
From Surface Water to the Deep Sea: A Review on Factors Affecting the Biodegradation of Spilled Oil in Marine Environment

 ,
,  ,
,  ,
,  ,
, 


 ,
,  ,
, 
Abstract
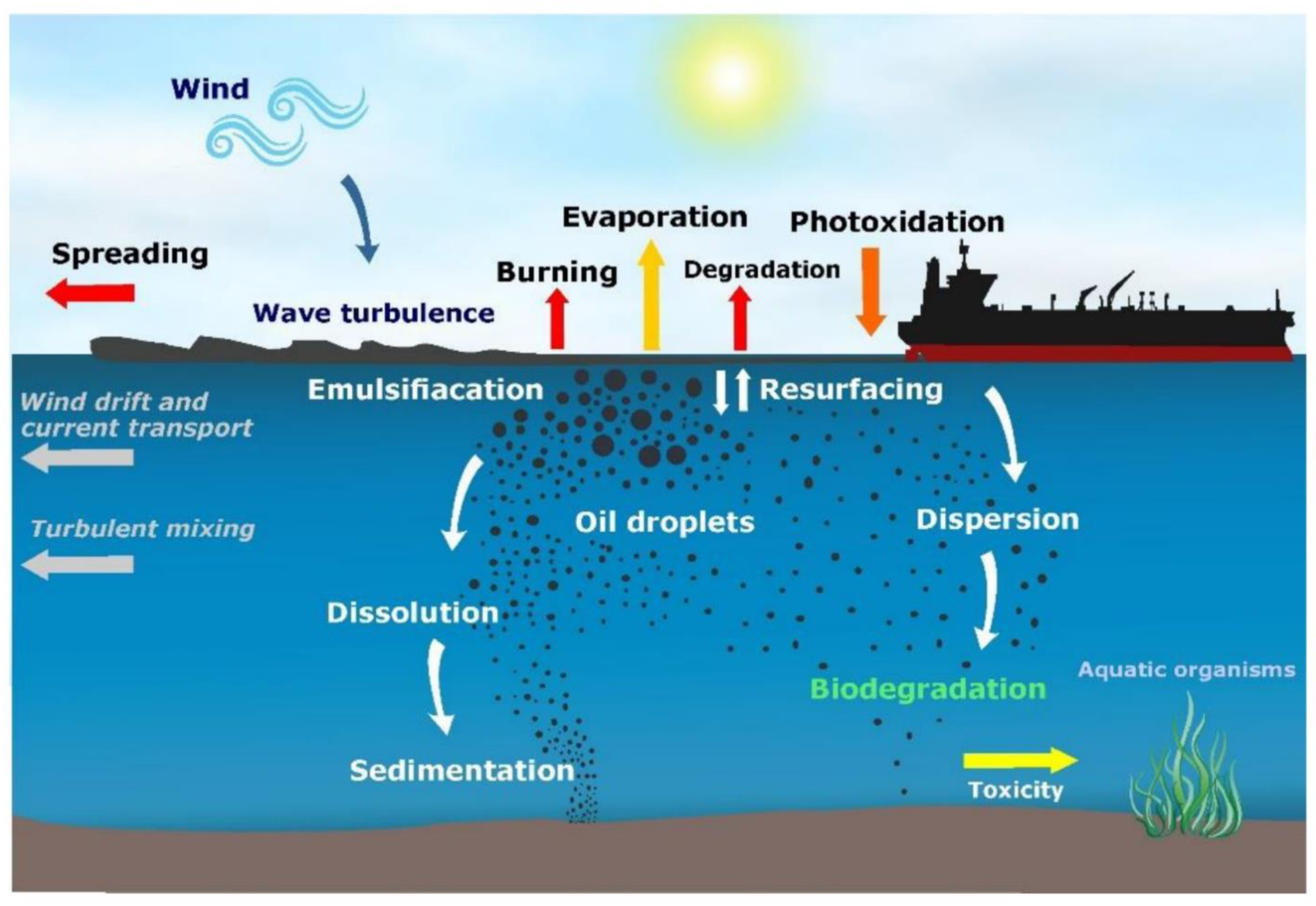
:1. Introduction
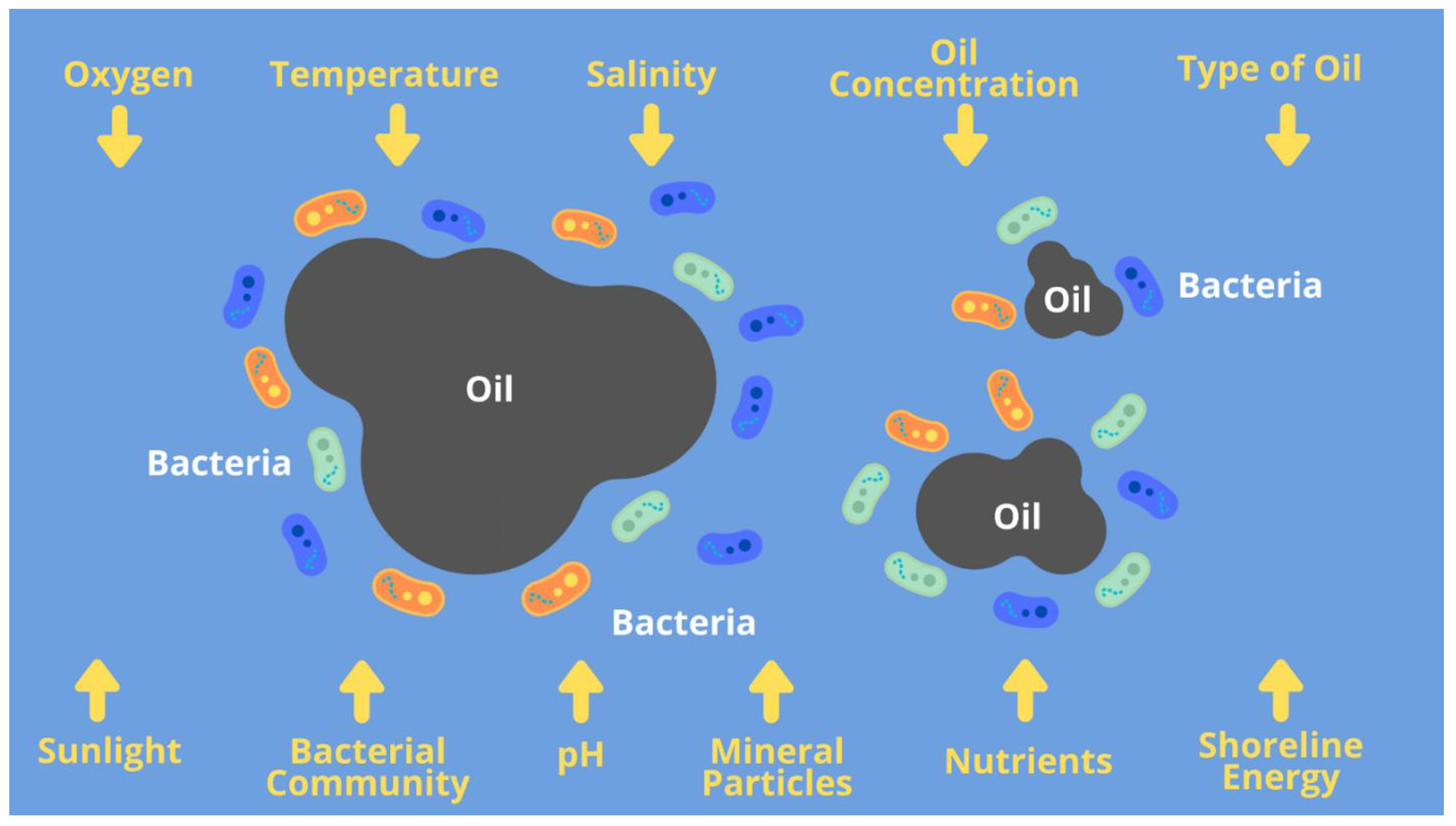
2. Factors Affecting Biodegradation of Oil Residues in Marine Environments
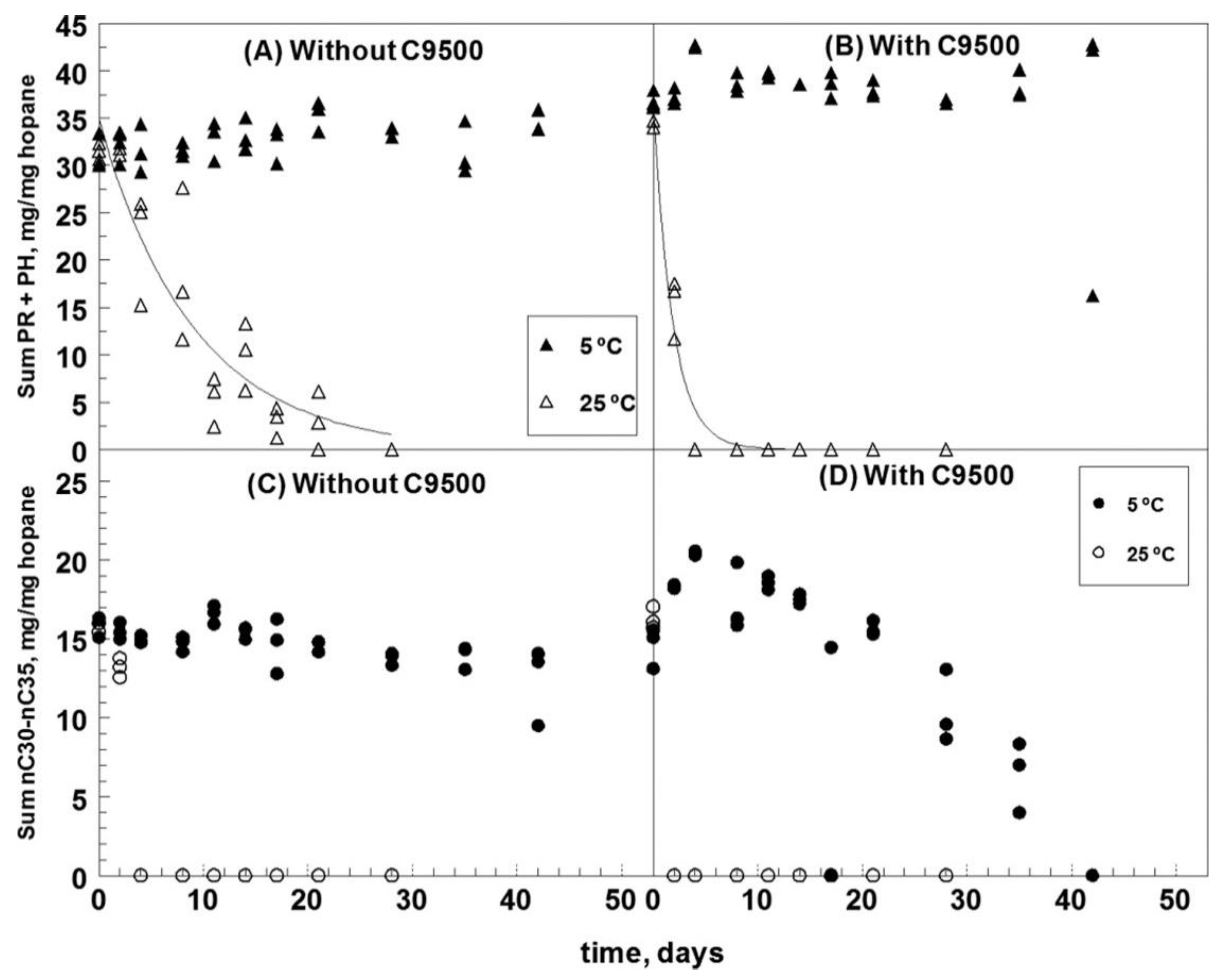
2.1. Temperature
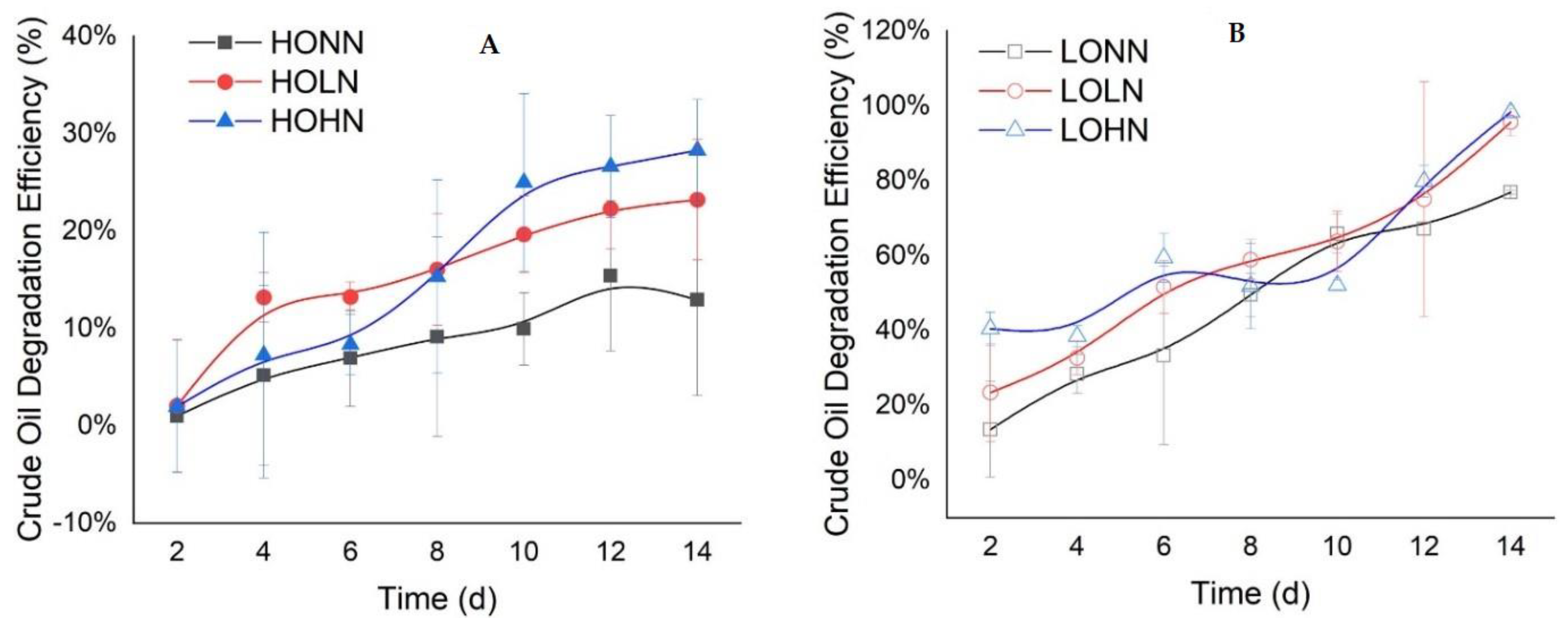
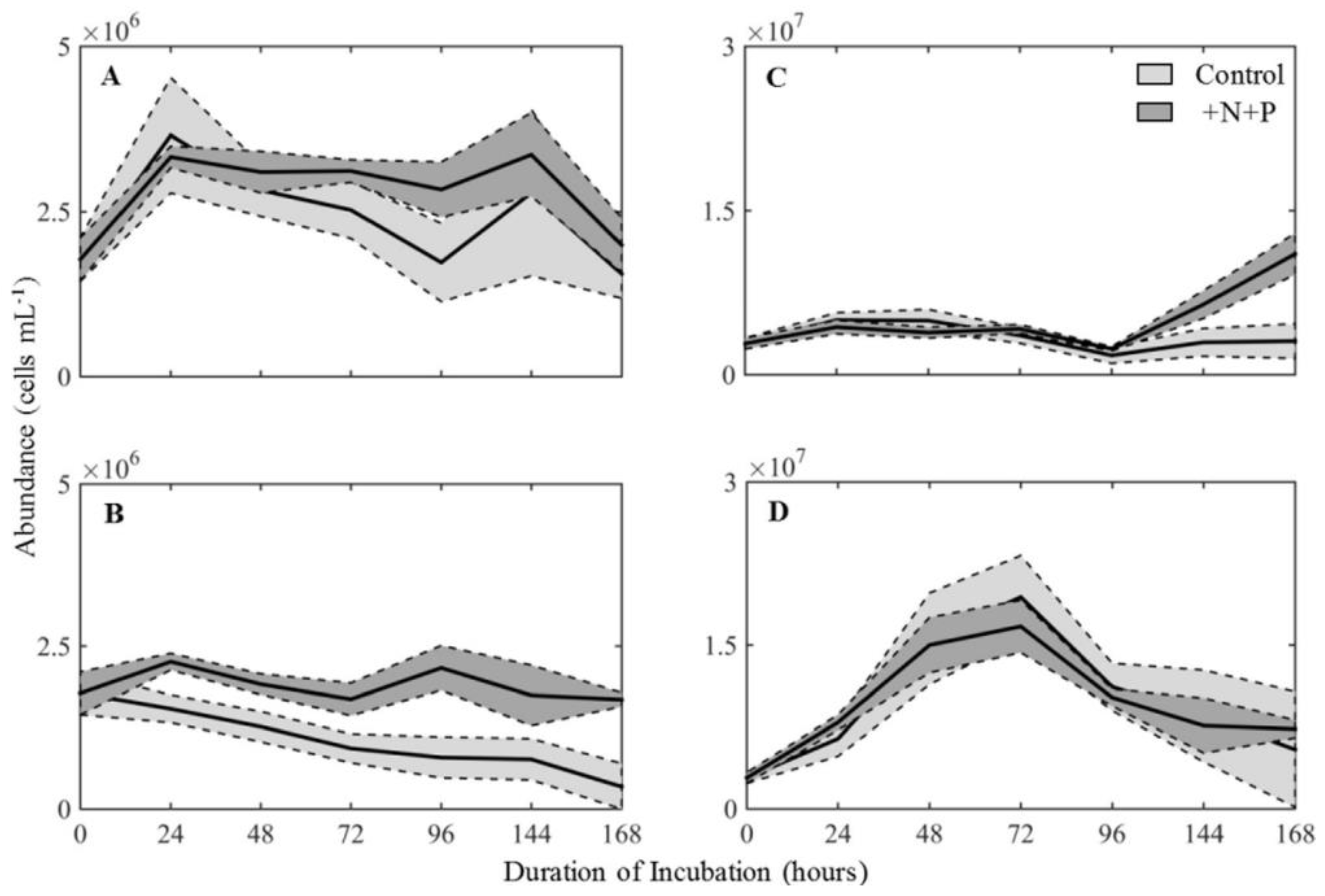
2.2. Nutrients
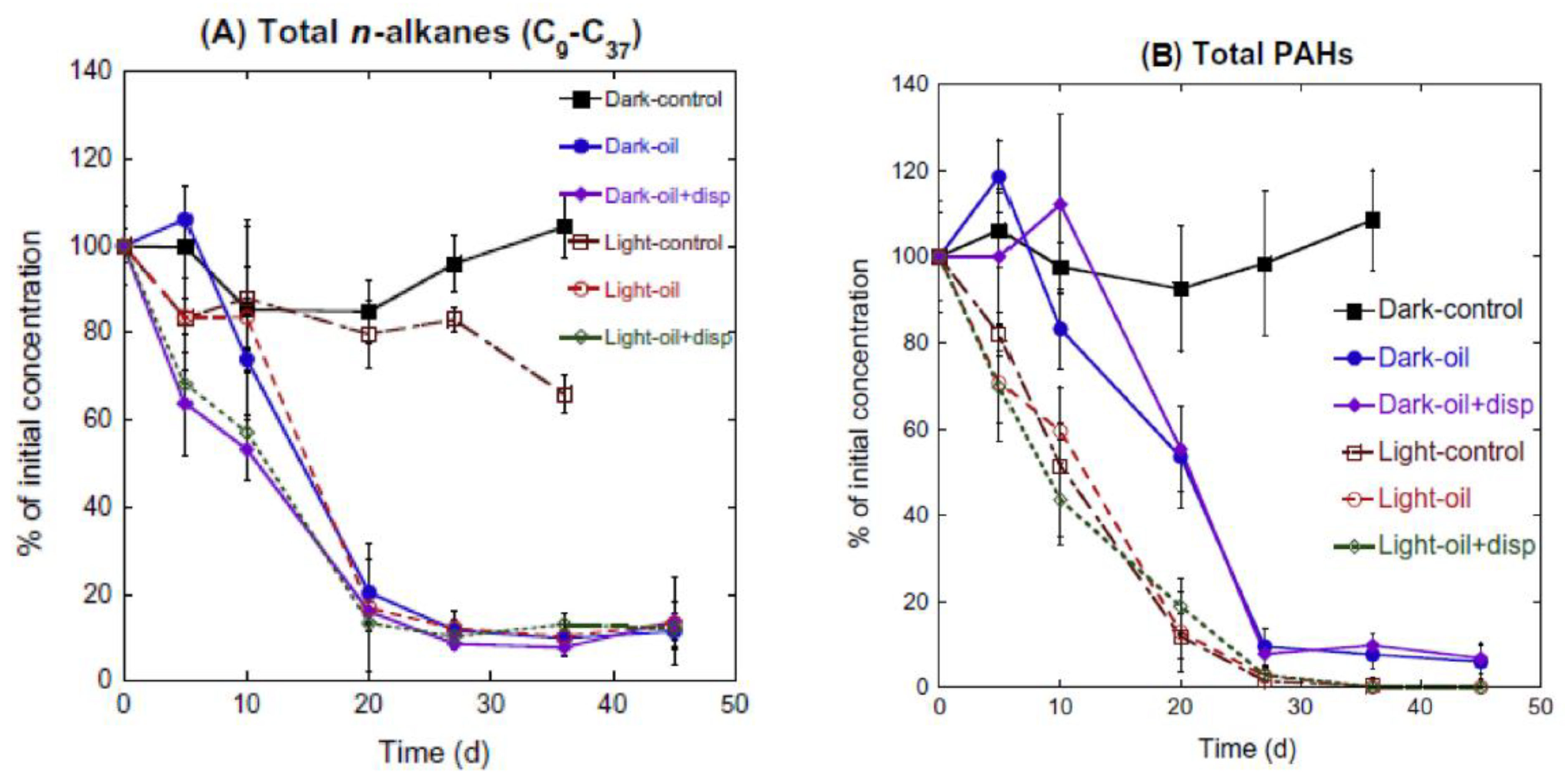
2.3. Sunlight
2.4. Oxygen
2.5. Type of Spilled-Oil
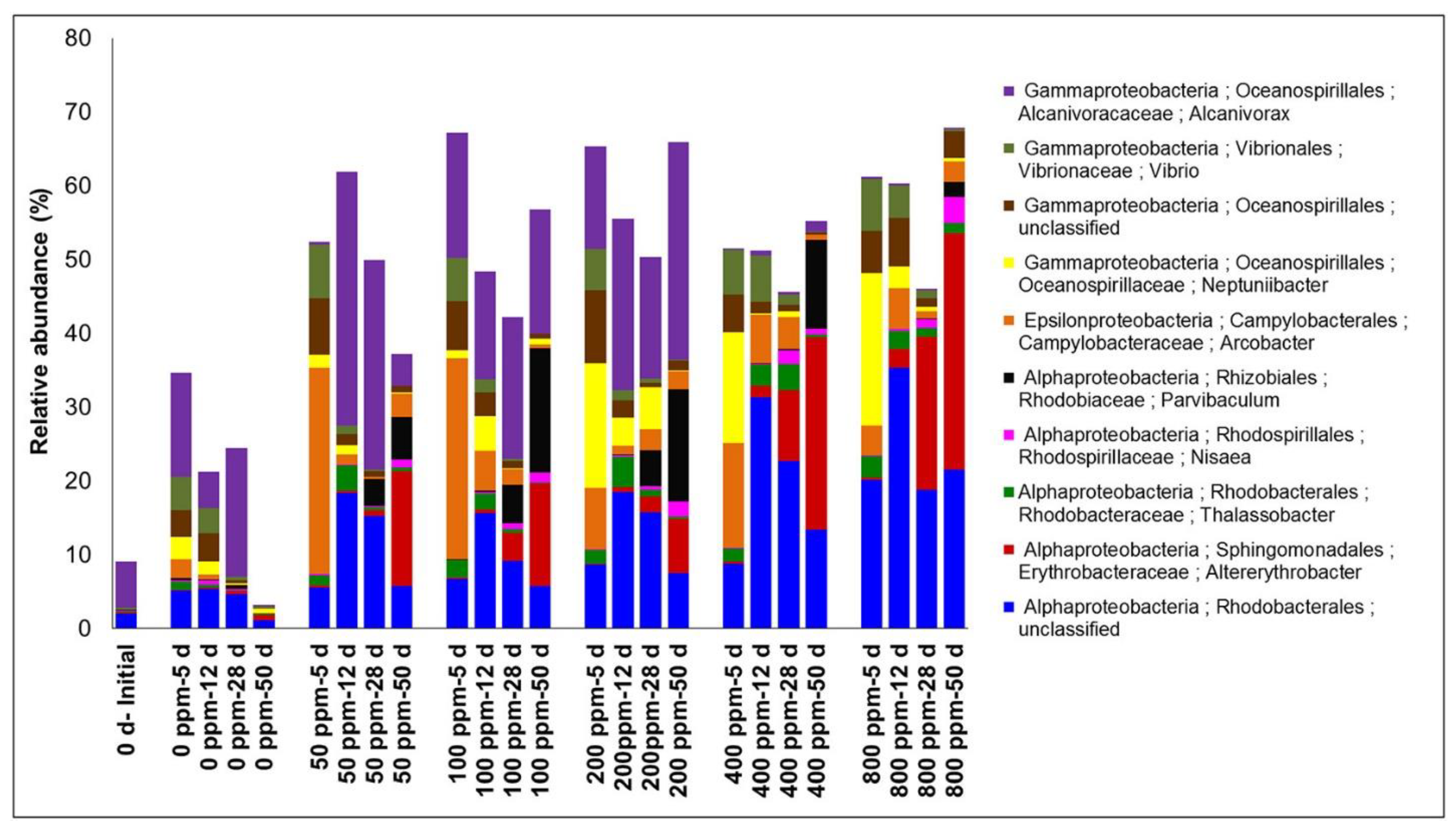
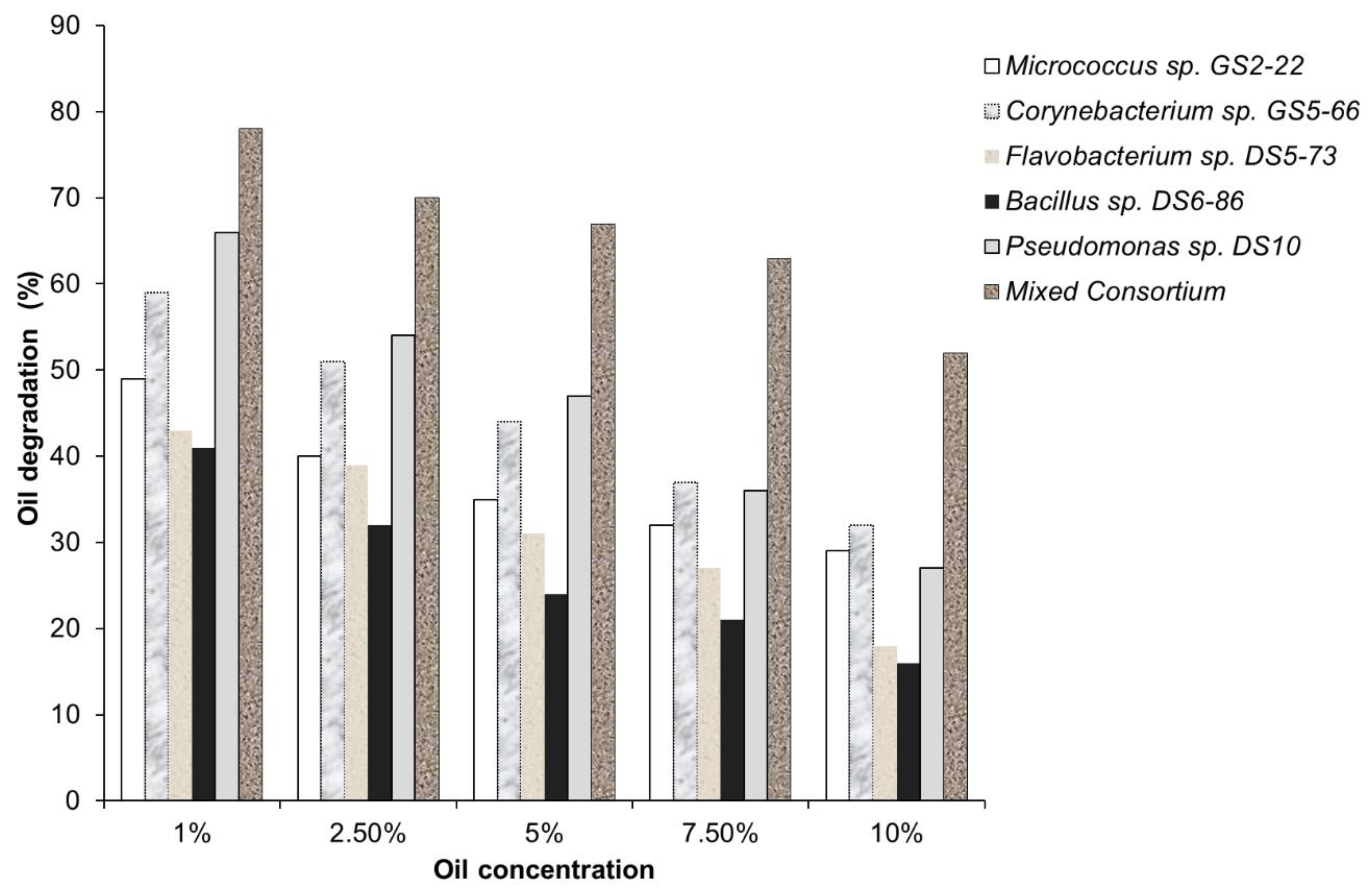
2.6. Oil Concentration
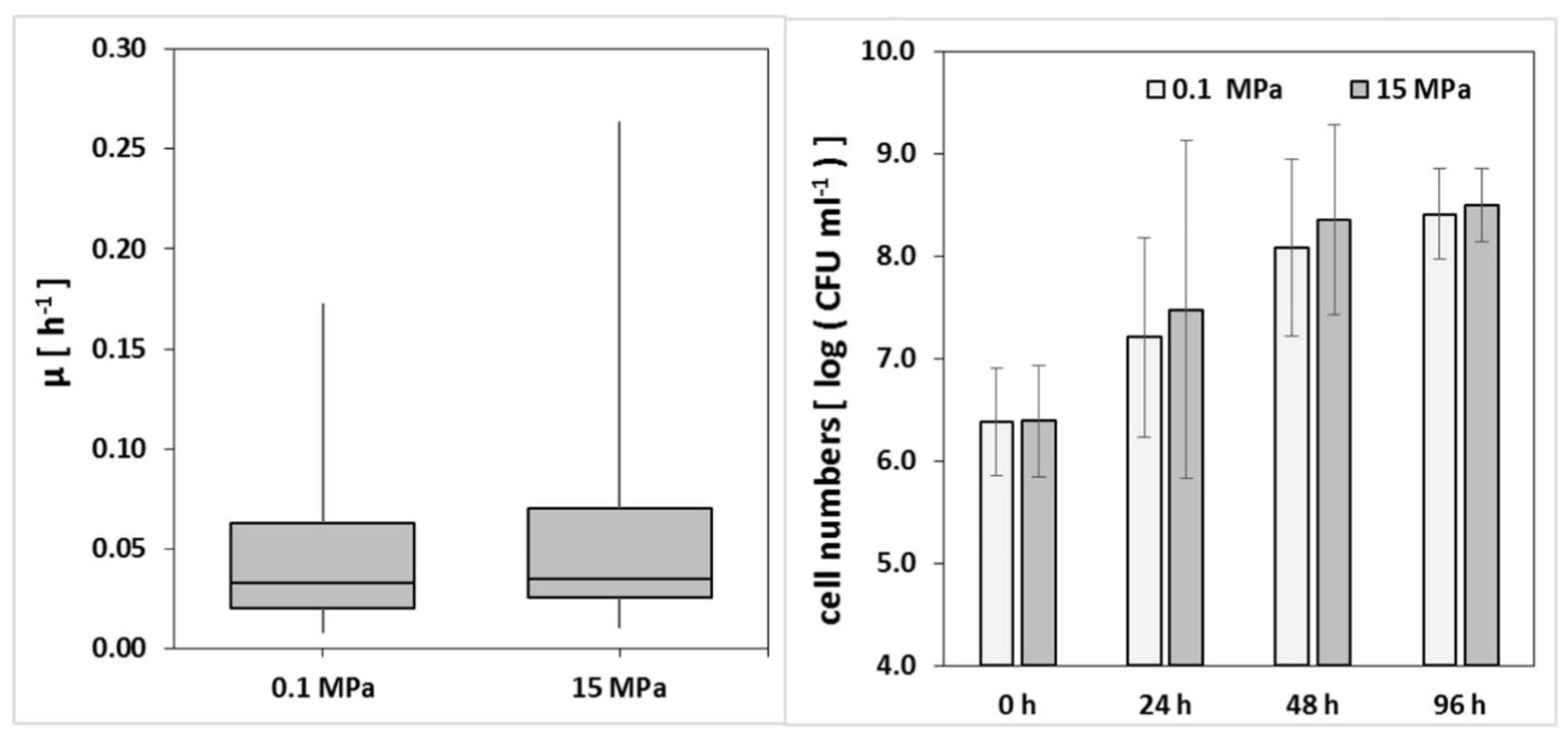
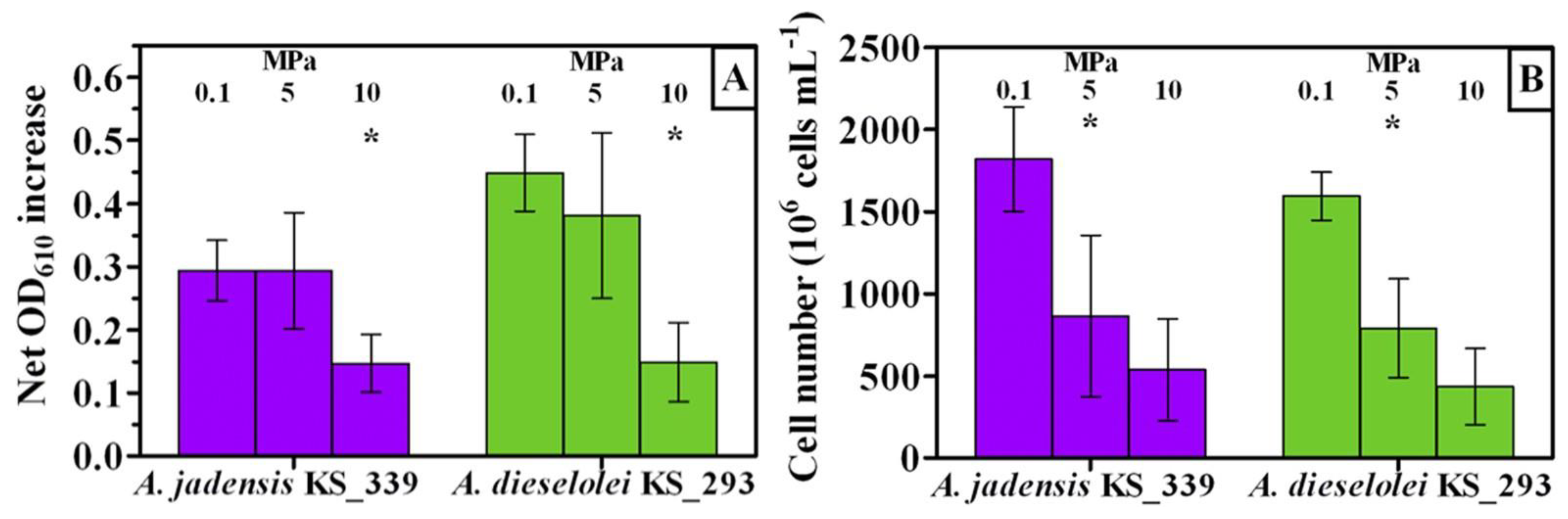
2.7. Pressure
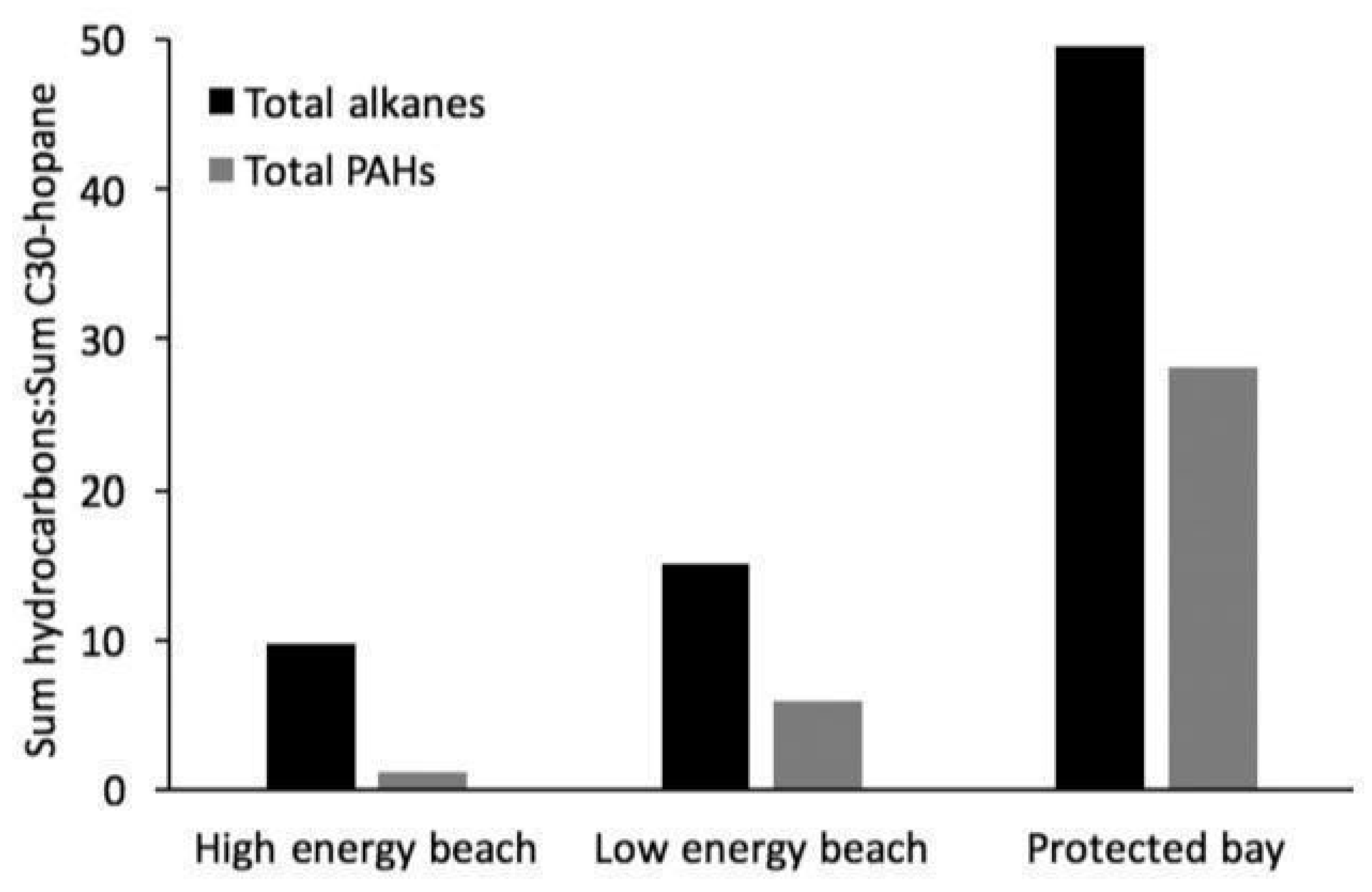
2.8. Shoreline Energy
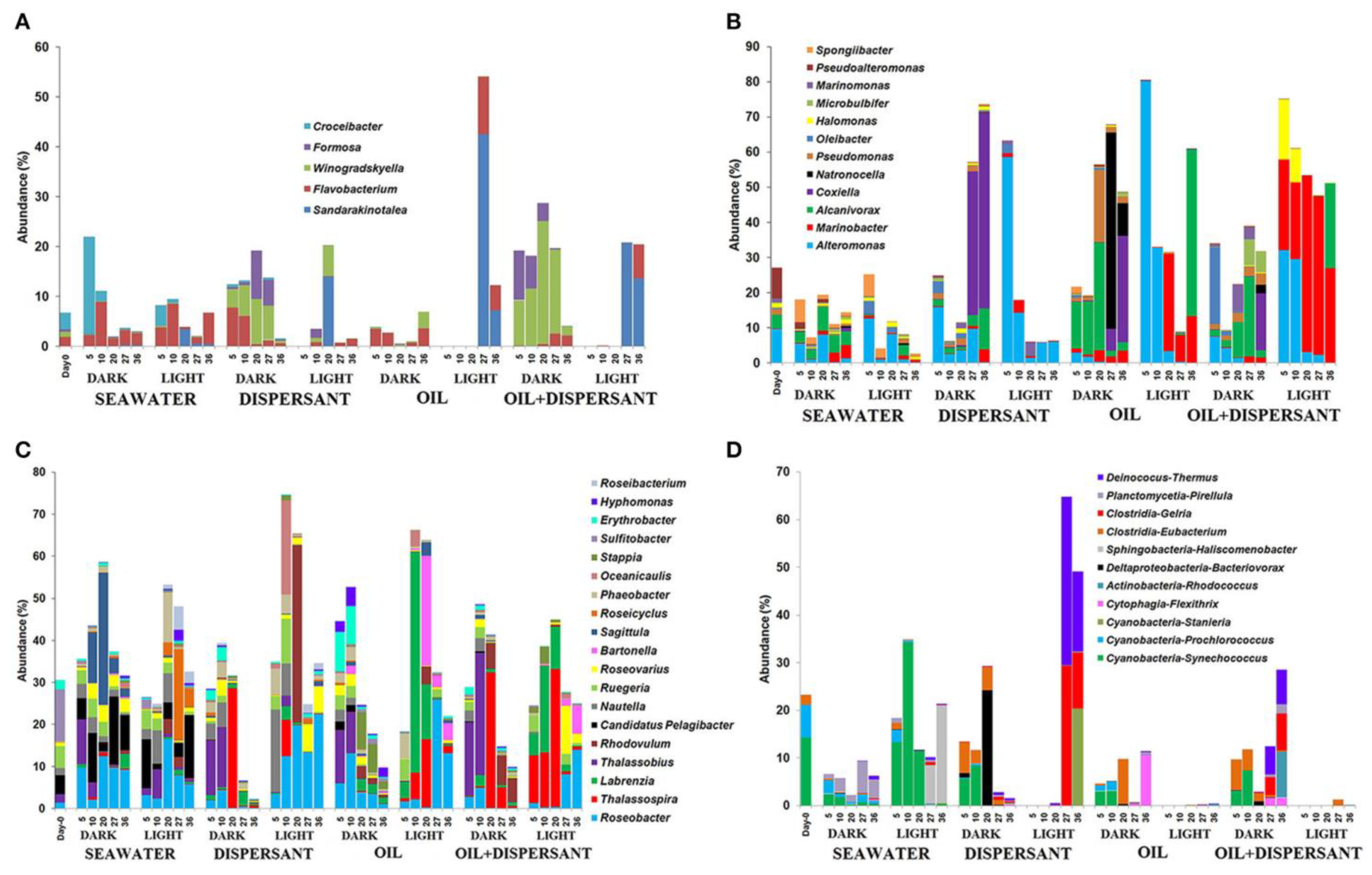
2.9. Bacterial Community
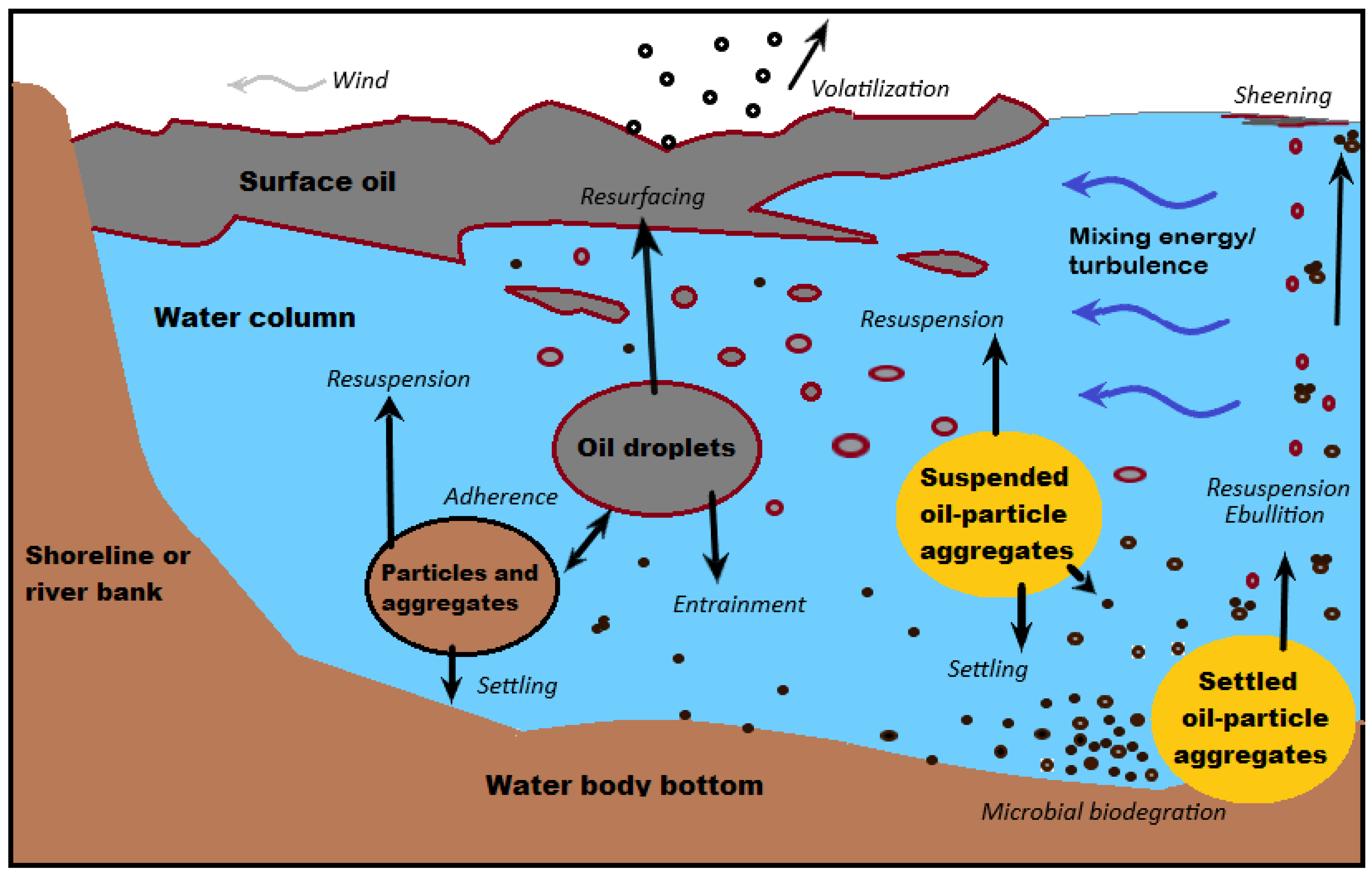
2.10. Mineral Particles
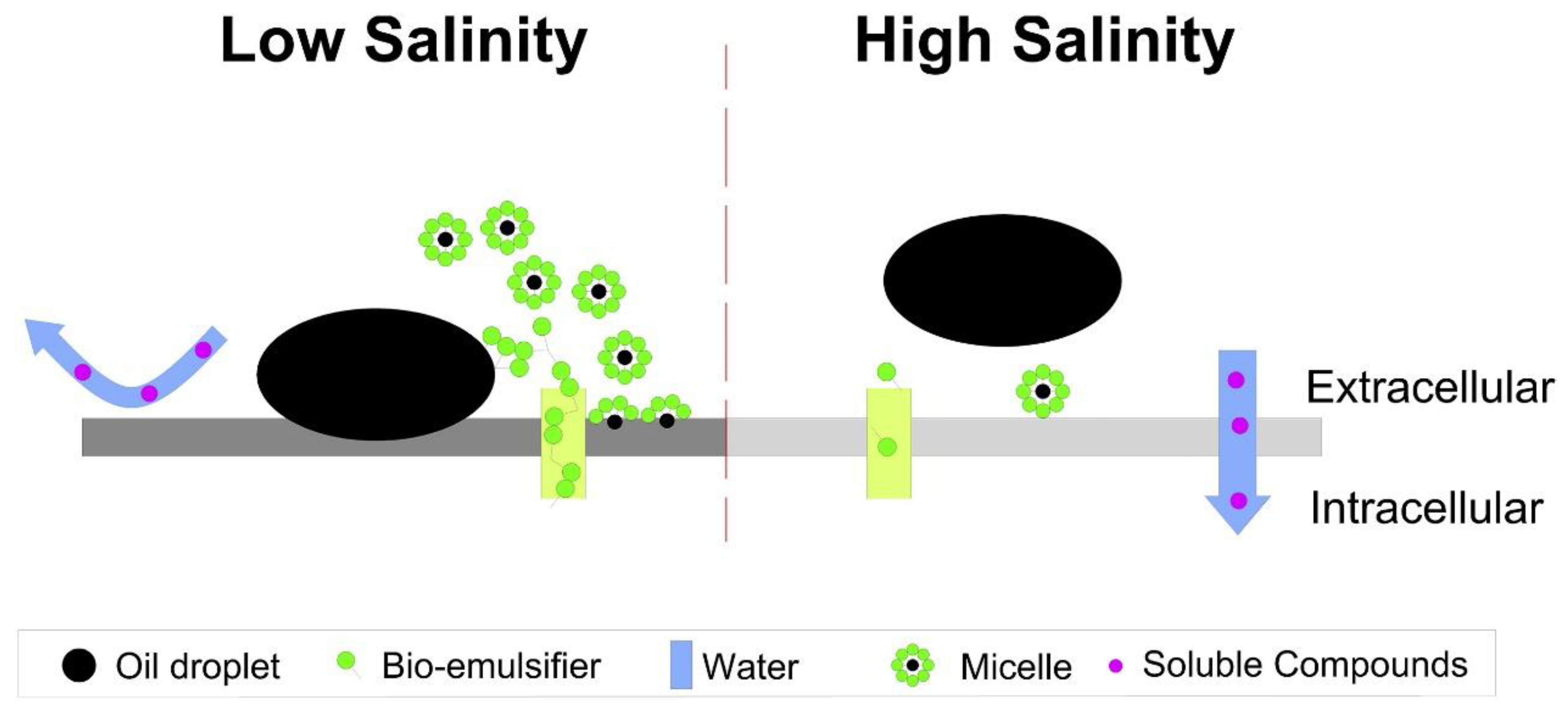
2.11. Salinity
2.12. pH
3. Conclusions
4. Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saadoun, I.M. Impact of oil spills on marine life. In Emerging Pollutants in the Environment-Current and Further Implications; BoD–Books on Demand: Norderstedt, Germany, 2015; pp. 75–104. [Google Scholar]
- Murphy, D.; Gemmell, B.; Vaccari, L.; Li, C.; Bacosa, H.; Evans, M.; Gemmell, C.; Harvey, T.; Jalali, M.; Niepa, T.H. An in-depth survey of the oil spill literature since 1968: Long term trends and changes since Deepwater Horizon. Mar. Pollut. Bull. 2016, 113, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Reddy, C.M.; Arey, J.S.; Seewald, J.S.; Sylva, S.P.; Lemkau, K.L.; Nelson, R.K.; Carmichael, C.A.; McIntyre, C.P.; Fenwick, J.; Ventura, G.T. Composition and fate of gas and oil released to the water column during the Deepwater Horizon oil spill. Proc. Natl. Acad. Sci. USA 2012, 109, 20229–20234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crone, T.J.; Tolstoy, M. Magnitude of the 2010 Gulf of Mexico oil leak. Science 2010, 330, 634. [Google Scholar] [CrossRef] [Green Version]
- Bacosa, H.P.; Evans, M.M.; Wang, Q.; Liu, Z. Assessing the role of environmental conditions on the degradation of oil following the deepwater horizon oil spill. In Oil Spill Environmental Forensics Case Studies; Elsevier: Amsterdam, The Netherlands, 2018; pp. 617–637. [Google Scholar]
- Goldstein, B.D.; Osofsky, H.J.; Lichtveld, M.Y. The Gulf oil spill. N. Engl. J. Med. 2011, 364, 1334–1348. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Bao, B.; Li, Y.; Liu, M.; Zhu, B.; Mu, J.; Chen, Z. Effects of marine oil pollution on microbial diversity in coastal waters and stimulating indigenous microorganism bioremediation with nutrients. Reg. Stud. Mar. Sci. 2020, 39, 101395. [Google Scholar] [CrossRef]
- Hassanshahian, M.; Cappello, S. Crude oil biodegradation in the marine environments. Biodegrad.-Eng. Technol. 2013, 5, 102–135. [Google Scholar]
- Park, B.S.; Erdner, D.L.; Bacosa, H.P.; Liu, Z.; Buskey, E.J. Potential effects of bacterial communities on the formation of blooms of the harmful dinoflagellate Prorocentrum after the 2014 Texas City “Y” oil spill (USA). Harmful Algae 2020, 95, 101802. [Google Scholar] [CrossRef]
