
Hydrolysis of Methylumbeliferyl Substrate Proxies for Esterase Activities as Indicator for Microbial Oil Degradation in the Ocean: Evidence from Observations in the Aftermath of the Deepwater Horizon Oil Spill (Gulf of Mexico)


Abstract
:1. Background
2. Esterase Activities in Oil-Contaminated Water under Controlled Laboratory Conditions
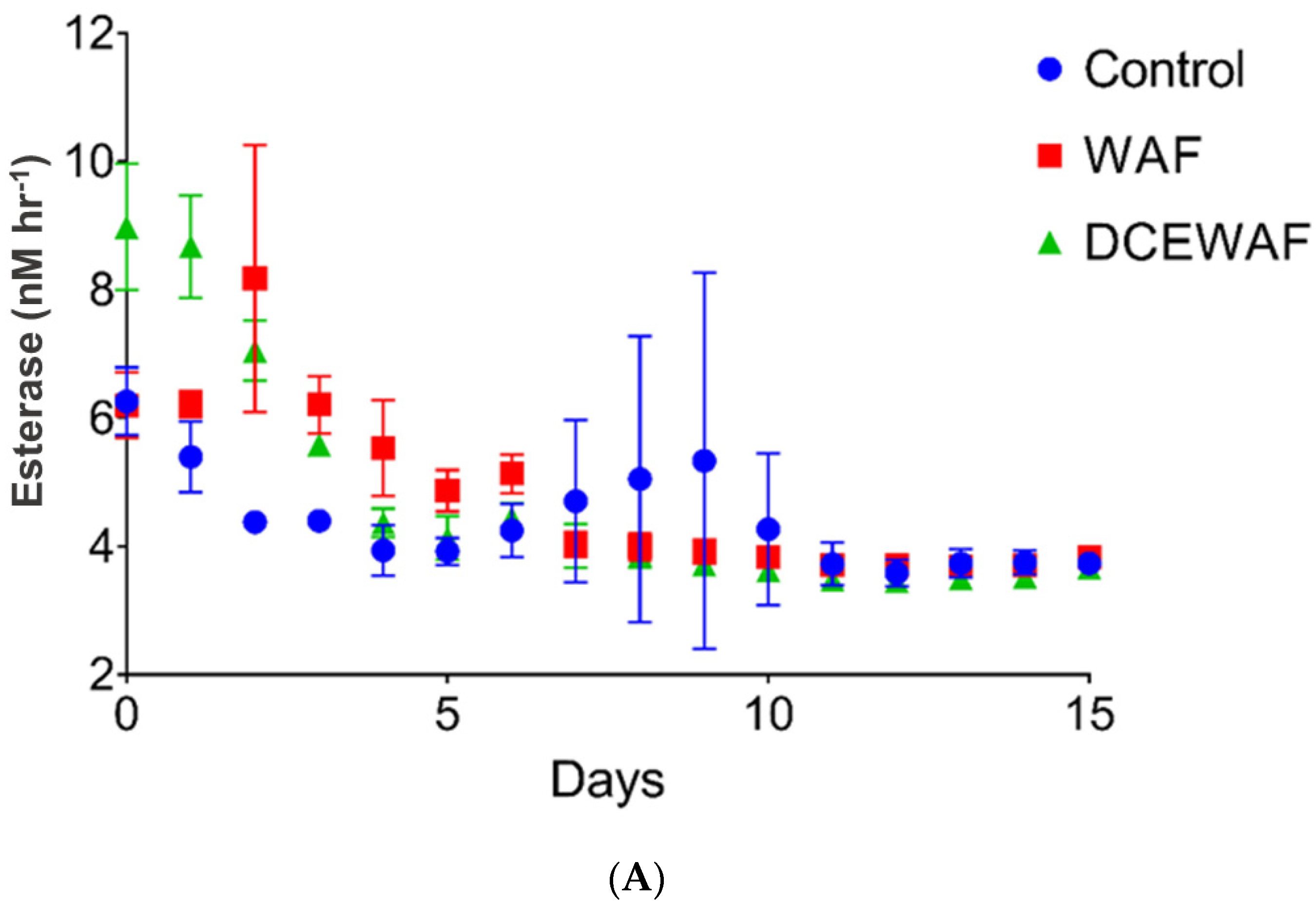
2.1. Case Study 1: Surface Water Incubation
2.2. Case Study 2: Deep-Water Incubation
3. Esterase Activities as Indicator for Oil Biodegradation
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Overton, E.; Wade, T.; Radovic, J.; Meyer, B.; Miles, M.S.; Larter, S. Chemical Composition of Macondo and Other Crude Oils and Compositional Alterations During Oil Spills. Oceanography 2016, 29, 50–63. [Google Scholar] [CrossRef] [Green Version]
- Passow, U.; Overton, E.B. The Complexity of Spills: The Fate of the Deepwater Horizon Oil. Annu. Rev. Mar. Sci. 2021, 13, 109–136. [Google Scholar] [CrossRef]
- Atlas, R.M.; Hazen, T.C. Oil Biodegradation and Bioremediation: A Tale of the Two Worst Spills in US History. Environ. Sci. Technol. 2011, 45, 6709–6715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diercks, A.R.; Highsmith, R.C.; Asper, V.L.; Joung, D.; Zhou, Z.; Guo, L.; Shiller, A.M.; Joye, S.B.; Teske, A.P.; Guinasso, N.; et al. Characterization of Subsurface Polycyclic Aromatic Hydrocarbons at the Deepwater Horizon Site. Geophys. Res. Lett. 2010, 37, L20602. [Google Scholar] [CrossRef] [Green Version]
- Reddy, C.M.; Arey, J.S.; Seewald, J.S.; Sylva, S.P.; Lemkau, K.L.; Nelson, R.K.; Carmichael, C.A.; McIntyre, C.P.; Fenwick, J.; Ventura, G.T.; et al. Composition and Fate of Gas and Oil Released to the Water Column during the Deepwater Horizon Oil Spill. Proc. Natl. Acad. Sci. USA 2011, 109, 20229–20234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kujawinski, L.; Sole, M.C.K.; Valentine, D.L.; Boysen, A.K.; Longnecker, K.; Redmond, M.C. Fate of Dispersants Associated with the Deepwater Horizon Oil Spill. Environ. Sci. Technol. 2011, 45, 1298–1306. [Google Scholar] [CrossRef]
- Crespo-Medina, M.; Meile, C.D.; Hunter, K.S.; Diercks, A.-R.; Asper, V.L.; Orphan, V.J.; Tavormina, P.L.; Nigro, L.M.; Battles, J.J.; Chanton, J.P.; et al. The Rise and Fall of Methanotrophy Following a Deepwater Oil-Well Blowout. Nat. Geosci. 2014, 7, 423–427. [Google Scholar] [CrossRef] [Green Version]
- Valentine, D.L.; Kessler, J.D.; Redmond, M.C.; Mendes, S.D.; Heintz, M.B.; Farwell, C.; Hu, L.; Kinnaman, F.S.; Yvon-Lewis, S.; Du, M.; et al. Propane Respiration Jump-Starts Microbial Response to a Deep Oil Spill. Science 2010, 330, 208–211. [Google Scholar] [CrossRef] [Green Version]
