, S.; Bartolini, A.; Galbrun, B. Where and when the earliest coccolithophores? Lethaia 2012, 45, 507–523. [Google Scholar] [CrossRef]
- Billard, C.; Inouye, I. What is new in coccolithophore biology? In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 1–29. [Google Scholar]
- Brownlee, C.; Wheeler, G.L.; Taylor, A.H. Coccolithophore biomineralization: New questions, new answers. Semin. Cell Dev. Biol. 2015, 46, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, C.; Langer, G.; Wheeler, G.L. Coccolithophore calcification: Changing paradigms in changing oceans. Acta Biomater. 2021, 120, 4–11. [Google Scholar] [CrossRef]
- Monteiro, F.M.; Bach, L.T.; Brownlee, C.; Bown, P.; Rickaby, R.E.M.; Poulton, A.J.; Tyrrell, T.; Beaufort, L.; Dutkiewicz, S.; Gibbs, S.; et al. Why marine phytoplankton calcify. Sci. Adv. 2016, 2, e1501822. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.R.; Brownlee, C.; Wheeler, G. Coccolithophore cell biology: Chalking up progress. Annu. Rev. Mar. Sci. 2017, 9, 283–310. [Google Scholar] [CrossRef] [Green Version]
- Young, J.R.; Bown, P.R.; Lees, J.A. Nannotax 3 (Online Resource and Guide to the Biodiversity and Taxonomy of Coccolithophores). Available online: https://www.mikrotax.org/Nannotax3 (accessed on 24 December 2021).
- Young, J.R.; Liu, H.; Probert, I.; Aris-Brosou, S.; de Vargas, C. Morphospecies versus phylospecies concepts for evaluating phytoplankton diversity: The case of the Coccolithophores. Cryptogam. Algol. 2014, 35, 353–377. [Google Scholar] [CrossRef]
- Gran-Stadniczenko, S.; Šupraha, L.; Egge, E.D.; Edvardsen, B. Haptophyte diversity and vertical distribution explored by 18S and 28S ribosomal RNA gene metabarcoding and scanning electron microscopy. J. Eukaryot. Microbiol. 2017, 64, 514–532. [Google Scholar] [CrossRef] [Green Version]
- Battacharya, D.; Medlin, L.K. The phylogeny of plastids: A review based on comparison of small-subunit ribosomal RNA coding regions. J. Phycol. 1995, 31, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Edvardsen, B.; Egge, E.S.; Vaulot, D. Diversity and distribution of haptophytes revealed by environmental sequencing and metabarcoding—A review. Perspect. Phycol. 2016, 3, 77–91. [Google Scholar] [CrossRef]
- Edvardsen, B.; Eikrem, W.; Green, J.C.; Andersen, R.A.; Moon-van der Staay, S.Y.; Medlin, L.K. Phylogenetic reconstructions of the Haptophyta inferred from 18S ribosomal DNA sequences and available morphological data. Phycologia 2000, 39, 19–35. [Google Scholar] [CrossRef]
- Edvardsen, B.; Medlin, L.K. Molecular systematics of the Haptophyta. In Unravelling the Algae—The Past, Present and Future of Algal Molecular Systematics; Lewis, J., Brodie, J., Eds.; The Systematics Association; Taylor and Francis: London, UK; New York, NY, USA, 2007; pp. 183–196. [Google Scholar]
- Egge, E.S.; Eikrem, W.; Edvardsen, B. Deep-branching novel lineages and high diversity of haptophytes in the Skagerrak (Norway) uncovered by 454 pyrosequencing. J. Eukaryot. Microbiol. 2015, 62, 121–140. [Google Scholar] [CrossRef]
- Fujiwara, S.; Tsuzuki, M.; Kawachi, M.; Minaka, N.; Inouye, I. Molecular phylogeny of the Haptophyta based on the rbcL gene and sequence variation in the spacer region of the Rubisco operon. J. Phycol. 2001, 37, 121–129. [Google Scholar] [CrossRef]
- Sáez, A.G.; Probert, I.; Geisen, M.; Quinn, P.; Young, J.; Medlin, L.K. Pseudo-cryptic speciation in coccolithophores. Proc. Natl. Acad. Sci. USA 2003, 100, 7163–7168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sáez, A.G.; Probert, I.; Young, J.R.; Edvardsen, B.; Eikrem, W.; Medlin, L.K. A review of the phylogeny of the Haptophyta. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 251–269. [Google Scholar]
- Medlin, L.K.; Barker, G.L.A.; Campbell, L.; Green, J.C.; Hayes, P.K.; Marie, D.; Wrieden, S.; Vaulot, D. Genetic characterisation of Emiliania huxleyi (Haptophyta). J. Mar. Syst. 1996, 9, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Hagino, K.; Takano, Y.; Horiguchi, T. Pseudo-cryptic speciation in Braarudosphaera bigelowii (Gran and Braarud) Deflandre. Mar. Micropaleontol. 2009, 72, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Van Bleijswijk, J.; van der Wal, P.; Kempers, R.; Veldhuis, M.; Young, J.; Muyzer, G.; de Vrind-de Jong, E.; Westbroek, P. Distribution of two types of Emiliania huxleyi (Prymnesiophyceae) in the Northeast Atlantic region as determined by immunofluorescence and coccolith morphology. J. Phycol. 1991, 27, 566–570. [Google Scholar] [CrossRef]
- Young, J.; Westbroek, P. Genotypic variation in the coccolithophorid species Emiliania huxleyi. Mar. Micropaleontol. 1991, 18, 5–23. [Google Scholar] [CrossRef]
- Schroeder, D.C.; Biggi, G.F.; Hall, M.; Davy, J.; Martínez Martínez, J.; Richardson, A.J.; Malin, G.; Wilson, W.H. A genetic marker to separate Emiliania huxleyi (Prymnesiophyceae) morphotypes. J. Phycol. 2005, 41, 874–879. [Google Scholar] [CrossRef]