- Kingston, P.F. Long-term environmental impact of oil spills. Spill Sci. Technol. Bull. 2002, 7, 53–61. [Google Scholar] [CrossRef]
- Fingas, M. Oil Spill Science and Technology; Gulf Professional Publishing: Oxford, UK, 2016. [Google Scholar]
- Kujawinski, E.B.; Kido Soule, M.C.; Valentine, D.L.; Boysen, A.K.; Longnecker, K.; Redmond, M.C. Fate of dispersants associated with the Deepwater Horizon oil spill. Environ. Sci. Technol. 2011, 45, 1298–1306. [Google Scholar] [CrossRef]
- Abbasian, F.; Lockington, R.; Mallavarapu, M.; Naidu, R. A comprehensive review of aliphatic hydrocarbon biodegradation by bacteria. Appl. Biochem. Biotechnol. 2015, 176, 670–699. [Google Scholar] [CrossRef] [PubMed]
- Overton, E.B.; Wetzel, D.L.; Wickliffe, J.K.; Adhikari, P.L. Spilled Oil Composition and the Natural Carbon Cycle: The True Drivers of Environmental Fate and Effects of Oil Spills. In Scenarios and Responses to Future Deep Oil Spills; Springer: Berlin/Heidelberg, Germany, 2020; pp. 33–56. [Google Scholar]
- Das, N.; Chandran, P. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacosa, H.; Suto, K.; Inoue, C. Preferential degradation of aromatic hydrocarbons in kerosene by a microbial consortium. Int. Biodeterior. Biodegrad. 2010, 64, 702–710. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Suto, K.; Inoue, C. Degradation potential and microbial community structure of heavy oil-enriched microbial consortia from mangrove sediments in Okinawa, Japan. J. Environ. Sci. Health Part A 