
All-In-One: Microbial Response to Natural and Anthropogenic Forcings in a Coastal Mediterranean Ecosystem, the Syracuse Bay (Ionian Sea, Italy)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
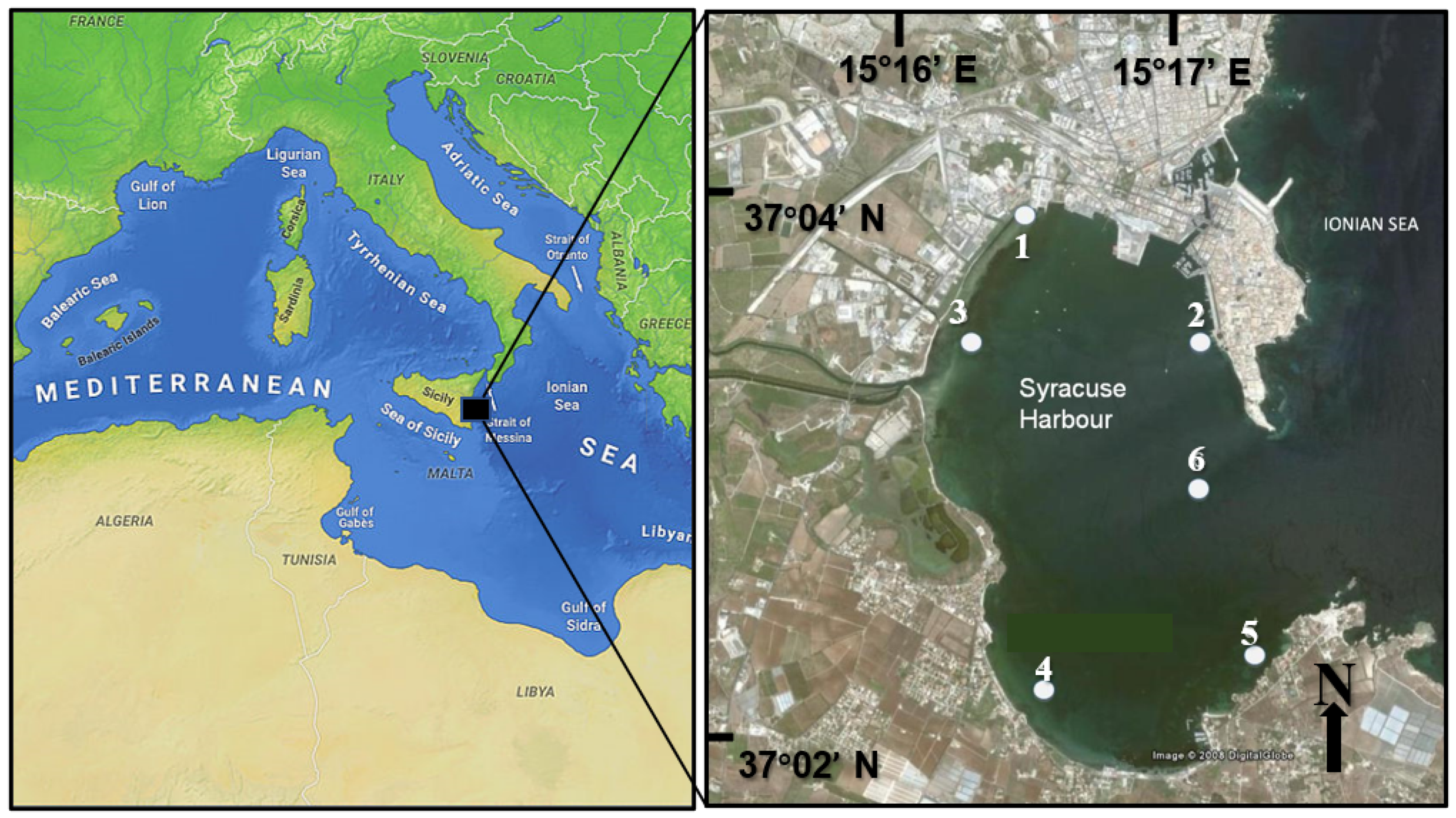
2.1. Site Description
2.2. Sample Collection
2.3. Physical and Chemical Parameters
2.4. Trophic State Index (TSI) Calculation as an Index of Water Quality
2.5. Microbial Parameters
2.5.1. Extracellular Enzymatic Activities (Leucine Aminopeptidase, LAP, Beta-Glucosidase, Beta-GLU and Alkaline Phosphatase, AP)
2.5.2. Culturable Heterotrophic Bacteria
2.5.3. Phytoplankton Samples
2.6. Statistical Analysis
3. Results and Discussion
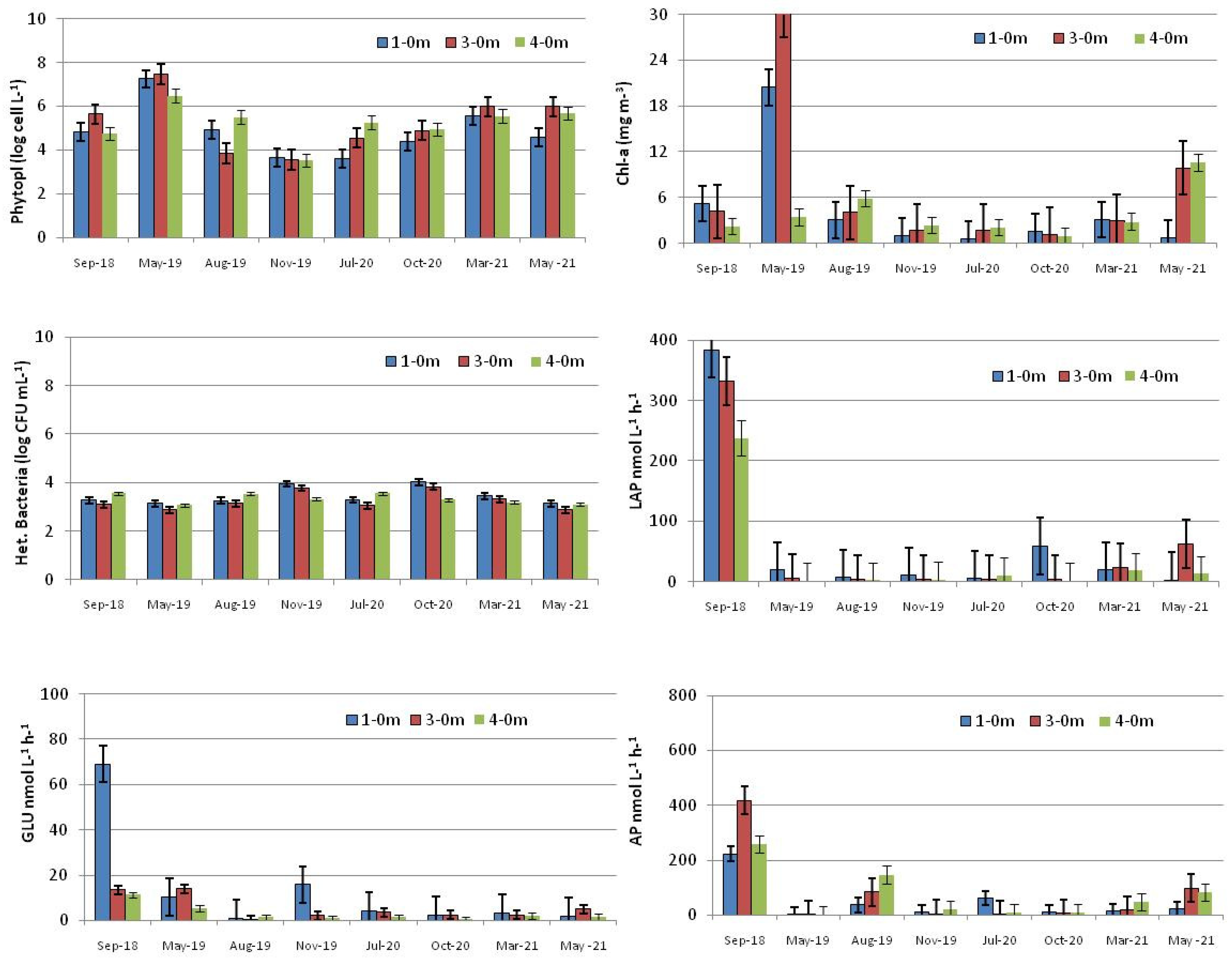
3.1. Human-Impacted Stations (HIS)
3.1.1. Environmental Parameters
3.1.2. Microbial Parameters
3.2. Freshwater-Affected Station (FAS)
3.2.1. Environmental Parameters
3.2.2. Microbial Parameters
3.3. Control Stations
3.3.1. Environmental Parameters
3.3.2. Microbial Parameters
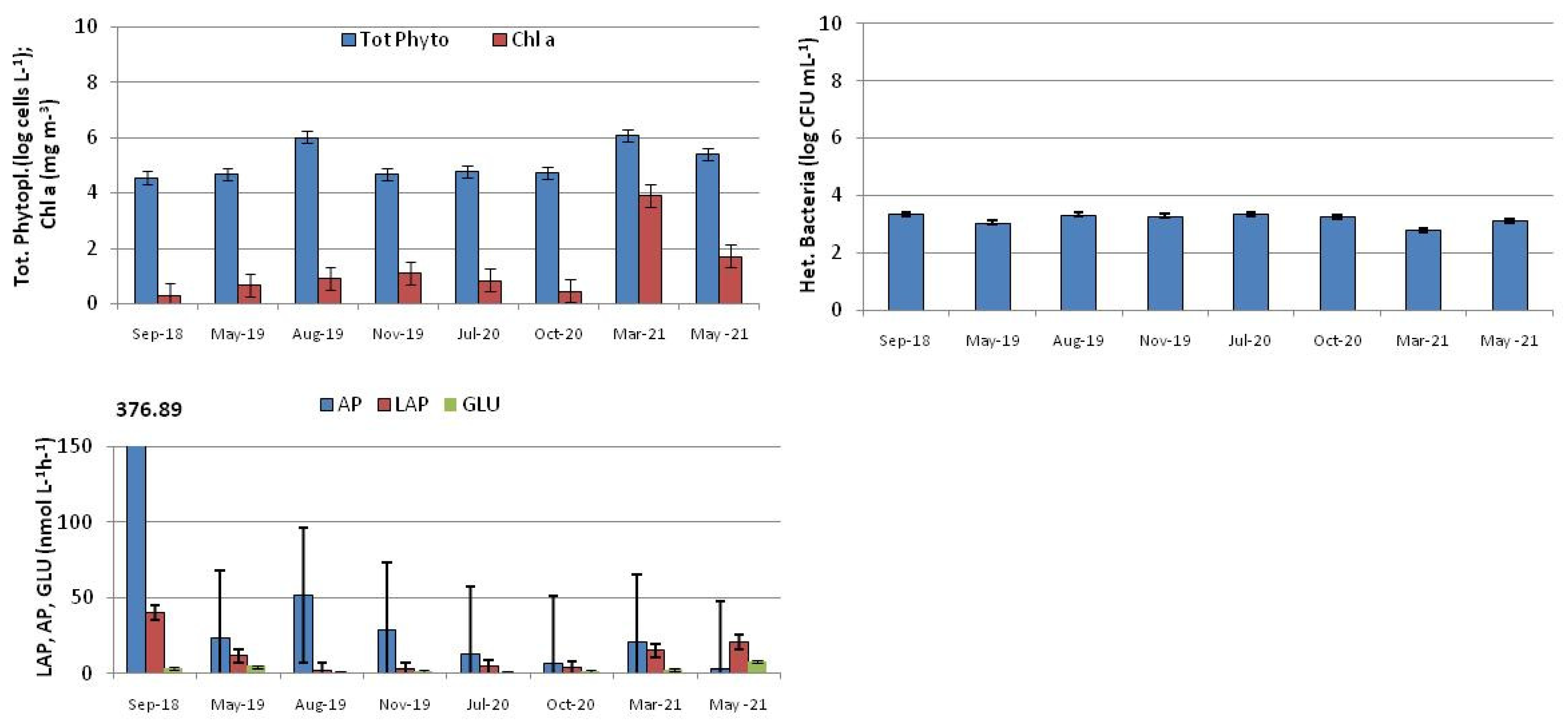
3.4. Water Quality Status Assessment of the Entire Syracuse Bay from the TSI Value
3.5. Qualitative Analysis of the Phytoplankton Community
3.6. Statistical Analysis of the Dataset
3.7. General Considerations
3.8. Microbial Response to Anthropic Activities
3.9. Microbial Response to Freshwater Inputs
3.10. Bacteria–Phytoplankton Interactions
3.11. Biogeochemical Implications of Microbial Activities and Related Stoichiometric Ratios
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coleman, J.M.; Huh, O.K.; DeWitt, B. Wetland loss in world deltas. J. Coast. Res. 2008, 24, 1–14. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [Green Version]
- Bonamano, S.; Piermattei, V.; Madonia, A.; Paladini de Mendoza, F.; Pierattini, A.; Martellucci, R.; Stefanì, C.; Zappalà, G.; Caruso, G.; Marcelli, M. The Civitavecchia Coastal Environment Monitoring System (C-CEMS): A new tool to analyze the conflicts between coastal pressures and sensitivity areas. Ocean Sci. 2016, 12, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Elliot, M.; Quintino, V. The estuarine quality paradox, environmental homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar. Pollut. Bull. 2007, 54, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Mitigating Harmful Cyanobacterial Blooms in a Human- and Climatically-Impacted World. Life 2014, 4, 988–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogales, B.; Lanfranconi, M.P.; Piña-Villalonga, J.M.; Bosch, R. Anthropogenic perturbations in marine microbial communities. FEMS Microbiol. Rev. 2011, 35, 275–298. [Google Scholar] [CrossRef] [Green Version]
- Buchan, A.; LeCleir, G.; Gulvik, C.; Gonzalez, J.M. Master recyclers: Features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Microbiol. 2014, 12, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Ponomarova, O.; Patil, K.R. Metabolic interactions in microbial communities: Untangling the Gordian knot. Curr. Opin. Microbiol. 2015, 27, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Amin, S.A.; Parker, M.S.; Armbrust, E.V. Interactions between diatoms and bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 667–684. [Google Scholar] [CrossRef] [Green Version]
- Bratbak, G.; Thingstad, T.F. Phytoplankton-bacteria interactions: An apparent paradox? Analysis of a model system with both competition and commensalism. Mar. Ecol. Progr. Ser. 1985, 25, 23–30. [Google Scholar] [CrossRef]
- Grossart, H.P.; Ploug, H. Microbial degradation of organic carbon and nitrogen on diatom aggregates. Limnol. Oceanogr. 2001, 46, 267–277. [Google Scholar] [CrossRef]
- Grossart, H.-P.; Czub, G.; Simon, M. Algae-bacteria interactions and their effects on aggregation and organic matter flux in the sea. Environ. Microbiol. 2006, 8, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Rubino, F.; Belmonte, M.; Caroppo, C.; Giacobbe, M.G. Dinoflagellate resting stages from surface sediments of Syracuse Bay (Western Ionian Sea, Mediterranean). Deep. Sea Res. II 2010, 57, 243–247. [Google Scholar] [CrossRef]