- Iglesias-Rodríguez, M.D.; Schofield, O.; Batley, J.; Medlin, L.K.; Hayes, P.K. Intraspecific genetic diversity in the marine coccolithophore Emiliania huxleyi (Prymnesiophyceae): The use of microsatellite analysis in marine phytoplankton population studies. J. Phycol. 2006, 42, 526–536. [Google Scholar] [CrossRef]
- Hagino, K.; Bendif, E.M.; Young, J.; Kogame, K.; Probert, I.; Takano, H.; Horiguchi, T.; de Vargas, C.; Okada, H. New evidence for morphological and genetic variation in the cosmopolitan coccolithophore Emiliania huxleyi (Prymnesiophyceae) from the COX1b-ATP4 genes. Phycologia 2011, 47, 1164–1176. [Google Scholar] [CrossRef]
- Cook, S.S.; Whittock, L.; Wright, S.W.; Hallegraeff, G.M. Photosynthetic pigment and genetic differences between two Southern Ocean morphotypes of Emiliania huxleyi (Haptophyta). J. Phycol. 2011, 47, 615–626. [Google Scholar] [CrossRef]
- Bendif, E.M.; Probert, I.; Carmichael, M.; Romac, S.; Hagino, K.; de Vargas, C. Genetic delineation between an within the widespread coccolithophore morpho-species Emiliania huxleyi and Gephyrocapsa oceanica (Haptophyta). J. Phycol. 2014, 50, 140–148. [Google Scholar] [CrossRef]
- Liu, H.; Probert, I.; Uitz, J.; Claustre, H.; Aris-Brosou, S.; Frada, M.; Not, F.; de Vargas, C. Extreme diversity in noncalcifying haptophytes explains a major pigment paradox in open oceans. Proc. Natl. Acad. Sci. USA 2009, 106, 12803–12808. [Google Scholar] [CrossRef] [Green Version]
- Jordan, R.W.; Cros, L.; Young, J.R. A revised classification scheme for living haptophytes. Micropaleontology 2004, 50, 55–79. [Google Scholar] [CrossRef]
- De Vargas, C.; Aubry, M.-P.; Probert, I.; Young, J. Origin and evolution of Coccolithophores: From coastal hunters to oceanic farmers. In Evolution of Primary Producers in the Sea; Falkowski, P.G., Knoll, A., Eds.; Academic Press: Cambridge, MA, USA, 2007; pp. 251–285. [Google Scholar]
- Klaveness, D.; Paasche, E. Two different Coccolithus huxleyi cell types incapable of coccolith formation. Arch. Mikrobiol. 1971, 75, 382–385. [Google Scholar] [CrossRef]
- Langer, G.; Taylor, A.R.; Walker, C.E.; Meyer, E.M.; Ben Joseph, O.; Gal, A.; Harper, G.M.; Probert, I.; Brownlee, C.; Wheeler, G.L. Role of silicon in the development of complex crystal shapes in coccolithophores. New Phytol. 2021, 231, 1845–1857. [Google Scholar] [CrossRef]
- Van der Wal, P.; de Jong, E.W.; Westbroek, P. Ultrastructural polysaccharide localization in calcifying and naked cells of the coccolithophorid Emiliania huxleyi. Protoplasma 1983, 118, 157–168. [Google Scholar] [CrossRef]
- Wilbur, K.M.; Watabe, N. Experimental studies on calcification in Molluscs and the alga Coccolithus huxleyi. Ann. N. Y. Acad. Sci. 1963, 109, 82–112. [Google Scholar] [CrossRef]
- Yoshida, M.; Noël, M.-H.; Nakayama, T.; Naganuma, T.; Inouye, I. A haptophyte bearing siliceous scales: Ultrastructure and phylogenetic position of Hyalolithus neolepis gen. et sp. nov. (Prymnesiophyceae, Haptophyta). Protist 2006, 157, 213–234. [Google Scholar] [CrossRef]
- De Vargas, C.; Probert, I. New keys to the Past: Current and future DNA studies in Coccolithophores. Micropaleontology 2004, 50, 45–54. [Google Scholar] [CrossRef]
- Perch-Nielsen, K. Mesozoic calcareous nannofossils. In Plankton Stratigraphy; Bolli, H.M., Saunders, J.B., Perch-Nielsen, K., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 329–426. [Google Scholar]
- Perch-Nielsen, K. Cenozoic calcareous nannofossils. In Plankton Stratigraphy; Bolli, H.M., Saunders, J.B., Perch-Nielsen, K., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 427–554. [Google Scholar]
- Aubry, M.-P. Phylogenetically based calcareous nannofossil taxonomy: Implications for the interpretation of geological events. In Nannofossils and Their Applications; Crux, J.A., Van Heck, S.E., Eds.; Ellis Horwood Limited: London, UK, 1989; pp. 21–40. [Google Scholar]
- Young, J.R.; Didymus, J.M.; Bown, P.R.; Prins, B.; Mann, S. Crystal assembly and phylogenetic evolution in heterococcoliths. Nature 1992, 356, 516–518. [Google Scholar] [CrossRef]
- Young, J.; Geisen, M.; Cros, L.; Kleijne, A.; Sprengel, C.; Probert, I.; Ostergaard, J. A guide to extant coccolithophore taxonomy. J. Nannoplankton Res. Spec. Issue 2003, 1, 1–132. [Google Scholar]
- Hagino, K.; Young, J.R.; Bown, P.R.; Godrijan, J.; Kulhanek, D.K.; Kogame, K.; Horiguchi, T. Re-discovery of a “living fossil” coccolithophore from the coastal waters of Japan and Croatia. Mar. Micropaleontol. 2015, 116, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Stanley, S.M.; Ries, J.B.; Hardie, L.A. Seawater chemistry, coccolithophore population growth, and the origin of Cretaceous chalk. Geology 2005, 33, 593–596. [Google Scholar] [CrossRef] [Green Version]