2013, 48, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Christova, N.; Kabaivanova, L.; Nacheva, L.; Petrov, P.; Stoineva, I. Biodegradation of crude oil hydrocarbons by a newly isolated biosurfactant producing strain. Biotechnol. Biotechnol. Equip. 2019, 33, 863–872. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Inoue, C. Polycyclic aromatic hydrocarbons (PAHs) biodegradation potential and diversity of microbial consortia enriched from tsunami sediments in Miyagi, Japan. J. Hazard. Mater. 2015, 283, 689–697. [Google Scholar] [CrossRef]
- Dominguez, J.J.A.; Bacosa, H.; Chien, M.-F.; Inoue, C. Enhanced degradation of polycyclic aromatic hydrocarbons (PAHs) in the rhizosphere of sudangrass (Sorghum × drummondii). Chemosphere 2019, 234, 789–795. [Google Scholar] [CrossRef]
- Keramea, P.; Spanoudaki, K.; Zodiatis, G.; Gikas, G.; Sylaios, G. Oil spill modeling: A critical review on current trends, perspectives, and challenges. J. Mar. Sci. Eng. 2021, 9, 181. [Google Scholar] [CrossRef]
- Bretherton, L.; Hillhouse, J.; Bacosa, H.; Setta, S.; Genzer, J.; Kamalanathan, M.; Finkel, Z.V.; Quigg, A. Growth dynamics and domoic acid production of Pseudo-nitzschia sp. in response to oil and dispersant exposure. Harmful Algae 2019, 86, 55–63. [Google Scholar] [CrossRef]
- Kamalanathan, M.; Chiu, M.-H.; Bacosa, H.; Schwehr, K.; Tsai, S.-M.; Doyle, S.; Yard, A.; Mapes, S.; Vasequez, C.; Bretherton, L.; et al. Role of polysaccharides in diatom Thalassiosira pseudonana and its associated bacteria in hydrocarbon presence. Plant Physiol. 2019, 180, 1898–1911. [Google Scholar] [CrossRef] [Green Version]