- Hazen, T.C.; Dubinsky, E.A.; DeSantis, T.Z.; Andersen, G.L.; Piceno, Y.M.; Singh, N.; Jansson, J.K.; Probst, A.; Borglin, S.E.; Fortney, J.L.; et al. Deep-Sea Oil Plume Enriches Indigenous Oil-Degrading Bacteria. Science 2010, 330, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Edwards, B.R.; Reddy, C.M.; Camilli, R.; Carmichael, C.A.; Longnecker, K.; Mooy, B.A.S.V. Rapid Microbial Respiration of Oil from the Deepwater Horizon Spill in Offshore Surface Waters of the Gulf of Mexico. Environ. Res. Lett. 2011, 6, 035301. [Google Scholar] [CrossRef]
- Shiller, A.M.; Joung, D. Nutrient Depletion as a Proxy for Microbial Growth in Deepwater Horizon Subsurface Oil/Gas Plumes. Environ. Res. Lett. 2012, 7, 045301. [Google Scholar] [CrossRef] [Green Version]
- Head, I.M.; Jones, D.M.; Röling, W.F.M. Marine Microorganisms Make a Meal of Oil. Nat. Rev. Microbiol. 2006, 4, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Joye, S.B.; Kleindienst, S.; Gilbert, J.A.; Handley, K.M.; Weisenhorn, P.; Overholt, W.A.; Kostka, J.E. Responses of Microbial Communities to Hydrocarbon Exposures. Oceanography 2016, 29, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Joye, S.B.; Kostka, J.E.; Halanych, K.M.; Colwell, R.R. GoMRI Insights into Microbial Genomics and Hydrocarbon Bioremediation Response in Marine Ecosystems. Oceanography 2021, 24, 124–135. [Google Scholar]
- Rojo, F. Degradation of Alkanes by Bacteria. Environ. Microbiol. 2009, 11, 2477–2490. [Google Scholar] [CrossRef]
- Ehrhardt, M.G.; Burns, K.A.; Bicego, M.C. Sunlight-Induced Compositional Alterations in the Seawater-Soluble Fraction of a Crude Oil. Mar. Chem. 1992, 37, 53–64. [Google Scholar] [CrossRef]
- Ji, H.; Gong, Y.; Duan, J.; Zhao, D.; Liu, W. Degradation of Petroleum Hydrocarbons in Seawater by Simulated Surface-Level Atmospheric Ozone: Reaction Kinetics and Effect of Oil Dispersant. Mar. Pollut. Bull. 2018, 135, 427–440. [Google Scholar] [CrossRef]
- Sun, L.; Chiu, M.-H.; Xu, C.; Lin, P.; Schwehr, K.A.; Bacosa, H.; Kamalanathan, M.; Quigg, A.; Chin, W.-C.; Santschi, P.H. The Effects of Sunlight on the Composition of Exopolymeric Substances and Subsequent Aggregate Formation during Oil Spills. Mar. Chem. 2018, 203, 49–54. [Google Scholar] [CrossRef]
- Ben Ali, W.; Chaduli, D.; Navarro, D.; Lechat, C.; Turbé-Doan, A.; Bertrand, E.; Faulds, C.B.; Sciara, G.; Lesage-Meessen, L.; Record, E.; et al. Screening of Five Marine-Derived Fungal Strains for Their Potential to Produce Oxidases with Laccase Activities Suitable for Biotechnological Applications. BMC Biotechnol. 2020, 20, 27. [Google Scholar] [CrossRef]
- Vasconcelos, M.R.D.S.; Vieira, G.A.L.; Otero, I.V.R.; Bonugli-Santos, R.C.; Rodrigues, M.V.N.; Rehder, V.L.G.; Ferro, M.; Boaventura, S.; Bacci, M.; Sette, L.D. Pyrene Degradation by Marine-Derived Ascomycete: Process Optimization, Toxicity, and Metabolic Analyses. Environ. Sci. Pollut. Res. 2019, 26, 12412–12424. [Google Scholar] [CrossRef]