- Krasovec, M.; Rickaby, R.E.M.; Filatov, D.A. Evolution of mutation rate in astronomically large phytoplankton populations. Genome Biol. Evol. 2020, 12, 1051–1059. [Google Scholar] [CrossRef]
- Read, B.A.; Kegel, J.; Klute, M.J.; Kuo, A.; Lefebvre, S.C.; Maumus, F.; Mayer, C.; Miller, J.; Monier, A.; Salamov, A.; et al. Pan genome of the phytoplankton Emiliania underpins its global distribution. Nature 2013, 499, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Bendif, E.M.; Nevado, B.; Wong, E.L.Y.; Hagino, K.; Probert, I.; Young, J.R.; Rickaby, R.E.M.; Filatov, D.A. Repeated species radiations in the recent evolution of the key marine phytoplankton lineage Gephyrocapsa. Nat. Commun. 2019, 10, 4234. [Google Scholar] [CrossRef] [Green Version]
- Filatov, D.A.; Bendif, E.M.; Archontikis, O.A.; Hagino, K.; Rickaby, R.E.M. The mode of speciation during a recent radiation in open-ocean phytoplankton. Curr. Biol. 2021, 31, 5439–5449.e5. [Google Scholar] [CrossRef]
- Abe, K.; Tsutsui, H.; Jordan, R.W. Hyalolithus tumescens sp. nov., a siliceous scale-bearing haptophyte from the middle Eocene. J. Micropalaeontol. 2016, 35, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Geisen, M.; Young, J.R.; Probert, I.; Sáez, A.G.; Baumann, K.-H.; Sprengel, C.; Bollmann, J.; Cros, L.; de Vargas, C.; Medlin, L.K. Species level variation in coccolithophores. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 327–366. [Google Scholar]
- Young, J.; Geisen, M.; Probert, I. A review of selected aspects of coccolithophore biology with implications for paleobiodiversity estimation. Micropaleontology 2005, 51, 267–288. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Austin, J.A.J.; Exon, N.; Humphris, S.; Kastner, M.; McKenzie, J.A.; Miller, K.G.; Suyehiro, K.; Taira, A. Fifty years of scientific ocean drilling. Oceanography 2019, 32, 17–21. [Google Scholar] [CrossRef]
- Bown, P.R. Triassic. In Calcareous Nannofossil Biostratigraphy; Bown, P.R., Ed.; Chapman & Hall: Cambridge, UK, 1998; pp. 29–33. [Google Scholar]
- Bown, P.R.; Cooper, M.K.E. Jurassic. In Calcareous Nannofossil Biostratigraphy; Bown, P.R., Ed.; Chapman & Hall: Cambridge, UK, 1998; pp. 34–85. [Google Scholar]
- Burnett, J.A. Upper Cretaceous. In Calcareous Nannofossil Biostratigraphy; Bown, P.R., Ed.; Chapman & Hall: Cambridge, UK, 1998; pp. 132–199. [Google Scholar]
- Agnini, C.; Fornaciari, E.; Raffi, I.; Catanzariti, R.; Pälike, H.; Backman, J.; Rio, D. Biozonation and biochronology of Paleogene calcareous nannofossils from low and middle latitudes. Newsl. Stratigr. 2014, 47, 131–181. [Google Scholar] [CrossRef]
- Raffi, I.; Backman, J.; Fornaciari, E.; Pälike, H.; Rio, D.; Lourens, L.; Hilgen, F. A review of calcareous nannofossil astrobiochronology encompassing the past 25 million years. Quat. Sci. Rev. 2006, 25, 3113–3137. [Google Scholar] [CrossRef]
- Backman, J.; Raffi, I.; Rio, D.; Fornaciari, E.; Pälike, H. Biozonation and biochronology of Miocene through Pleistocene calcareous nannofossils from low and middle latitudes. Newsl. Stratigr. 2012, 47, 131–181. [Google Scholar] [CrossRef]
- Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D.; Ogg, G.M. The Geologic Time Scale 2012; Elsevier: Amsterdam, The Netherlands, 2012; p. 1176. [Google Scholar] [CrossRef]
- Bown, P.R. Taxonomy, evolution, and biostratigraphy of late Triassic-early Jurassic calcareous nannofossils. Spec. Pap. Palaeontol. 1987, 38, 118. [Google Scholar]
- Farrimond, P.; Eglinton, G.; Brassell, S.C. Alkenones in Cretaceous black shales, Blake-Bahama Basin, western North Atlantic. Org. Geochem. 1986, 10, 897–903. [Google Scholar] [CrossRef]
- Brassell, S.C.; Dumitrescu, M.; the ODP Leg 198 Shipboard Scientific Party. Recognition of alkenones in a lower Aptian porcellanite from the west-central Pacific. Org. Geochem. 2004, 35, 181–188. [Google Scholar] [CrossRef]
- Paasche, E. A review of the coccolithophorid Emiliania huxleyi (Prymnesiophyceae), with particular reference to growth, coccolith formation, and calcification-photosynthesis interactions. Phycologia 2002, 40, 503–529. [Google Scholar] [CrossRef]
- Durak, G.M.; Taylor, A.R.; Walker, C.E.; Probert, I.; de Vargas, C.; Audic, S.; Schroeder, D.; Brownlee, C.; Wheeler, G.L. A role for diatom-like silicon transporters in calcifying coccolithophores. Nat. Commun. 2016, 7, 10543. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.E.; Taylor, A.R.; Langer, G.; Durak, G.M.; Heath, S.; Probert, I.; Tyrrell, T.; Brownlee, C.; Wheeler, G.L. The requirement for calcification differs between ecologically important coccolithophore species. New Phytol. 2018, 220, 147–162. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.L.; Lowery, C.M.; Bralower, T.J. Delayed calcareous nannoplankton boom-bust successions in the earliest Paleocene Chicxulub (Mexico) impact crater. Geology 2019, 47, 753–756. [Google Scholar] [CrossRef]