- Severin, T.; Bacosa, H.; Sato, A.; Erdner, D. Dynamics of Heterocapsa sp. and the associated attached and free-living bacteria under the influence of dispersed and undispersed crude oil. Lett. Appl. Microbiol. 2016, 63, 419–425. [Google Scholar] [CrossRef] [PubMed]
- McGenity, T.J.; Folwell, B.D.; McKew, B.A.; Sanni, G.O. Marine crude-oil biodegradation: A central role for interspecies interactions. Aquat. Biosyst. 2012, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacosa, H.P.; Thyng, K.M.; Plunkett, S.; Erdner, D.L.; Liu, Z. The tarballs on Texas beaches following the 2014 Texas City “Y” Spill: Modeling, chemical, and microbiological studies. Mar. Pollut. Bull. 2016, 109, 236–244. [Google Scholar] [CrossRef]
- Hazen, T.C.; Prince, R.C.; Mahmoudi, N. Marine Oil Biodegradation; American Chemical Society (ACS) Publications: Washington, DC, USA, 2016. [Google Scholar]
- Steichen, J.L.; Labonté, J.M.; Windham, R.; Hala, D.; Kaiser, K.; Setta, S.; Faulkner, P.C.; Bacosa, H.; Yan, G.; Kamalanathan, M. Microbial, physical, and chemical changes in Galveston Bay following an extreme flooding event, Hurricane Harvey. Front. Mar. Sci. 2020, 7, 186. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Steichen, J.; Kamalanathan, M.; Windham, R.; Lubguban, A.; Labonté, J.M.; Kaiser, K.; Hala, D.; Santschi, P.H.; Quigg, A. Polycyclic aromatic hydrocarbons (PAHs) and putative PAH-degrading bacteria in Galveston Bay, TX (USA), following Hurricane Harvey (2017). Environ. Sci. Pollut. Res. 2020, 27, 34987–34999. [Google Scholar] [CrossRef] [PubMed]