- Yusoff, D.F.; Raja Abd Rahman, R.N.; Masomian, M.; Ali, M.S.; Leow, T.C. Newly Isolated Alkane Hydroxylase and Lipase Producing Geobacillus and Anoxybacillus Species Involved in Crude Oil Degradation. Catalysts 2020, 10, 851. [Google Scholar] [CrossRef]
- Breuil, C.; Shindler, D.B.; Sijher, J.S.; Kushner, D.J. Stimulation of Lipase Production During Bacterial Growth on Alkanes. J. Bacteriol. 1978, 133, 601–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margesin, R.; Zimmerbauer, A.; Schinner, F. Soil Lipase Activity–A Useful Indicator of Oil Biodegradation. Biotechnol. Tech. 1999, 13, 859–863. [Google Scholar] [CrossRef]
- Glogauer, A.; Martini, V.P.; Faoro, H.; Couto, G.H.; Müller-Santos, M.; Monteiro, R.A.; Mitchell, D.A.; de Souza, E.M.; Pedrosa, F.O.; Krieger, N. Identification and Characterization of a New True Lipase Isolated through Metagenomic Approach. Microb. Cell Factories 2011, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, H.-G. Significance of Exoenzymatic Activities in the Ecology of Brackish Water: Measurements by Means of Methylumbelliferyl-Substrates. Mar. Ecol. Prog. Ser. 1983, 11, 299–308. [Google Scholar] [CrossRef]
- Martinez, J.; Azam, F. Periplasmic Aminopeptidase and Alkaline-Phosphatase Activities in a Marine Bacterium-mplications for Substrate Processing in the Sea. Mar. Ecol. Prog. Ser. 1993, 92, 89–97. [Google Scholar] [CrossRef]
- Chrost, R.J. Microbial Enzymes in Aquatic Environments; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Cunha, Â.; Almeida, A.; Gomes, N.C.M.; Oliveira, V.; Santos, A.L. Bacterial Extracellular Enzymatic Activity in Globally Changing Aquatic Ecosystems. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 1, 124–135. [Google Scholar]
- Hoppe, H.-G. Phosphatase Activity in the Sea. Hydrobiologia 2003, 493, 187–200. [Google Scholar] [CrossRef]
- Hoppe, H.-G.; Arnosti, C.; Herndl, G.J. Ecological Significance of Bacterial Enzymes in the Marine Environment. In Enzymes in the Environment; Burns, R.G., Dick, R.P., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 73–107. [Google Scholar]
- Rullkötter, J. Organic Matter: The Driving Force for Early Diagenesis; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Boetius, A. Microbial Hydrolytic Enzyme Activities in Deep-Sea Sediments. Helgol. Meeresunters. 1995, 49, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Boetius, A.; Springer, B.; Petry, C. Microbial Activity and Particulate Matter in the Benthic Nepheloid Layer (BNL) of the Deep Arabian Sea. Deep-Sea Res. Part II-Top. Stud. Oceanogr. 2000, 47, 2687–2706. [Google Scholar] [CrossRef]