- Kelly, D.C.; Norris, R.D.; Zachos, J. Deciphering the paleoceanographic significance of Early Oligocene Braarudosphaera chalks in the South Atlantic. Mar. Micropaleontol. 2003, 49, 49–63. [Google Scholar] [CrossRef]
- Bown, P.R.; Dunkley Jones, T.; Young, J.R.; Randell, R. A Palaeogene record of extant lower photic zone calcareous nannoplankton. Palaeontology 2009, 52, 457–469. [Google Scholar] [CrossRef]
- Bown, P.R. Paleocene calcareous nannofossils from Tanzania (TDP sites 19, 27 and 38). J. Nannoplankton Res. 2016, 36, 1–32. [Google Scholar]
- Stanley, S.M. Macroevolution, Pattern and Process; Freeman and Company: San Francisco, CA, USA, 1979; p. 332. [Google Scholar]
- Budd, G.E.; Mann, R.P. History is written by the victors: The effect of the push of the past on the fossil record. Evolution 2018, 72, 2276–2291. [Google Scholar] [CrossRef] [Green Version]
- Hannisdal, B.; Henderiks, J.; Liow, L.H. Long-term evolutionary and ecological responses of calcifying phytoplankton to changes in atmospheric CO2. Glob. Chang. Biol. 2012, 18, 3504–3516. [Google Scholar] [CrossRef]
- Šupraha, L.; Henderiks, J. A 15-million-year-long record of phenotypic evolution in the heavily calcified coccolithophore Helicosphaera and its biogeochemical implications. Biogeosciences 2020, 17, 2955–2969. [Google Scholar] [CrossRef]
- Van Valen, L. A New Evolutionary Law. Evol. Theory 1973, 1, 1–30. [Google Scholar]
- Raup, D.M. Taxonomic survivorship curves and Van Valen’s Law. Paleobiology 1975, 1, 82–96. [Google Scholar] [CrossRef]
- Liow, L.H.; Van Valen, L.; Stenseth, N.C. Red Queen: From populations to taxa and communities. Trends Ecol. Evol. 2011, 26, 349–358. [Google Scholar] [CrossRef]
- Rengefors, K.; Kremp, A.; Reusch, T.B.H.; Wood, A.M. Genetic diversity and evolution in eukaryotic phytoplankton: Revelations from population genetic studies. J. Plankton Res. 2017, 39, 165–179. [Google Scholar] [CrossRef] [Green Version]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- De Vargas, C.; Sáez, A.G.; Medlin, L.K.; Thierstein, H.R. Super-species in the calcareous plankton. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 271–298. [Google Scholar]
- Van Bleijswijk, J.D.L.; Kempers, R.S.; Veldhuis, M.J. Cell and growth characteristics of types A and B of Emiliania huxleyi (Prymnesiophyceae) as determined by flow cytometry and chemical analyses. J. Phycol. 1994, 30, 230–241. [Google Scholar] [CrossRef]
- Langer, G.; Nehrke, G.; Probert, I.; Ly, J.; Ziveri, P. Strain-specific responses of Emiliania huxleyi to changing seawater carbonate chemistry. Biogeosciences 2009, 6, 2637–2646. [Google Scholar] [CrossRef] [Green Version]
- Gingerich, P.D. Rates of evolution on the time scale of the evolutionary process. Genetica 2001, 112/113, 127–144. [Google Scholar] [CrossRef] [Green Version]
- Eldredge, N.; Gould, S.J. Punctuated equilibria: An alternative to phyletic gradualism. In Models in Paleobiology; Schopf, T.J.M., Thomas, J.M., Eds.; Freeman, Cooper, & Co.: San Francisco, CA, USA, 1972; pp. 82–115. [Google Scholar]
- Malmgren, B.A.; Berggren, W.A.; Lohmann, G.P. Evidence for punctuated gradualism in the Late Neogene Globorotalia tumida lineage of planktonic foraminifera. Paleobiology 1983, 9, 377–389. [Google Scholar] [CrossRef]
- Hunt, G.; Rabosky, D.L. Phenotypic evolution in fossil species: Pattern and Process. Annu. Rev. Earth Planet. Sci. 2014, 42, 421–441. [Google Scholar] [CrossRef]
- Beaufort, L. Size variations in Late Miocene Reticulofenestra and implication for paleoclimatic interpretation. Mem. Di Sci. Geol. 1992, 43, 339–350. [Google Scholar]
- Imai, R.; Farida, M.; Sato, T.; Iryu, Y. Evidence for eutrophication in the northwestern Pacific and eastern Indian oceans during the Miocene to Pleistocene based on the nannofossil accumulation rate, Discoaster abundance, and coccolith size distribution of Reticulofenestra. Mar. Micropaleontol. 2015, 116, 15–27. [Google Scholar] [CrossRef]
- Matsuoka, H.; Okada, H. Time progressive morphometric changes of the genus Gephyrocapsa in the Quaternary sequence of the tropical Indian Ocean, Site 709. In Proceedings of the Ocean Drilling Program, Scientific Results; Duncan, R.A., Backman, J., Peterson, L.C., Eds.; Ocean Drilling Program: College Station, TX, USA, 1990; Volume 115, pp. 225–270. [Google Scholar]
- Young, J. Size variation of Neogene Reticulofenestra coccoliths from Indian Ocean DSDP Cores. J. Micropaleontol. 1990, 9, 71–86. [Google Scholar] [CrossRef] [Green Version]
- Backman, J.; Hermelin, J.O.R. Morphometry of the Eocene nannofossil Reticulofenestra umbilicus lineage and its biochronological consequences. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1986, 57, 103–116. [Google Scholar] [CrossRef]