- Dell’Aversano, C.; Tartaglione, L.; Polito, G.; Dean, K.; Giacobbe, M.G.; Casabianca, S.; Capellacci, S.; Penna, A.; Turner, D. First detection of tetrodotoxin and high levels of paralytic shellfish poisoning toxins in shellfish from Sicily (Italy) by three different analytical methods. Chemosphere 2019, 215, 881–892. [Google Scholar] [CrossRef]
- Vila, M.; Giacobbe, M.G.; Masó, M.; Gangemi, E.; Penna, A.; Sampedro, N.; Azzaro, F.; Camp, J.; Galluzzi, L. A comparative study on recurrent blooms of Alexandrium minutum in two Mediterranean harbours. Harmful Algae 2005, 4, 673–695. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A practical handbook of seawater analysis. second ed. Fish. Res. Bd. Can. 1972, 167, 1–311. [Google Scholar]
- Aminot, A.; Chaussepied, M. Manuel des Analyses Chimiques en Milieu Marin; Centre National pour l’Exploration des Oceans: Brest, France, 1983; pp. 1–395. [Google Scholar]
- Valderrama, J.C. The simultaneous analysis of total nitrogen and total phosphorus in natural waters. Mar. Chem. 1981, 10, 109–122. [Google Scholar] [CrossRef]
- Koroleff, F. Determination of Phosphorus. In Methods of Seawater Analysis; Grasshoff, K., Ehrhardt, M., Kremling, K., Eds.; Verlag Chemie GmbH: Weinheim, Germany, 1983; pp. 125–139. [Google Scholar]
- Koroleff, F. Total and Organic Nitrogen. In Methods of Seawater Analysis; Grasshoff, K., Ehrhardt, M., Kremling, K., Eds.; Verlag Chemie GmbH: Weinheim, Germany, 1983; pp. 162–173. [Google Scholar]
- Lorenzen, C.J. Determination of chlorophyll and pheo-pigments: Spectrophotometric equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- Iseki, K.; MacDonald, R.W.; Carmack, E. Distribution of particulate matter in the Southeastern Beaufort Sea in late summer. Proc. NIPR Symp. Polar Biol. 1987, 1, 35–46. [Google Scholar]
- University of Southern Florida Water Institute (TSI Trophic State Index). Available online: http://www.lake.wateratlas.usf.edu/library/learn-more/learnmore.aspx?toolsection=lm_tsi (accessed on 24 June 2021).