- Boetius, A.; Scheibe, S.; Tselepides, A.; Thiel, H. Microbial Biomass and Activities in Deep-Sea Sediments of the Eastern Mediterranean: Trenches Are Benthic Hotspots. DEEP-SEA Res. PART-Oceanogr. Res. Pap. 1996, 43, 1439–1460. [Google Scholar] [CrossRef]
- Boetius, A.; Lochte, K. Effect of Organic Enrichments on Hydrolytic Potentials and Growth of Bacteria in Deep-Sea Sediments. Mar. Ecol. Prog. Ser. 1996, 140, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Boetius, A.; Lochte, K. Regulation of Microbial Enzymatic Degradation of Organic Matter in Deep-Sea Sediments. Mar. Ecol. Prog. Ser. 1994, 104, 299–307. [Google Scholar] [CrossRef]
- King, G.M. Characterization of β-Glucosidase Activity in Intertidal Marine Sediments. Appl. Environ. Microbiol. 1986, 51, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Ziervogel, K.; McKay, L.; Rhodes, B.; Osburn, C.L.; Dickson-Brown, J.; Arnosti, C.; Teske, A. Microbial Activities and Dissolved Organic Matter Dynamics in Oil-Contaminated Surface Seawater from the Deepwater Horizon Oil Spill Site. PLoS ONE 2012, 7, e34816. [Google Scholar] [CrossRef]
- Ziervogel, K.; Joye, S.B.; Arnosti, C. Microbial Enzymatic Activity and Secondary Production in Sediments Affected by the Sedimentation Pulse Following the Deepwater Horizon Oil Spill. Deep Sea Res. Part II Top. Stud. Oceanogr. 2016, 129, 241–248. [Google Scholar] [CrossRef]
- Ziervogel, K.; Sweet, J.; Juhl, A.R.; Passow, U. Sediment Resuspension and Associated Extracellular Enzyme Activities Measured Ex Situ: A Mechanism for Benthic-Pelagic Coupling in the Deep Gulf of Mexico. Front. Mar. Sci. 2021, 8, 1654. [Google Scholar] [CrossRef]
- Shi, D.; Bera, G.; Knap, A.H.; Quigg, A.; Al Atwah, I.; Gold-Bouchot, G.; Wade, T.L. A Mesocosm Experiment to Determine Half-Lives of Individual Hydrocarbons in Simulated Oil Spill Scenarios with and without the Dispersant, Corexit. Mar. Pollut. Bull. 2020, 151, 110804. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, S.; Beaver, M.; Lin, P.; Sun, L.; Doyle, S.M.; Sylvan, J.B.; Wozniak, A.; Hatcher, P.G.; Kaiser, K.; et al. The Role of Microbially-Mediated Exopolymeric Substances (EPS) in Regulating Macondo Oil Transport in a Mesocosm Experiment. Mar. Chem. 2018, 206, 52–61. [Google Scholar] [CrossRef]
- Doyle, S.M.; Whitaker, E.A.; De Pascuale, V.; Wade, T.L.; Knap, A.H.; Santschi, P.H.; Quigg, A.; Sylvan, J.B. Rapid Formation of Microbe-Oil Aggregates and Changes in Community Composition in Coastal Surface Water Following Exposure to Oil and the Dispersant Corexit. Front. Microbiol. 2018, 9, 689. [Google Scholar] [CrossRef] [Green Version]