- Henderiks, J.; Pagani, M. Coccolithophore cell size and the Paleogene decline in atmospheric CO2. Earth Planet. Sci. Lett. 2008, 269, 575–583. [Google Scholar] [CrossRef]
- Aubry, M.-P. A major Pliocene coccolithophore turnover: Change in morphological strategy in the photic zone. In Large Ecosystem Perturbations: Causes and Consequences; Monechi, S., Coccioni, R., Rampino, M.R., Eds.; The Geological Society of America Special Paper: Boulder, CO, USA, 2007; Volume 424, pp. 25–51. [Google Scholar]
- Beaufort, L.; Bolton, C.T.; Sarr, A.C.; Suchéras-Marx, B.; Rosenthal, Y.; Donnadieu, Y.; Barbarin, N.; Bova, S.; Cornuault, P.; Gally, Y.; et al. Cyclic evolution of phytoplankton forced by changes in tropical seasonality. Nature 2022, 601, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Knappertsbusch, M. Morphologic evolution of the coccolithophorid Calcidiscus leptoporus from the Early Miocene to Recent. J. Paleontol. 2000, 74, 712–730. [Google Scholar] [CrossRef]
- Rivero-Calle, S.; Gnanadesikan, A.; Del Castillo, C.E.; Balch, W.M.; Guikema, S.D. Multidecadal increase in North Atlantic coccolithophores and the potential role of rising CO2. Science 2015, 350, 1533–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, A.; Henderiks, J.; Beaufort, L.; Rickaby, R.E.M.; Brown, C.W. Poleward expansion of the coccolithophore Emiliania huxleyi. J. Plankton Res. 2014, 36, 316–325. [Google Scholar] [CrossRef]
- Bach, L.T.; Riebesell, U.; Gutowska, M.A.; Federwisch, L.; Schulz, K.G. A unifying concept of coccolithophore sensitivity to changing carbonate chemistry embedded in an ecological framework. Prog. Oceanogr. 2015, 135, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Krumhardt, K.M.; Lovenduski, N.S.; Long, M.C.; Levy, M.; Lindsay, K.; Moore, J.K.; Nissen, C. Coccolithophore growth and calcification in an acidified ocean: Insights from community earth system model simulations. J. Adv. Modeling Earth Syst. 2019, 11, 1418–1437. [Google Scholar] [CrossRef] [Green Version]
- Gafar, N.A.; Eyre, B.D.; Schulz, K.G. Particulate inorganic to organic carbon production as a predictor for coccolithophorid sensitivity to ongoing ocean acidification. Limnol. Oceanogr. Lett. 2019, 4, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Finkel, Z.V.; Katz, M.E.; Wright, J.D.; Schofield, O.M.E.; Falkowski, P.G. Climatically driven macroevolutionary patterns in the size of marine diatoms over the Cenozoic. Proc. Natl. Acad. Sci. USA 2005, 102, 8927–8932. [Google Scholar] [CrossRef] [Green Version]
- Westbroek, P.; Marin, F. A marriage of bone and nacre. Nature 1998, 392, 861–862. [Google Scholar] [CrossRef]
- Klaveness, D. The microanatomy of Calyptrosphaera sphaeroidea, with some supplementary observations on the motile stage of Coccolithus pelagicus. Nor. J. Bot. 1973, 20, 151–162. [Google Scholar]
- Rowson, J.D.; Leadbeater, B.S.C.; Green, J.C. Calcium carbonate deposition in the motile (Crystallolithus) phase of Coccolithus pelagicus (Prymnesiophyceae). Br. Phycol. J. 1986, 21, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Young, J.R.; Davis, S.A.; Bown, P.R.; Mann, S. Coccolith ultrastructure and biomineralisation. J. Struct. Biol. 1999, 126, 195–215. [Google Scholar] [CrossRef]
- Langer, G.; Oetjen, K.; Brenneis, T. On culture artefacts in coccolith morphology. Helgol. Mar. Res. 2013, 67, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Gerecht, A.C.; Šupraha, L.; Edvardsen, B.; Probert, I.; Henderiks, J. High temperature decreases the PIC/POC ratio and increases phosphorus requirements in Coccolithus pelagicus (Haptophyta). Biogeosciences 2014, 11, 3531–3545. [Google Scholar] [CrossRef] [Green Version]
- Gerecht, A.C.; Šupraha, L.; Edvardsen, B.; Langer, G.; Henderiks, J. Phosphorus availability modifies carbon production in Coccolithus pelagicus (Wallich) Schiller 1930. J. Exp. Mar. Biol. Ecol. 2015, 472, 24–31. [Google Scholar] [CrossRef]
- Suchéras-Marx, B.; Guihou, A.; Giraud, F.; Lecuyer, C.; Allemand, P.; Pittet, B.; Mattioli, E. Impact of the Middle Jurassic diversification of Watznaueria (coccolith-bearing algae) on the carbon cycle and d13C of bulk marine carbonates. Glob. Planet. Chang. 2012, 86–87, 92–100. [Google Scholar] [CrossRef]
- Young, J. Functions of coccoliths. In Coccolithophores; Winter, A., Siesser, W.G., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 63–82. [Google Scholar]
- Jaya, B.N.; Hoffman, R.; Kirchlechner, C.; Dehm, G.; Scheu, G.; Langer, G. Coccospheres confer mechanical protection: New evidence for an old hypothesis. Acta Biomater. 2016, 42, 258–264. [Google Scholar] [CrossRef]
- Quintero-Torres, R.; Arago, J.L.; Torres, M.; Estrada, M.; Cros, L. Strong far-field coherent scattering of ultraviolet radiation by holococcolithophores. Phys. Rev. E 2006, 74, e032901. [Google Scholar] [CrossRef] [Green Version]