- Abed, R.M.; Köster, J. The direct role of aerobic heterotrophic bacteria associated with cyanobacteria in the degradation of oil compounds. Int. Biodeterior. Biodegrad. 2005, 55, 29–37. [Google Scholar] [CrossRef]
- Williams, A.K.; Bacosa, H.P.; Quigg, A. The impact of dissolved inorganic nitrogen and phosphorous on responses of microbial plankton to the Texas City “Y” oil spill in Galveston Bay, Texas (USA). Mar. Pollut. Bull. 2017, 121, 32–44. [Google Scholar] [CrossRef]
- Gemmell, B.J.; Bacosa, H.P.; Dickey, B.O.; Gemmell, C.G.; Alqasemi, L.R.; Buskey, E.J. Rapid alterations to marine microbiota communities following an oil spill. Ecotoxicology 2018, 27, 505–516. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Erdner, D.L.; Liu, Z. Differentiating the roles of photooxidation and biodegradation in the weathering of Light Louisiana Sweet crude oil in surface water from the Deepwater Horizon site. Mar. Pollut. Bull. 2015, 95, 265–272. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Erdner, D.L.; Rosenheim, B.E.; Shetty, P.; Seitz, K.W.; Baker, B.J.; Liu, Z. Hydrocarbon degradation and response of seafloor sediment bacterial community in the northern Gulf of Mexico to light Louisiana sweet crude oil. ISME J. 2018, 12, 2532–2543. [Google Scholar] [CrossRef]
- Chikere, C.B.; Okpokwasili, G.C.; Chikere, B.O. Monitoring of microbial hydrocarbon remediation in the soil. 3 Biotech 2011, 1, 117–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Transportation Research Board and National Research Council. Oil in the Sea III: Inputs, Fates, and Effects; The National Academies Press: Washington, DC, USA, 2003. [Google Scholar]