- Bretherton, L.; Kamalanathan, M.; Genzer, J.; Hillhouse, J.; Setta, S.; Liang, Y.; Brown, C.M.; Xu, C.; Sweet, J.; Passow, U.; et al. Response of Natural Phytoplankton Communities Exposed to Crude Oil and Chemical Dispersants during a Mesocosm Experiment. Aquat. Toxicol. 2019, 206, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Kleindienst, S.; Seidel, M.; Ziervogel, K.; Grim, S.; Loftis, K.; Harrison, S.; Malkin, S.Y.; Perkins, M.J.; Field, J.; Sogin, M.L.; et al. Chemical Dispersants Can Suppress the Activity of Natural Oil-Degrading Microorganisms. Proc. Natl. Acad. Sci. USA 2015, 112, 14900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziervogel, K.; D’souza, N.; Sweet, J.; Yan, B.; Passow, U. Natural Oil Slicks Fuel Surface Water Microbial Activities in the Northern Gulf of Mexico. Front. Microbiol. 2014, 5, 188. [Google Scholar] [CrossRef] [PubMed]
- Kamalanathan, M.; Xu, C.; Schwehr, K.; Bretherton, L.; Beaver, M.; Doyle, S.M.; Genzer, J.; Hillhouse, J.; Sylvan, J.B.; Santschi, P.; et al. Extracellular Enzyme Activity Profile in a Chemically Enhanced Water Accommodated Fraction of Surrogate Oil: Toward Understanding Microbial Activities After the Deepwater Horizon Oil Spill. Front. Microbiol. 2018, 9, 798. [Google Scholar] [CrossRef]
- Kamalanathan, M.; Doyle, S.M.; Xu, C.; Achberger, A.M.; Wade, T.L.; Schwehr, K.; Santschi, P.H.; Sylvan, J.B.; Quigg, A. Exoenzymes as a Signature of Microbial Response to Marine Environmental Conditions. mSystems 2020, 5, e00290-20. [Google Scholar] [CrossRef] [Green Version]
- Wade, T.L.; Sweet, S.T.; Sericano, J.L.; Guinasso, N.L., Jr.; Diercks, A.-R.; Highsmith, R.C.; Asper, V.L.; Joung, D.; Shiller, A.M.; Lohrenz, S.E.; et al. Analyses of Water Samples From the Deepwater Horizon Oil Spill: Documentation of the Subsurface Plume. In Monitoring and Modeling the Deepwater Horizon Oil Spill: A Record-Breaking Enterprise; American Geophysical Union (AGU): Washington, DC, USA, 2011; pp. 77–82. ISBN 978-1-118-66675-3. [Google Scholar]
- Bera, G.; Gold-Bouchot, G.; Passow, U.; Wade, T.L.; Shi, D.; Morales-McDevitt, M.; Ramirez-Miss, N.; Knap, A.H.; Quigg, A. Inter-Laboratory Calibration of Estimated Oil Equivalent (EOE) Concentrations of a Water Accommodated Fraction (WAF) of Oil and a Chemically Enhanced WAF (CEWAF). Heliyon 2019, 5, e01174. [Google Scholar] [CrossRef] [Green Version]
- Morales-McDevitt, M.E.; Shi, D.; Knap, A.H.; Quigg, A.; Sweet, S.T.; Sericano, J.L.; Wade, T.L. Mesocosm Experiments to Better Understand Hydrocarbon Half-Lives for Oil and Oil Dispersant Mixtures. PLoS ONE 2020, 15, e0228554. [Google Scholar] [CrossRef]
- Medeiros, P.M.; Bícego, M.C. Investigation of Natural and Anthropogenic Hydrocarbon Inputs in Sediments Using Geochemical Markers. II. São Sebastião, SP––Brazil. Mar. Pollut. Bull. 2004, 49, 892–899. [Google Scholar] [CrossRef]