- Monticelli, L.S.; Caruso, G.; Decembrini, F.; Caroppo, C.; Fiesoletti, F. Role of prokaryotic biomasses and activities in Carbon and Phosphorus cycles at a coastal, thermohaline front and in offshore waters (Gulf of Manfredonia, Southern Adriatic Sea). Microb. Ecol. 2014, 67, 501–519. [Google Scholar] [CrossRef]
- Zaccone, R.; Caruso, G.; Calì, C. Heterotrophic bacteria in the northern Adriatic Sea: Seasonal changes and ectoenzyme profile. Mar. Environ. Res. 2002, 54, 1–19. [Google Scholar] [CrossRef]
- Bužančić, M.; Ninčević Gladan, Z.; Marasović, I.; Kušpilić, G.; Grbec, B. Eutrophication influence on phytoplankton community composition in three bays on the eastern Adriatic coast. Oceanologia 2016, 58, 302–316. [Google Scholar] [CrossRef] [Green Version]
- Edler, L.; Elbrächter, M. The Utermöhl method for quantitative phytoplankton analysis. In Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis; IOC Manuals and Guides n. 55; Karlson, B., Cusack, C., Bresnan, E., Eds.; UNESCO: Paris, France, 2010; pp. 13–20. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software package for education and data analysis. Paleontol. Electronica 2001, 4, 1–9. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Similarity-based testing for community pattern: The 2-way layout with no replication. Mar. Biol. 1994, 118, 167–176. [Google Scholar] [CrossRef]
- Caruso, G.; Azzaro, M.; Monticelli, L.S.; Leonardi, M.; Cao, X.Y.; Zhou, Y.Y.; Song, C.L. Seasonal variations in microbial parameters and trophic states in a large Chinese shallow lake (Lake Taihu). Fresenius Environ. Bull. 2017, 26, 785–796. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial (Plymouth Routines in Multivariate Ecological Research); Primer-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Adams, S.M. Assessing cause and effect of multiple stressors on marine systems. Mar. Pollut. Bull. 2005, 51, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Zaccone, R.; Caruso, G. Microbial enzymes in the Mediterranean Sea: Relationship with climate changes. AIMS Microbiol. 2019, 5, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Celussi, M.; Zoccarato, L.; Bernardi Aubry, F.; Bastianini, M.; Casotti, R.; Balestra, C.; Giani, M.; Del Negro, P. Links between microbial processing of organic matter and the thermohaline and productivity features of a temperate river-influenced Mediterranean coastal area. Estuar. Coast. Shelf Sci. 2019, 228, 106378. [Google Scholar] [CrossRef]
- Zaccone, R.; Caroppo, C.; La Ferla, R.; Zampino, D.; Caruso, G.; Leonardi, M.; Maimone, G.; Azzaro, M.; Sitran, R. Deep-chlorophyll maximum time series in the Augusta Gulf (Ionian Sea): Microbial community structures and functions. Chem. Ecol. 2004, 20 (Suppl. 1), 267–284. [Google Scholar] [CrossRef]
- Caruso, G.; Azzaro, F.; Azzaro, M.; Decembrini, F.; La Ferla, R.; Maimone, G.; De Pasquale, F.; Monticelli, L.S.; Zaccone, R.; Zappalà, G.; et al. Environmental variability in a transitional Mediterranean system (Oliveri-Tindari, Italy): Focusing on the response of microbial activities and prokaryotic abundance. Estuar. Coastal Shelf Sci. 2013, 135, 158–170. [Google Scholar] [CrossRef]
- Decembrini, F.; Caroppo, C.; Caruso, G.; Bergamasco, A. Linking Microbial Functioning and Trophic Pathways to Ecological Status in a Coastal Mediterranean Ecosystem. Water 2021, 13, 1325. [Google Scholar] [CrossRef]
- Zaccone, R.; Azzaro, M.; Azzaro, F.; Bergamasco, A.; Caruso, G.; Leonardi, M.; La Ferla, R.; Maimone, G.; Mancuso, M.; Monticelli, L.S.; et al. Seasonal Dynamics of Prokaryotic Abundance and Activities in Relation to Environmental Parameters in a Transitional Aquatic Ecosystem (Cape Peloro, Italy). Microb. Ecol. 2014, 67, 45–56. [Google Scholar] [CrossRef]
- Li, Z.; Xing, M.; Wang, W.; Wang, D.; Zhu, J.; Sun, M. Phylogenetic diversity of culturable bacteria in surface seawater from the Drake Passage, Antarctica. Chin. J. Oceanol. Limnol. 2016, 34, 952–963. [Google Scholar] [CrossRef]
- Islam, M.S.; Tanaka, M. Impacts of pollution on coastal and marine ecosystems including coastal and marine fisheries and approach for management: A review and synthesis. Mar. Pollut. Bull. 2004, 48, 624–649. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Ma, J.; Murinda, S.E. Bacterial community composition and structure in an Urban River impacted by different pollutant sources. Sci. Total. Environ. 2016, 566–567, 1176–1185. [Google Scholar] [CrossRef] [Green Version]