- Campo, P.; Venosa, A.D.; Suidan, M.T. Biodegradability of Corexit 9500 and dispersed South Louisiana crude oil at 5 and 25 °C. Environ. Sci. Technol. 2013, 47, 1960–1967. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Liu, H.; Bao, M.; Sun, P. Microbial community structure shifts are associated with temperature, dispersants and nutrients in crude oil-contaminated seawaters. Mar. Pollut. Bull. 2016, 111, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bacosa, H.P.; Liu, Z. Potential environmental factors affecting oil-degrading bacterial populations in deep and surface waters of the northern Gulf of Mexico. Front. Microbiol. 2017, 7, 2131. [Google Scholar] [CrossRef] [Green Version]
- Bacosa, H.P.; Kang, A.; Lu, K.; Liu, Z. Initial oil concentration affects hydrocarbon biodegradation rates and bacterial community composition in seawater. Mar. Pollut. Bull. 2021, 162, 111867. [Google Scholar] [CrossRef]
- Ostroumov, V.; Siegert, C. Exobiological aspects of mass transfer in microzones of permafrost deposits. Adv. Space Res. 1996, 18, 79–86. [Google Scholar] [CrossRef]
- Aislabie, J.; Saul, D.J.; Foght, J.M. Bioremediation of hydrocarbon-contaminated polar soils. Extremophiles 2006, 10, 171–179. [Google Scholar] [CrossRef]
- AUNG, M.T.M.; LI, Q.; TAKAHASHI, S.; UTSUMI, M. Effect of Temperature on Hydrocarbon Bioremediation in Simulated Petroleum-Polluted Seawater Collected from Tokyo Bay. Jpn. J. Water Treat. Biol. 2018, 54, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Siron, R.; Pelletier, E.; Brochu, C. Environmental factors influencing the biodegradation of petroleum hydrocarbons in cold seawater. Arch. Environ. Contam. Toxicol. 1995, 28, 406–416. [Google Scholar] [CrossRef]