- Yamada, M.; Takada, H.; Toyoda, K.; Yoshida, A.; Shibata, A.; Nomura, H.; Wada, M.; Nishimura, M.; Okamoto, K.; Ohwada, K. Study on the Fate of Petroleum-Derived Polycyclic Aromatic Hydrocarbons (PAHs) and the Effect of Chemical Dispersant Using an Enclosed Ecosystem, Mesocosm. Environ. Manag. Enclosed Coast. Seas 2003, 47, 105–113. [Google Scholar] [CrossRef]
- Doyle, S.M.; Lin, G.; Morales-McDevitt, M.; Wade Terry, L.; Quigg, A.; Sylvan Jason, B. Niche Partitioning between Coastal and Offshore Shelf Waters Results in Differential Expression of Alkane and Polycyclic Aromatic Hydrocarbon Catabolic Pathways. mSystems 2020, 5, e00668-20. [Google Scholar] [CrossRef]
- Kadri, T.; Rouissi, T.; Kaur Brar, S.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of Polycyclic Aromatic Hydrocarbons (PAHs) by Fungal Enzymes: A Review. J. Environ. Sci. 2017, 51, 52–74. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.-H.; Xiong, A.-S.; Xue, Y.; Fu, X.-Y.; Gao, F.; Zhao, W.; Tian, Y.-S.; Yao, Q.-H. Microbial Biodegradation of Polyaromatic Hydrocarbons. FEMS Microbiol. Rev. 2008, 32, 927–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driskell, W.B.; Payne, J.R. Macondo Oil in Northern Gulf of Mexico Waters–Part 2: Dispersant-Accelerated PAH Dissolution in the Deepwater Horizon Plume. Mar. Pollut. Bull. 2018, 129, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuijdgeest, A.; Huettel, M. Dispersants as Used in Response to the MC252-Spill Lead to Higher Mobility of Polycyclic Aromatic Hydrocarbons in Oil-Contaminated Gulf of Mexico Sand. PLoS ONE 2012, 7, e50549. [Google Scholar] [CrossRef]
- Rughöft, S.; Vogel, A.L.; Joye, S.B.; Gutierrez, T.; Kleindienst, S. Starvation-Dependent Inhibition of the Hydrocarbon Degrader Marinobacter Sp. TT1 by a Chemical Dispersant. J. Mar. Sci. Eng. 2020, 8, 925. [Google Scholar] [CrossRef]
- Seidel, M.; Kleindienst, S.; Dittmar, T.; Joye, S.B.; Medeiros, P.M. Biodegradation of Crude Oil and Dispersants in Deep Seawater from the Gulf of Mexico: Insights from Ultra-High Resolution Mass Spectrometry. Deep Sea Res. Part II Top. Stud. Oceanogr. 2016, 129, 108–118. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Substrate | Empirical Formula | Millipore Sigma Product Number |
|---|---|---|
| 4-Methylumbelliferyl butyrate (MUF BU) | C14H14O4 | 19362 |
| 4-Methylumbelliferyl stearate (MUF ST) | C28H42O4 | M1010 |
| 4-Methylumbelliferyl palmitate (MUF PA) | C26H38O4 | M7259 |
| 4-Methylumbelliferyl oleate (MUF OL) | C28H40O4 | 75164 |
| Sampling Location; Type of Sample; Sampling Depth | Substrates Assayed | Comments | References |
|---|---|---|---|
| NW Atlantic; Intertidal sediments (Gulf of Maine) | MUF ST, MUF PA | MUF ST hydrolysis not detectable; MUF PA hydrolysis rates up to one order of magnitude lower than glucosidases. | [37] |
| NE Atlantic; deep-sea sediments (4500 m) | MUF ST | MUF ST hydrolysis in the same range as glucosidases, and not stimulated by fresh DOM. | [32,36] |