- Henderiks, J. Coccolithophore size rules—Reconstructing ancient cell geometry and cellular calcite quota from fossil coccoliths. Mar. Micropaleontol. 2008, 67, 143–154. [Google Scholar] [CrossRef]
- Gibbs, S.; Poulton, A.J.; Bown, P.R.; Daniels, C.J.; Hopkins, J.; Young, J.R.; Jones, H.L.; Thiemann, G.J.; O’Dea, S.A.; Newsam, C. Species-specific growth response of coccolithophores to Palaeocene-Eocene environmental change. Nat. Geosci. 2013, 6, 218–222. [Google Scholar] [CrossRef]
- Sheward, R.M.; Poulton, A.J.; Gibbs, S.J.; Daniels, C.J.; Bown, P.R. Physiology regulates the relationship between coccosphere geometry and growth phase in coccolithophores. Biogeosciences 2017, 14, 1493–1509. [Google Scholar] [CrossRef] [Green Version]
- Suchéras-Marx, B.; Viseur, S.; Walker, C.E.; Beaufort, L.; Bolton, C. Coccolith size rules—What controls the size of coccoliths during coccolithogenesis? Mar. Micropaleontol. 2021, 170, 102080. [Google Scholar] [CrossRef]
- Young, J.; Ziveri, P. Calculation of coccolith volume and its use in calibration of carbonate flux estimates. Deep.-Sea Res. II 2000, 47, 1679–1700. [Google Scholar] [CrossRef]
- Beaufort, L. Weight estimates of coccoliths using the optical properties (birefringence) of calcite. Micropaleontology 2005, 51, 289–298. [Google Scholar] [CrossRef]
- Beaufort, L.; Barbarin, N.; Gally, Y. Optical measurements to determine the thickness of calcite crystals and the mass of thin carbonate particles such as coccoliths. Nat. Protoc. 2014, 9, 633–642. [Google Scholar] [CrossRef]
- McClelland, H.L.O.; Barbarin, N.; Beaufort, L.; Hermoso, M.; Ferretti, P.; Greaves, M.; Rickaby, R.E.M. Calcification response of a key phytoplankton family to millennial-scale environmental change. Sci. Rep. 2016, 6, 34263. [Google Scholar] [CrossRef] [Green Version]
- Hermoso, M.; Lefeuvre, B.; Minoletti, F.; de Rafélis, M. Extreme strontium concentrations reveal specific biomineralization pathways in certain coccolithophores with implications for the Sr/Ca paleoproductivity proxy. PLoS ONE 2017, 12, e0185655. [Google Scholar] [CrossRef] [Green Version]
- Meyer, E.M.; Langer, G.; Brownlee, C.; Wheeler, G.L.; Taylor, A.R. Sr in coccoliths of Scyphosphaera apsteinii: Partitioning behavior and role in coccolith morphogenesis. Geochim. Cosmochim. Acta 2020, 285, 41–54. [Google Scholar] [CrossRef]
- Suchéras-Marx, B.; Giraud, F.; Simionovici, A.; Tucoulou, R.; Daniel, I. Evidence of high Sr/Ca in a Middle Jurassic murolith coccolith species. Peer Community J. 2021, 1, e25. [Google Scholar] [CrossRef]
- Unrein, F.; Gasol, J.M.; Not, F.; Forn, I.; Massana, R. Mixotrophic haptophytes are key bacterial grazers in oligotrophic coastal waters. ISME J. 2014, 8, 164–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avrahami, Y.; Frada, M.J. Detection of phagotrophy in the marine phytoplankton group of the coccolithophores (Calcihaptophycidae, Haptophyta) during nutrient-replete and phosphate-limited growth. J. Phycol. 2020, 56, 1103–1108. [Google Scholar] [CrossRef]
- Kawachi, M.; Inouye, I.; Maeda, O.; Chihara, M. The haptonema as a food-capturing device: Observations on Chrysochromulina hirta (Prymnesiophyceae). Phycologia 1991, 30, 563–573. [Google Scholar] [CrossRef]
- Pintner, I.J.; Provasoli, L. Heterotrophy in subdued light of 3 Chrysochromulina species. Proc. US-Jpn. Semin. Mar. Microbiol. 1968, 12, 25–31. [Google Scholar]
- Rokitta, S.D.; de Nooijer, L.J.; Trimborn, S.; de Vargas, C.; Rost, B.; John, U. Transcriptome analyses reveal differential gene expression patterns between the life-cycle stages of Emiliania huxleyi (Haptophyta) and reflect specialization to different ecological niches. J. Phycol. 2011, 47, 829–838. [Google Scholar] [CrossRef]
- Houdan, A.; Probert, I.; Zatylny, C.; Véron, B.; Billard, C. Ecology of oceanic coccolithophores. I. Nutritional preferences of the two stages in the life cycle of Coccolithus braarudii and Calcidiscus leptoporus. Aquat. Microb. Ecol. 2006, 44, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Godrijan, J.; Drapeau, D.T.; Balch, W.M. Mixotrophic uptake of organic compounds by coccolithophores. Limnol. Oceanogr. 2020, 65, 1410–1421. [Google Scholar] [CrossRef]
- Langer, G.; Wen Jie, V.; Kottmeier, D.; Flori, S.; Sturm, D.; de Vries, J.; Harper, G.M.; Brownlee, C.; Wheeler, G. Distinct physiological responses of Coccolithus braarudii life cycle phases to light intensity and nutrient availability. Eur. J. Phycol. 2022, 1–14. [Google Scholar] [CrossRef]
- Okada, H.; Honjo, S. The distribution of oceanic coccolithophorids in the Pacific. Deep.-Sea Res. 1973, 20, 355–374. [Google Scholar] [CrossRef]
- Marchant, H.J.; Thomsen, H.A. Haptophytes in polar waters. In The Haptophyte Algae; Green, J.C., Leadbeater, B.S.C., Eds.; Clarendon Press: Oxford, UK, 1994; Volume 51, pp. 209–228. [Google Scholar]