- Liao, K.; Bai, Y.; Huo, Y.; Jian, Z.; Hu, W.; Zhao, C.; Qu, J. Integrating microbial biomass, composition and function to discern the level of anthropogenic activity in a river ecosystem. Environ. Int. 2018, 116, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Edgcumbe Ford, T. Response of marine microbial communities to anthropogenic stress. J. Aquat. Ecosyst. Stress Recovery 2000, 7, 75–89. [Google Scholar] [CrossRef]
- Teira, E.; Hernando-Morales, V.; Martínez-García, S.; Figueiras, F.G.; Arbones, B.; Álvarez-Salgado, X.A. Response of bacterial community structure and function to experimental rainwater additions in a coastal eutrophic embayment. Estuar. Coast. Shelf Sci. 2013, 119, 44–53. [Google Scholar] [CrossRef]
- Teira, E.; Hernández-Ruiz, M.; Barber-Lluch, E.; Sobrino, C.; Teixeira, I.G.; Álvarez-Salgado, X.A.; Nieto-Cid, M.; Martínez-García, S.; Figueiras, F.G.; Fernández, E. Bacterioplankton responses to riverine and atmospheric inputs in a coastal upwelling system (Ría de Vigo, NW Spain). Mar. Ecol. Prog. Ser. 2016, 542, 39–50. [Google Scholar] [CrossRef]
- Gutiérrez-Barral, A.; Teira, E.; Hernández-Ruiz, M.; Fernández, E. Response of prokaryotic community composition to riverine and atmospheric nutrients in a coastal embayment: Role of organic matter on Vibrionales. Estuar. Coast. Shelf Sci. 2021, 251, 107196. [Google Scholar] [CrossRef]
- Catalano, G.; Azzaro, M.; Bastianini, M.; Bellucci, L.G.; Bernardi Aubry, F.; Bianchi, F.; Burca, M.; Cantoni, C.; Caruso, G.; Casotti, R.; et al. The Carbon budget in the northern Adriatic Sea, a winter case study. J. Geophys. Res. Biogeosci. 2014, 119, 1399–1417. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Zaccone, R. Estimates of leucine aminopeptidase activity in different marine and brackish environments. J. Appl. Microbiol. 2000, 89, 951–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Jiang, X.; Jing, Z.; Li, G.; Chen, Z.; Zhou, L.; Zhao, C.; Liu, J.; Tan, Y. Spatial and seasonal distributions of bacterioplankton in the Pearl River Estuary: The combined effects of riverine inputs, temperature, and phytoplankton. Mar. Pollut. Bull. 2017, 125, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Cibic, T.; Fazi, S.; Nasi, F.; Pin, L.; Alvisi, F.; Berto, D.; Vigano, L.; Zoppini, A.; Del Negro, P. Natural and anthropogenic disturbances shape benthic phototrophic and heterotrophic microbial communities in the Po River Delta system. Estuar. Coast. Shelf Sci. 2019, 222, 168–182. [Google Scholar] [CrossRef]
- Povinec, P.P.; Aggarwal, P.K.; Aureli, A.; Burnett, W.C.; Kontar, E.A.; Kulkarni, K.M.; Moore, W.S.; Rajar, R.; Taniguchi, M.; Comanducci, J.-F.; et al. Characterisation of submarine groundwater discharge offshore south-eastern Sicily. J. Environ. Radioactivity 2006, 89, 81–101. [Google Scholar] [CrossRef]
- Bi, R.; Cao, Z.; Ismar-Rebitz, S.M.H.; Sommer, U.; Zhang, H.; Ding, Y.; Zhao, M. Responses of marine Diatom-Dinoflagellate competition to multiple environmental drivers: Abundance, elemental, and biochemical aspects. Front. Microbiol. 2021, 12, 731786. [Google Scholar] [CrossRef]
- Teeling, H.; Fuchs, B.M.; Becher, D.; Klockow, C.; Gardebrecht, A.; Bennke, C.M.; Kassabgy, M.; Huang, S.; Mann, A.J.; Waldmann, J.; et al. Substrate-controlled succession of marine bacterioplankton populations induced by a phytoplankton bloom. Science 2012, 336, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Gasol, J.M.; Estrada, M.; Franco-Vidal, L.; Díaz-Pérez, L.; Ferrera, I.; Morán, X.A.G. Heterotrophic bacterial responses to the winter–spring phytoplankton bloom in open waters of the NW Mediterranean. Deep Sea Res. Part I 2015, 96, 59–68. [Google Scholar] [CrossRef]
- Sarmento, H.; Gasol, J.M. Use of phytoplankton-derived dissolved organic carbon by different types of bacterioplankton. Environ. Microbiol. 2012, 14, 2348–2360. [Google Scholar] [CrossRef]
- Biddanda, B.; Benner, R. Carbon, nitrogen, and carbohydrate fluxes during the production of particulate and dissolved organic matter by marine phytoplankton. Limnol. Oceanogr. 1997, 42, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xing, P.; Wu, Q.L. The high resilience of the bacterioplankton community in the face of a catastrophic disturbance by a heavy Microcystis bloom. FEMS Microbiol. Ecol. 2012, 82, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zhang, H.; Xiong, J.; Wang, K.; Zhu, J.; Zhu, X.; Zhou, X.; Zhang, D. Successional trajectories of bacterioplankton community over the complete cycle of a sudden phytoplankton bloom in the Xiangshan Bay, East China Sea. Environ. Poll. 2016, 219, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Sintes, E.; Witte, H.; Stodderegger, K.; Steiner, P.; Herndl, G.J. Temporal dynamics in the free-living bacterial community composition in the coastal North Sea. FEMS Microbiol. Ecol. 2013, 83, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Franzo, A.; Cibic, T.; Del Negro, P.; Solidoro, C. Microphytobenthic response to mussel farm biodeposition in coastal sediments of the northern Adriatic Sea. Mar. Pollut. Bull. 2014, 79, 379–388. [Google Scholar] [CrossRef]