| Arabian Sea; deep-sea sediments (3000–4500 m) | MUF BU, MUF ST | MUF ST hydrolysis at detection limit and one order of magnitude lower than MUF BU. | [33] |
| Northern Gulf of Mexio; surface water near DwH spill site | MUF BU | MUF BU hydrolysis up to four times higher in oil-contaminated compared with uncontaminated surface waters. MUF BU two orders of magnitude higher than phosphatases. | [10] |
| MUF BU | MUF BU hydrolysis in MOS double that of glucosidases. | [38] | |
| Northern Gulf of Mexico; deep-water oil plume | MUF BU | MUF BU hydrolysis 1.75 times higher in plume compared with non-plume water and one order of magnitude higher than glucosidases. | Ziervogel (unpubl.) |
| Northern Gulf of Mexico, deep-sea sediments affected by the DwH fallout (1500–1900 m) | MUF BU | MUF BU hydrolysis one to two orders of magnitude higher than glucosidases. | [39] |
| Northern Gulf of Mexico; resuspended deep-sea sediments near natural seeps (530–1600 m) | MUF BU | MUF BU hydrolysis one to two orders of magnitude higher compared with glucosidases. | [40] |
| n-C17/Pristane | n-C18/Phytane | n-C17/n-C18 | |
|---|---|---|---|
| WAF | r2 = 0.4365 (p = 0.053) | r2 = 0.4115 (p = 0.063) | r2 = 0.6953 (p = 0.005) |
| DCEWAF | r2 = 0.784 (p = 0.001) | r2 = 0.8808 (p < 0.001) | r2 = 0.265 (p = 0.156) |
| Hexadecane | n-Alkanes | Naphthalene | Phenanthrene | |
|---|---|---|---|---|
| WAF | −0.83 (p = 0.08) | −0.85 (p = 0.07) | −0.87 (p = 0.06) | −0.88 (p = 0.05) |
| CEWAF | −0.94 (p = 0.02) | −0.89 (p = 0.04) | −0.85 (p = 0.07) | −0.92 (p = 0.03) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziervogel, K.; Kamalanathan, M.; Quigg, A. Hydrolysis of Methylumbeliferyl Substrate Proxies for Esterase Activities as Indicator for Microbial Oil Degradation in the Ocean: Evidence from Observations in the Aftermath of the Deepwater Horizon Oil Spill (Gulf of Mexico). J. Mar. Sci. Eng. 2022, 10, 583. https://doi.org/10.3390/jmse10050583
Ziervogel K, Kamalanathan M, Quigg A. Hydrolysis of Methylumbeliferyl Substrate Proxies for Esterase Activities as Indicator for Microbial Oil Degradation in the Ocean: Evidence from Observations in the Aftermath of the Deepwater Horizon Oil Spill (Gulf of Mexico). Journal of Marine Science and Engineering. 2022; 10(5):583. https://doi.org/10.3390/jmse10050583
Chicago/Turabian StyleZiervogel, Kai, Manoj Kamalanathan, and Antonietta Quigg. 2022. "Hydrolysis of Methylumbeliferyl Substrate Proxies for Esterase Activities as Indicator for Microbial Oil Degradation in the Ocean: Evidence from Observations in the Aftermath of the Deepwater Horizon Oil Spill (Gulf of Mexico)" Journal of Marine Science and Engineering 10, no. 5: 583. https://doi.org/10.3390/jmse10050583