- Hagino, K.; Okada, H. Gladiolithus striatus sp. nov. (Prymnesiophyceae), a living coccolithophore from the lower photic zone of the Pacific Ocean. Phycologia 1998, 37, 246–250. [Google Scholar] [CrossRef]
- Van Lenning, K.; Probert, I.; Latasa, M.; Estrada, M.; Young, J.R. Pigment diversity of coccolithophores in relation to taxonomy, phylogeny and ecological preferences. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 51–73. [Google Scholar]
- Von Dassow, P.; Montresor, M. Unveiling the mysteries of phytoplankton life cycles: Patterns and opportunities behind complexity. J. Plankton Res. 2011, 33, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Valero, M.; Richerd, S.; Perrot, V.; Destombe, C. Evolution of alternation of haploid and diploid phases in life cycles. TREE 1992, 7, 25–29. [Google Scholar] [CrossRef]
- Noël, M.-H.; Kawachi, M.; Inouye, I. Induced dimorphic life cycle of a coccolithophorid, Calyptrosphaera sphaeroidea (Prymnesiophyceae, Haptophyta). J. Phycol. 2004, 40, 112–129. [Google Scholar] [CrossRef]
- Parke, M.; Adams, I. The motile (Crystallolithus hyalinus Gaarder & Markali) and non-motile phases in the life history of Coccolithus pelagicus (Wallich) Schiller. J. Mar. Biol. Ass. UK 1960, 39, 263–274. [Google Scholar]
- Rayns, D.G. Alternation of generations in a coccolithophorid, Cricosphaera carterae (Braarud & Fagerl.) Braarud. J. Mar. Biol. Ass. U.K. 1962, 42, 481–484. [Google Scholar]
- Leadbeater, B.S.C. Preliminary observations on differences of scale morphology at various stages in the life cycle of ‘Apistonema-Syracosphaera’ sensu von Stosch. Br. Phycol. J. 1970, 5, 57–69. [Google Scholar] [CrossRef]
- Jordan, R.W.; Kleijne, A. A. A classification system for living coccolithophores. In Coccolithophores; Winter, A., Siesser, W.G., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 83–105. [Google Scholar]
- Paasche, E.; Klaveness, D. A physiological comparison of coccolith-forming and naked cells of Coccolithus huxleyi. Arch. Mikrobiol. 1970, 73, 143–152. [Google Scholar] [CrossRef]
- Green, J.C.; Course, P.A.; Tarran, G.A. The life-cycle of Emiliania huxleyi: A brief review and a study of the relative ploidy levels analysed by flow cytometry. J. Mar. Syst. 1996, 9, 33–44. [Google Scholar] [CrossRef]
- Cros, L.; Estrada, M. Holo-heterococcolithophore life cycles: Ecological implications. Mar. Ecol. Prog. Ser. 2013, 492, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Dimiza, M.D.; Triantaphyllou, M.V.; Dermitzakis, M.D. Vertical distribution and ecology of living coccolithophores in the marine ecosystems of Andros Island (Middle Aegean Sea) during late summer 2001. Hell. J. Geosci. 2008, 43, 7–20. [Google Scholar]
- Oviedo, A.M.; Ziveri, P.; Álvarez, M.; Tanhua, T. Is coccolithophore distribution in the Mediterranean Sea related to seawater carbonate chemistry? Ocean Sci. 2015, 11, 13–32. [Google Scholar] [CrossRef] [Green Version]
- Šupraha, L.; Ljubešić, Z.; Mihanović, H.; Henderiks, J. Coccolithophore life-cycle dynamics in a coastal Mediterranean ecosystem: Seasonality and species-specific patterns. J. Plankton Res. 2016, 38, 1178–1193. [Google Scholar] [CrossRef] [Green Version]
- Tyrrell, T.; Merico, A. Emiliania huxleyi: Bloom observations and the conditions that induce them. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 75–97. [Google Scholar]
- Henderiks, J.; Winter, A.; Elbrächter, M.; Feistel, R.; van der Plas, A.K.; Nausch, G.; Barlow, R. Environmental controls on Emiliania huxleyi morphotypes in the Benguela coastal upwelling system (SE Atlantic). Mar. Ecol. Prog. Ser. 2012, 448, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Frada, M.J.; Bidle, K.D.; Probert, I.; de Vargas, C. In situ survey of life cycle phases of the coccolithophore Emiliania huxleyi (Haptophyta). Environ. Microbiol. 2012, 14, 1558–1569. [Google Scholar] [CrossRef]
- Keeling, P.J.; Burki, F.; Wilcox, H.W.; Allam, B.; Allen, E.E.; Amaral-Zettler, L.A.; Armbrust, E.A.; Archibald, J.M.; Bharti, A.K.; Bell, C.J.; et al. The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): Illuminating the functional diversity of eukaryotic life in the oceans through transcriptome sequencing. PLoS Biol. 2014, 12, e1001889. [Google Scholar] [CrossRef] [Green Version]
- Von Dassow, P.; Ogata, H.; Probert, I.; Wincker, P.; Da Silva, C.; Audic, S.; Claverie, J.-M.; de Vargas, C. Transcriptome analysis of functional differentiation between haploid and diploid cells of Emiliania huxleyi, a globally significant photosynthetic calcifying cell. Genome Biol. 2009, 10, R114. [Google Scholar] [CrossRef]
- Mackinder, L.; Wheeler, G.; Schroeder, D.; Riebesell, U.; Brownlee, C. Molecular mechanisms underlying calcification in coccolithophores. Geomicrobiol. J. 2010, 27, 585–595. [Google Scholar] [CrossRef]