- Valenzuela-Sanchez, C.G.; Pasten-Miranda, N.M.A.; Enriquez-Ocaña, L.F.; Barraza-Guardado, R.H.; Valdez Holguin, J.E.; Martinez-Cordova, L.R. Phytoplankton composition and abundance as indicators of aquaculture effluents impact in coastal environments of mid Gulf of California. Heliyon 2021, 7, e06203. [Google Scholar] [CrossRef] [PubMed]
- Cibic, T.; Comici, C.; Bussani, A.; Del Negro, P. Benthic diatom response to changing environmental conditions. Estuar. Coast. Shelf Sci. 2012, 115, 158–169. [Google Scholar] [CrossRef]
- Falkowski, P.G. The Role of Phytoplankton Photosynthesis in Global Biogeochemical Cycles. Photosynthesis Res. 1994, 39, 235–258. [Google Scholar] [CrossRef] [Green Version]
- Cunha, A.; Almeida, A.; Coelho, F.J.R.C.; Gomes, N.C.M.; Oliveira, V.; Santos, A.L. Bacterial Extracellular Enzymatic Activity in Globally Changing Aquatic Ecosystems. In Current Research, Technology and Education. Topics in Applied Microbiology and Microbial Technology; Méndez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2010; pp. 124–135. [Google Scholar]
- Rekik, A.; Denis, M.; Dugenne, M.; Barani, A.; Maalej, S.; Ayadi, H. Seasonal distribution of ultraphytoplankton and heterotrophic prokaryotes in relation to abiotic variables on the north coast of Sfax after restoration. Mar. Pollut. Bull. 2014, 84, 280–305. [Google Scholar] [CrossRef]
- Richa, K.; Balestra, C.; Piredda, R.; Benes, V.; Borra, M.; Passarelli, A.; Margiotta, F.; Saggiomo, M.; Biffali, E.; Sanges, R.; et al. Distribution, Community composition, and potential metabolic activityof bacterioplankton in an urbanized Mediterranean Sea coastal zone. Appl. Environ. Microbiol. 2017, 83, e00494-17. [Google Scholar] [CrossRef] [Green Version]
- Misic, C.; Castellano, M.; Covazzi Harriague, A. Organic matter features, degradation and remineralisation at two coastal sites he Ligurian Sea (NW Mediterranean) differently influenced by anthropogenic forcing. Mar. Environ. Res. 2011, 72, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems a global problem. Environ. Sci. Poll. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Malone, T.C.; Newton, A. The globalization of cultural eutrophication in the coastal ocean: Causes and consequences. Front. Mar. Sci. 2020, 7, 670. [Google Scholar] [CrossRef]
- European Commission. Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 Establishing a Framework for Community Action in the Field of Marine environmental Policy. Available online: https://eur-lex.europa.eu/legal-content/EL/TXT/PDF/?uri=CELEX:32008L0056&from=EN (accessed on 24 March 2020).
- Ferreira, J.G.; Andersen, J.H.; Borja, A.; Bricker, S.B.; Camp, J.; Cardoso da Silva, M.; Garcés, E.; Heiskanen, A.-S.; Humborg, C.; Ignatiades, L.; et al. Overview of eutrophication indicators to assess environmental status within the European Marine Strategy Framework Directive. Estuar. Coast. Shelf Sci. 2011, 93, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Martiny, A.C.; Vrugt, J.A.; Lomas, M.W. Concentrations and ratios of particulate organic carbon, nitrogen, and phosphorus in the global ocean. Sci. Data 2014, 1, 140048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redfield, A.C. The biological control of chemical factors in the environment. Sci. Prog. 1960, 11, 150–170. [Google Scholar]
- Friedrich, J.; Janssen, F.; Aleynik, D.; Bange, H.W.; Boltacheva, N.; Çagatay, M.N.; Dale, A.W.; Etiope, G.; Erdem, Z.; Geraga, M.; et al. Investigating hypoxia in aquatic environments: Diverse approaches to addressing a complex phenomenon. Biogeoscience 2014, 11, 1215–1259. [Google Scholar] [CrossRef] [Green Version]
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nature 2008, 14, 293–296. [Google Scholar] [CrossRef]
- Song, C.; Cao, X.; Zhou, Y.; Azzaro, M.; Monticelli, L.S.; Leonardi, M.; La Ferla, R.; Caruso, G. Different pathways of nitrogen and phosphorus regeneration mediated by extracellular enzymes in temperate lakes under various trophic state. Environ. Sci. Pollut. Res. 2018, 25, 31603–31615. [Google Scholar] [CrossRef]
- Granéli, E.; Wallström, K.; Larsson, U.; Granéli, W.; Elmgren, R. Nutrient limitation of primary production in the Baltic Sea area. Ambio 1990, 19, 142–151. [Google Scholar]
- Garmendia, M.; Borja, Á.; Franco, J.; Revilla, M. Phytoplankton composition indicators for the assessment of eutrophication in marine waters: Present state and challenges within the European directives. Mar. Pollut. Bull. 2013, 66, 7–16. [Google Scholar] [CrossRef]
- Marić, D.; Kraus, R.; Godrijan, J.; Supić, N.; Djakovac, T.; Precali, R. Phytoplankton response to climatic and anthropogenic influences in the north-eastern Adriatic during the last four decades. Estuar. Coast. Shelf Sci. 2012, 115, 98–112. [Google Scholar] [CrossRef]
- Tarafdar, L.; Kim, J.Y.; Srichandan, S.; Mohapatra, M.; Muduli, P.R.; Kumar, A.; Mishra, D.R.; Rastogi, G. Responses of phytoplankton community structure and association to variability in environmental drivers in a tropical coastal lagoon. Sci. Tot. Environ. 2021, 783, 146873. [Google Scholar] [CrossRef]
- Cruz de Carvalho, M.H.; Sun, H.X.; Bowler, C.; Chua, N.H. Noncoding and coding transcriptome responses of a marine diatom to phosphate fluctuations. New Phytol. 2016, 210, 497–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrzanowski, T.H.; Kyle, M.; Elser, J.J.; Sterner, R.W. Element ratios and growth dynamics of bacteria in an oligotrophic shield. Lake. Aquat. Microb. Ecol. 1996, 11, 119–125. [Google Scholar] [CrossRef]
- Lee, S.; Fuhrman, J.A. Relationships between biovolume and biomass of naturally derived marine bacterioplankton. Appl. Environ. Microbiol. 1987, 53, 1298–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, M.; Azzaro, F.; Azzaro, M.; Caruso, G.; Mancuso, M.; Monticelli, L.S.; Maimone, G.; La Ferla, R.; Raffa, F.; Zaccone, R. A multidisciplinary study of the Cape Peloro brackish area (Messina, Italy): Characterisation of trophic conditions, microbial abundances and activities. Mar. Ecol. 2009, 30, 33–42. [Google Scholar] [CrossRef]
- Perliński, P.; Mudryk, Z.J.; Antonowicz, J. Enzymatic activity in the surface microlayer and subsurface water in the harbour channel. Estuar. Coast. Shelf Sci. 2017, 196, 150–158. [Google Scholar] [CrossRef]
- Ivančić, I.; Kraus, R.; Najdek, M.; Cozzi, S. Ecological importance of Alkaline Phosphatase Activity in Changing Marine Environmental Conditions. Water 2021, 13, 2750. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Main Characteristics | Latitude N | Longitude E |
|---|---|---|---|
| 1 | Shipyard station, harbor | 37.064 | 15.279 |
| 2 | Aretusa springwater | 37.056 | 15.292 |
| 3 | Anapo-Ciane river mouth | 37.056 | 15.272 |
| 4 | Shellfish farm | 37.035 | 15.281 |
| 5 | Control marine station | 37.039 | 15.296 |
| 6 | Control marine station | 37.048 | 15.291 |
| Station | Main Characteristics | Squared Distance | Highly Contributing Parameters |
|---|---|---|---|
| 1 | Human-impacted | 29.64 | NO3, TN |
| 3 | Human-impacted | 34.37 | NH4, TN, PO4, S |
| 4 | Human-impacted | 14.74 | DO, TP, TN, NO3 |
| 2 | Freshwater-affected | 26.36 | S, TSM, NH4, NO3, DO |
| 5 | Control | 16.18 | TN, pH, DO |
| 6 | Control | 29.20 | pH, TSM, TN, S, DO |
| Stations | ||||||
|---|---|---|---|---|---|---|
| HIS | FAS | Control | ||||
| Parameters | 1 | 3 | 4 | 2 | 5 | 6 |
| Temperature (°C) | 22.77 ± 4.18 | 22.73 ± 5.05 | 22.01 ± 4.41 | 22.08 ± 4.57 | 22.72 ± 4.71 | 22.18 ± 4.88 |
| Salinity | 38.56 ± 0.35 | 38.06 ± 0.99 | 38.10 ± 0.64 | 37.55 ± 1.55 | 37.98 ± 0.29 | 38.10 ± 0.77 |
| Diss.Oxygen (mg/L) | 6.72 ± 0.96 | 7.22 ± 0.97 | 7.08 ± 1.04 | 6.87 ± 1.35 | 7.14 ± 1.10 | 7.65 ± 0.94 |
| pH | 8.22 ± 0.28 | 8.34 ± 0.48 | 8.24 ± 0.28 | 8.17 ± 0.33 | 8.31 ± 0.37 | 8.37 ± 0.49 |
| NH4(µM) | 1.06 ± 0.44 | 1.68± 1.17 | 1.13 ± 0.33 | 1.65 ± 1.65 | 1.15 ± 0.50 | 1.06 ± 0.51 |
| NO2 (µM) | 0.23 ± 0.29 | 0.61 ± 1.09 | 0.15 ± 0.09 | 0.20 ± 0.31 | 0.19 ± 0.09 | 0.18 ± 0.08 |
| NO3 (µM) | 3.90 ± 4.30 | 3.81 ± 2.52 | 3.91 ± 4.14 | 5.01 ± 5.45 | 2.07 ± 1.20 | 3.07 ± 2.29 |
| TN (µM) | 7.78 ± 5.00 | 9.75 ± 7.81 | 8.75 ± 6.83 | 8.15 ± 6.17 | 7.31 ± 6.90 | 7.39 ± 5.82 |
| PO4 (µM) | 0.33 ± 0.26 | 0.91 ± 1.23 | 0.26 ± 0.19 | 0.24 ± 0.12 | 0.25 ± 0.21 | 0.25 ± 0.13 |
| TP (µM) | 1.81 ± 0.77 | 2.56 ± 1.53 | 2.18 ± 2.10 | 1.59 ± 0.98 | 1.40 ± 0.54 | 1.38 ± 0.47 |
| N/P | 15.51 ± 14.11 | 6.74 ± 13.71 | 20.08 ± 23.12 | 28.58 ± 29.81 | 13.64 ± 18.16 | 17.24 ± 18.20 |
| TSM (mg/L) | 82.91 ± 136.83 | 75.41 ± 54.43 | 43.46 ± 42.47 | 172.92 ± 348.66 | 40.24 ± 22.41 | 120.80 ± 187.06 |