- Von Dassow, P.; John, U.; Ogata, H.; Probert, I.; Bendif, E.M.; Kegel, J.U.; Audic, S.; Wincker, P.; Da Silva, C.; Claverie, J.-M.; et al. Life-cycle modification in open oceans accounts for genome variability in a cosmopolitan phytoplankton. ISME J. 2015, 9, 1365–1377. [Google Scholar] [CrossRef] [Green Version]
- Frada, M.J.; Rosenwasser, S.; Ben-Dor, S.; Shemi, A.; Sabanay, H.; Vardi, A. Morphological switch to a resistant subpopulation in response to viral infection in the bloom-forming coccolithophore Emiliania huxleyi. PLoS Pathog. 2017, 13, e1006775. [Google Scholar] [CrossRef] [Green Version]
- Godrijan, J.; Young, J.R.; Marić Pfannkuchen, D.; Precali, R.; Pfannkuchen, M. Coastal zones as important habitats of coccolithophores: A study of species diversity, succession, and life-cycle phases. Limnol. Oceanogr. 2018, 63, 1692–1710. [Google Scholar] [CrossRef]
- Frada, M.; Probert, I.; Allen, M.J.; Wilson, W.H.; de Vargas, C. The “Cheshire Cat” escape strategy of the coccolithophore Emiliania huxleyi in response to viral infection. Proc. Natl. Acad. Sci. USA 2008, 105, 15944–15949. [Google Scholar] [CrossRef] [Green Version]
- Krueger-Hadfield, S.A.; Balestreri, C.; Schroeder, J.; Highfield, A.; Helaouët, P.; Allum, J.; Moate, R.; Lohbeck, K.T.; Miller, P.I.; Riebesell, U.; et al. Genotyping an Emiliania huxleyi (Prymnesiophyceae) bloom event in the North Sea reveals evidence of asexual reproduction. Biogeosciences 2014, 11, 5215–5234. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.S.; Jones, R.C.; Vaillancourt, R.E.; Hallegraeff, G.M. Genetic differentiation among Australian and Southern Ocean populations of the ubiquitous coccolithophore Emiliania huxleyi (Haptophyta). Phycologia 2013, 52, 368–374. [Google Scholar] [CrossRef]
- Otto, S.P. The evolutionary enigma of sex. Am. Nat. 2009, 174, S1–S14. [Google Scholar] [CrossRef] [Green Version]
- Westbroek, P.; Brown, C.W.; van Bleijswijk, J.; Brownlee, C.; Brummer, G.-J.; Conte, M.; Egge, J.; Fernández, E.; Jordan, R.; Knappertsbusch, M.; et al. A model system approach to biological climate forcing. The example of Emiliania huxleyi. Glob. Planet. Chang. 1993, 8, 27–46. [Google Scholar] [CrossRef]
- Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R.; Young, J.R. (Eds.) Springer: Berlin/Heidelberg, Germany, 2004; p. 565. [Google Scholar]
- Mock, T. Silicon drives the evolution of complex crystal morphology in calcifying algae. New Phytol. 2021, 231, 1663–1666. [Google Scholar] [CrossRef]
- Probert, I.; Houdan, A. The laboratory culture of coccolithophores. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 217–249. [Google Scholar]








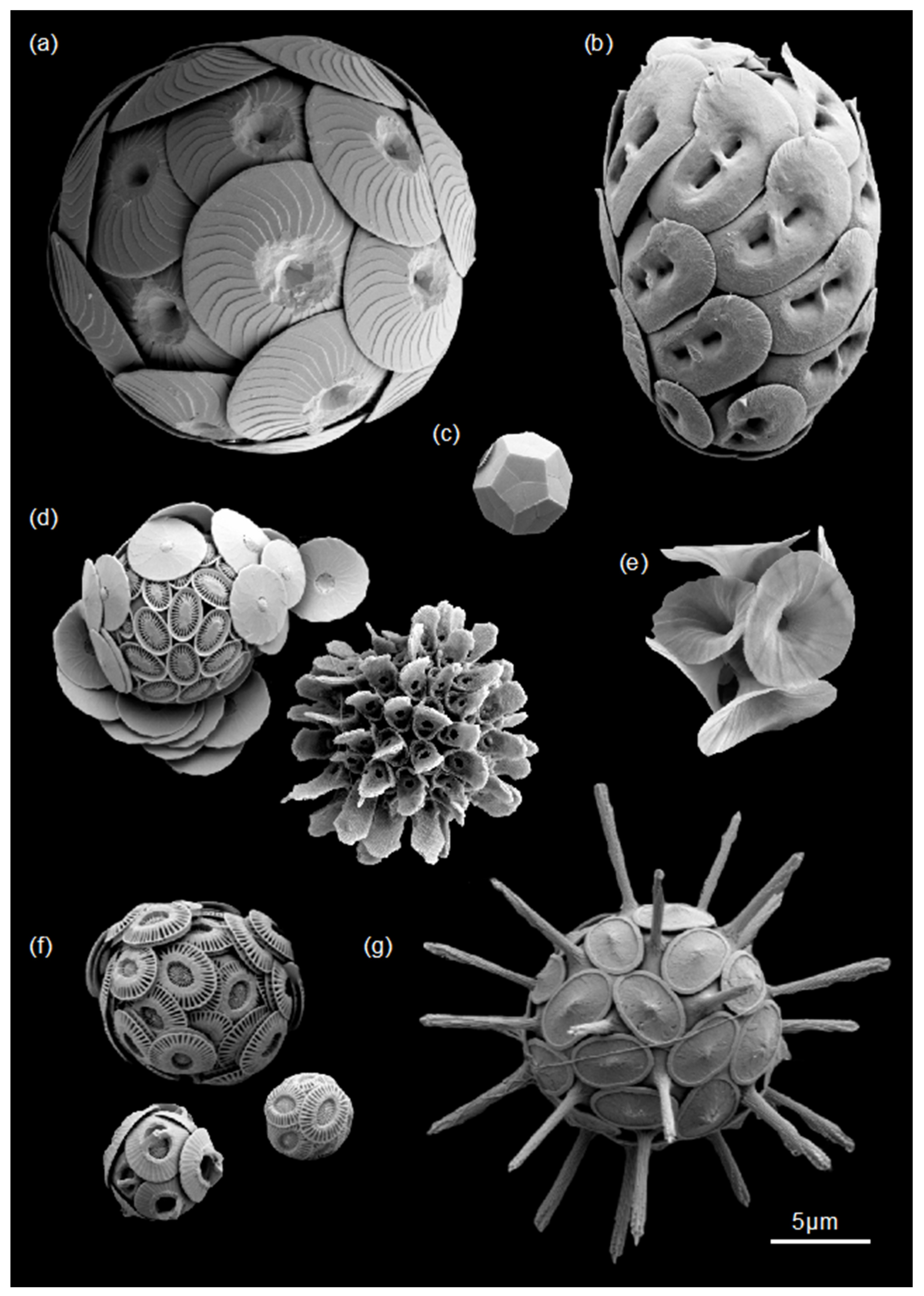{kind=link}
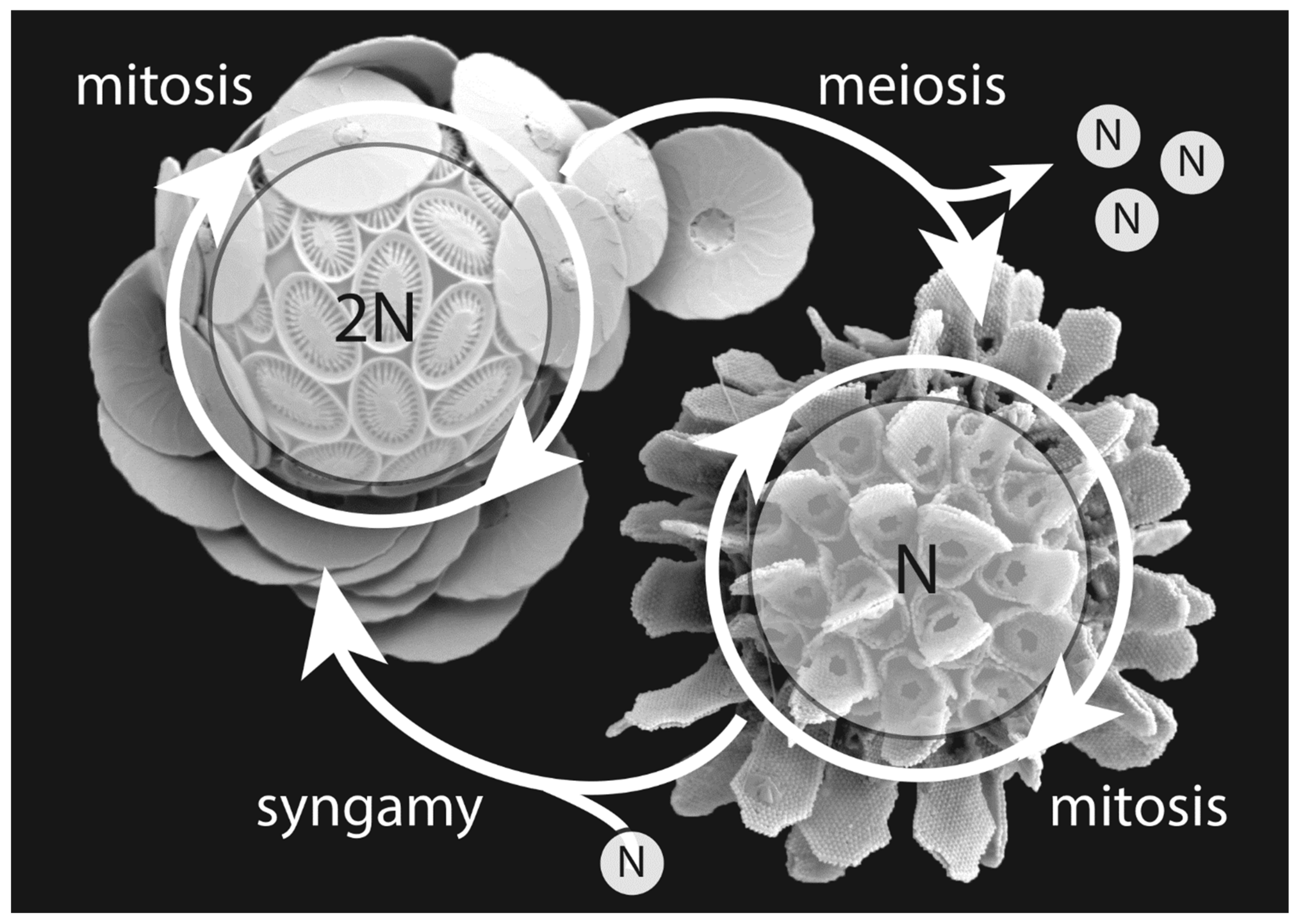{kind=link}
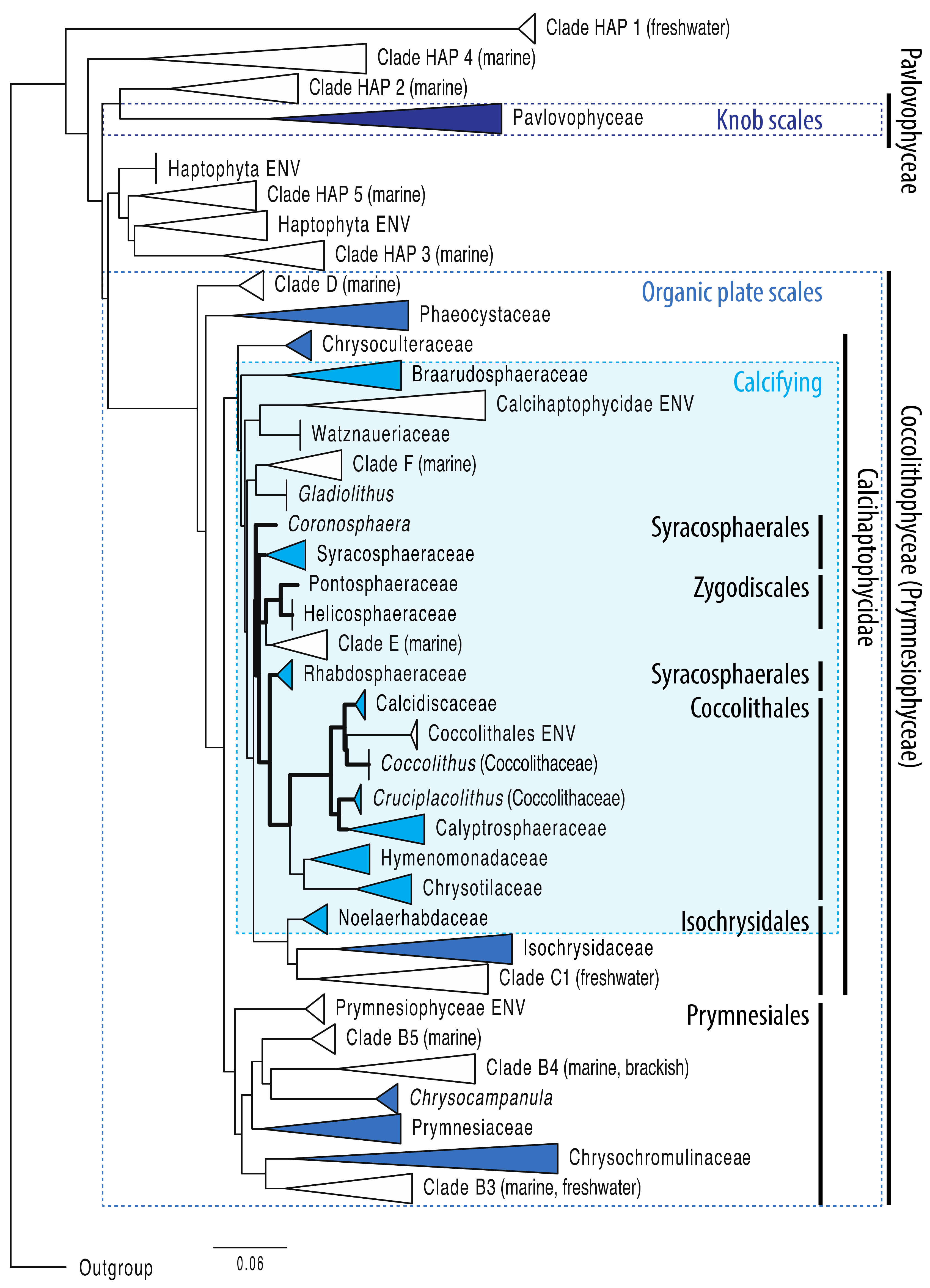{kind=link}
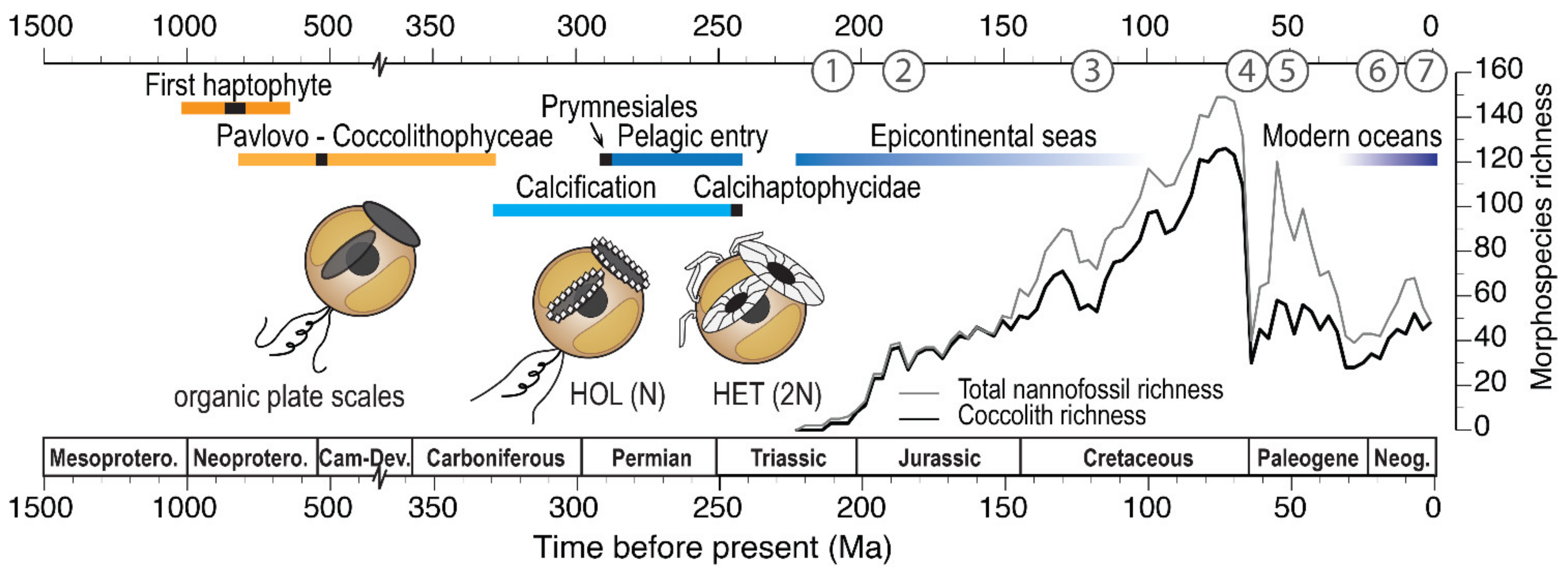{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
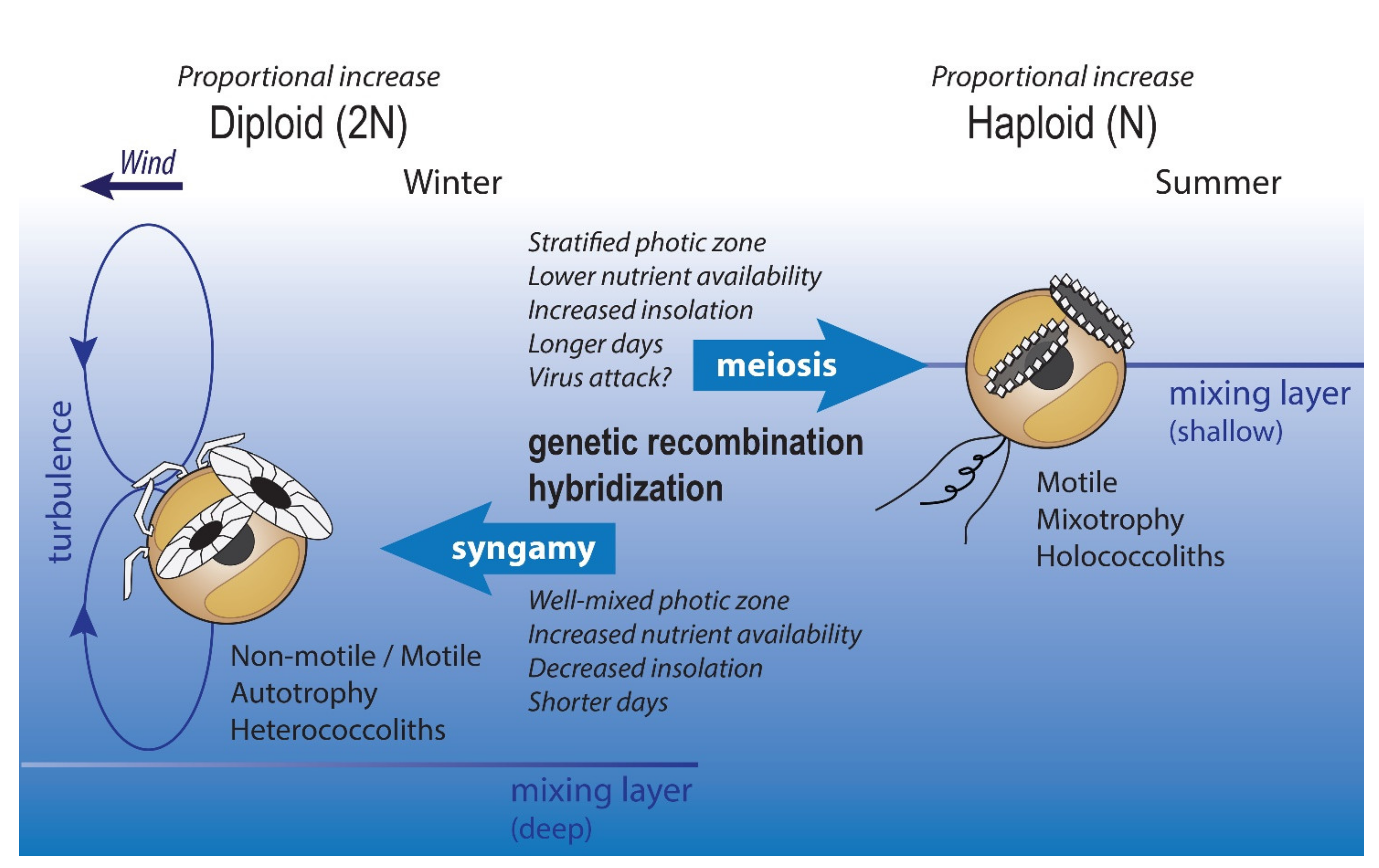{kind=link}
| Coccolithophore Family | Input Data | Longevity (Myr) | Rate Type | Arithmetic Rate Myr−1 | Exponential Rate Myr−1 | |
|---|---|---|---|---|---|---|
| Braarudosphaeraceae | FO (Ma) | 125 | 16.82 (aver) | S | 0.049 | 0.059 |
| Extinct | 32 | 100.5 (max) | E | 0.054 | 0.059 | |
| Extant | 1 | 0.57 (min) | D | −0.005 | 0.000 | |
| Syracosphaeraceae 1 | FO (Ma) | 59 | ? | S | ? | ? |
| Extinct | (11) | 58.8? | E | ? | ? | |
| Extant | 55 | <2? | D | ? | 0.068 | |
| Helicosphaeraceae | FO (Ma) | 52 | 10.39 | S | 0.112 | 0.123 |
| Extinct | 44 | 33.27 | E | 0.080 | 0.096 | |
| Extant | 4 | 0.68 | D | 0.032 | 0.027 | |
| Calcidiscaceae | FO (Ma) | 62 | 14.4 | S | 0.088 | 0.105 |
| Extinct | 26 | 59.5 | E | 0.049 | 0.069 | |
| Extant | 9 | 1.55 | D | 0.039 | 0.035 | |
| Coccolithaceae | FO (Ma) | 66 | 13.33 | S | 0.073 | 0.092 |
| Extinct | 85 | 66 | E | 0.069 | 0.075 | |
| Extant | 3 | 0.5 | D | 0.004 | 0.017 | |
| Noelaerhabdaceae | FO (Ma) | 57 | 9.49 | S | 0.123 | 0.142 |
| Extinct | 51 | 47.91 | E | 0.083 | 0.105 | |
| Extant | 8 | 0.22 | D | 0.041 | 0.036 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henderiks, J.; Sturm, D.; Šupraha, L.; Langer, G. Evolutionary Rates in the Haptophyta: Exploring Molecular and Phenotypic Diversity. J. Mar. Sci. Eng. 2022, 10, 798. https://doi.org/10.3390/jmse10060798
Henderiks J, Sturm D, Šupraha L, Langer G. Evolutionary Rates in the Haptophyta: Exploring Molecular and Phenotypic Diversity. Journal of Marine Science and Engineering. 2022; 10(6):798. https://doi.org/10.3390/jmse10060798
Chicago/Turabian StyleHenderiks, Jorijntje, Daniela Sturm, Luka Šupraha, and Gerald Langer. 2022. "Evolutionary Rates in the Haptophyta: Exploring Molecular and Phenotypic Diversity" Journal of Marine Science and Engineering 10, no. 6: 798. https://doi.org/10.3390/jmse10060798