| POC µgC/L) | 818.23 ± 1497.82 | 2496.55 ± 5375.91 | 504.21 ± 409.47 | 333.01 ± 264.87 | 433.66 ± 425.30 | 553.87 ± 651.08 |
| PN (µgN/L) | 159.53 ± 332.92 | 430.50 ± 933.11 | 82.62 ± 61.36 | 45.04 ± 22.94 | 73.59 ± 63.75 | 97.17 ± 116.46 |
| C/N | 6.04 ± 0.92 | 6.02 ± 0.59 | 6.11 ± 1.21 | 7.17 ± 2.45 | 5.85 ± 0.84 | 5.86 ± 0.49 |
| Stations | ||||||
|---|---|---|---|---|---|---|
| HIS | FAS | Control | ||||
| Parameters | 1 | 3 | 4 | 2 | 5 | 6 |
| Tot.Phytopl. (cells/L) | 8.70 × 105 ± 3.66 × 106 | 3.99 × 106 ± 1.03 × 107 | 4.65 × 105 ± 8.79 × 105 | 3.39 × 105 ± 4.78 × 105 | 1.80 × 105 ± 1.88 × 105 | 5.77 × 106 ± 1.57 × 107 |
| Diatoms (cells/L) | 1.04 × 104 ± 1.90 × 104 | 6.09 × 104 ± 1.58 × 105 | 5.73 × 104 ± 1.64 × 105 | 1.31 × 105 ± 3.57 × 105 | 3.71 × 104 ± 9.15 × 104 | 5.57 × 106 ± 1.57 × 107 |
| Dinofl. (cells/L) | 8.92 × 105 ± 3.74 × 106 | 3.93 × 106 ± 1.03 × 107 | 3.57 × 105 ± 8.97 × 105 | 1.80 × 105 ± 4.04 × 106 | 8.19 × 104 ± 1.69 × 105 | 1.65 × 105 ± 2.36 × 105 |
| Others (cells/L) | 6.44 × 103 ± 9.67 × 103 | 1.13 × 104 ± 1.46 × 104 | 5.11 × 104 ± 9.54 × 104 | 2.76 × 104 ± 2.07 × 104 | 6.14 × 104 ± 1.05 × 105 | 4.01 × 104 ± 9.63 × 104 |
| Chl a (mg/m3) | 2.95 ± 4.04 | 7.02 ± 9.92 | 3.00 ± 2.39 | 1.23 ± 1.16 | 3.21 ± 2.16 | 3.63 ± 2.21 |
| Het. Bacteria (CFU/mL) | 2.88 × 103 ± 2.81 × 103 | 2.46 × 103 ± 2.37 × 103 | 2.39 × 103 ± 1.40 × 103 | 1.64 × 103 ± 5.84 × 102 | 1.21 × 103 ± 3.97 × 102 | 2.28 × 103 ± 9.24 × 102 |
| LAP (nmol/L/h) | 41.75 ± 81.45 | 54.58 ± 113.91 | 36.56 ± 70.55 | 12.31 ± 13.11 | 48.35 ± 94.40 | 36.43 ± 82.29 |
| GLU (nmol/L/h) | 6.91 ± 14.12 | 5.39 ± 5.36 | 3.42 ± 4.21 | 2.50 ± 2.51 | 2.69 ± 5.16 | 4.06 ± 4.81 |
| AP (nmol/L/h) | 61.77 ± 80.50 | 79.36 ± 142.33 | 91.25 ± 164.63 | 65.27 ± 126.83 | 82.56 ± 124.12 | 53.90 ± 105.16 |
| Variable | HIS | FAS | Controls | |||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | PC1 | PC2 | |
| T | 0.072 | 0.258 | −0.326 | 0.061 | −0.168 | 0.219 |
| S | 0.208 | −0.222 | 0.249 | 0.197 | 0.311 | −0.003 |
| DO | 0.027 | 0.136 | 0.257 | −0.183 | 0.072 | −0.353 |
| Transp | 0.194 | 0.111 | −0.287 | 0.013 | 0.245 | 0.088 |
| pH | −0.011 | 0.296 | −0.280 | −0.041 | −0.252 | 0.252 |
| TP | −0.122 | −0.237 | 0.103 | −0.112 | 0.145 | 0.101 |
| PO4 | 0.018 | −0.133 | −0.153 | 0.234 | −0.090 | 0.283 |
| TN | −0.261 | 0.256 | −0.107 | −0.365 | −0.285 | −0.237 |
| NH4 | −0.038 | 0.236 | 0.043 | −0.285 | −0.227 | −0.081 |
| NO2 | −0.042 | −0.147 | 0.059 | −0.218 | −0.085 | −0.163 |
| NO3 | −0.287 | 0.094 | −0.133 | −0.344 | −0.304 | −0.017 |
| -TSM | −0.145 | −0.178 | 0.015 | 0.082 | 0.123 | 0.030 |
| POC | −0.364 | −0.044 | −0.182 | 0.164 | −0.292 | 0.077 |
| PN | −0.371 | −0.047 | −0.053 | 0.018 | −0.298 | 0.018 |
| Tot Phytopl. | −0.367 | −0.076 | 0.067 | 0.276 | −0.120 | 0.262 |
| Diatoms | 0.010 | 0.310 | −0.228 | 0.228 | −0.123 | 0.263 |
| Dinoflagellates | −0.367 | −0.080 | 0.283 | 0.122 | 0.128 | −0.102 |
| Others | 0.096 | −0.115 | −0.025 | 0.080 | 0.163 | −0.050 |
| Chl a | −0.349 | −0.036 | 0.282 | 0.175 | −0.248 | 0.199 |
| Phaeo | −0.140 | 0.103 | 0.290 | 0.132 | 0.034 | 0.285 |
| Het.bacteria | 0.085 | −0.081 | −0.321 | 0.009 | 0.049 | 0.117 |
| LAP | −0.022 | 0.376 | 0.021 | −0.282 | −0.195 | −0.249 |
| GLU | −0.089 | 0.233 | 0.176 | −0.207 | −0.257 | −0.297 |
| AP | 0.011 | 0.410 | −0.161 | −0.234 | −0.233 | −0.126 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, G.; Giacobbe, M.G.; Azzaro, F.; Decembrini, F.; Leonardi, M.; Miserocchi, S.; Cao, X.; Song, C.; Zhou, Y. All-In-One: Microbial Response to Natural and Anthropogenic Forcings in a Coastal Mediterranean Ecosystem, the Syracuse Bay (Ionian Sea, Italy). J. Mar. Sci. Eng. 2022, 10, 19. https://doi.org/10.3390/jmse10010019
Caruso G, Giacobbe MG, Azzaro F, Decembrini F, Leonardi M, Miserocchi S, Cao X, Song C, Zhou Y. All-In-One: Microbial Response to Natural and Anthropogenic Forcings in a Coastal Mediterranean Ecosystem, the Syracuse Bay (Ionian Sea, Italy). Journal of Marine Science and Engineering. 2022; 10(1):19. https://doi.org/10.3390/jmse10010019
Chicago/Turabian StyleCaruso, Gabriella, Maria Grazia Giacobbe, Filippo Azzaro, Franco Decembrini, Marcella Leonardi, Stefano Miserocchi, Xiuyun Cao, Chunlei Song, and Yiyong Zhou. 2022. "All-In-One: Microbial Response to Natural and Anthropogenic Forcings in a Coastal Mediterranean Ecosystem, the Syracuse Bay (Ionian Sea, Italy)" Journal of Marine Science and Engineering 10, no. 1: 19. https://doi.org/10.3390/jmse10010019