- Redmond, M.C.; Valentine, D.L. Natural gas and temperature structured a microbial community response to the Deepwater Horizon oil spill. Proc. Natl. Acad. Sci. USA 2012, 109, 20292–20297. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Liu, J. Evaluating bacterial community structures in oil collected from the sea surface and sediment in the northern Gulf of Mexico after the Deepwater Horizon oil spill. Microbiol. Open 2013, 2, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Coulon, F.; Pelletier, E.; Gourhant, L.; Delille, D. Effects of nutrient and temperature on degradation of petroleum hydrocarbons in contaminated sub-Antarctic soil. Chemosphere 2005, 58, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Kirchman, D.L. Processes in Microbial Ecology; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Vázquez-Domínguez, E.; Vaqué, D.; Gasol, J.M. Temperature effects on the heterotrophic bacteria, heterotrophic nanoflagellates, and microbial top predators of the NW Mediterranean. Aquat. Microb. Ecol. 2012, 67, 107–121. [Google Scholar] [CrossRef]
- Morán, X.A.G.; Alonso-Sáez, L.; Nogueira, E.; Ducklow, H.W.; González, N.; López-Urrutia, Á.; Díaz-Pérez, L.; Calvo-Díaz, A.; Arandia-Gorostidi, N.; Huete-Stauffer, T.M. More, smaller bacteria in response to ocean’s warming? Proc. R. Soc. B 2015, 282, 20150371. [Google Scholar] [CrossRef] [Green Version]
- Semboung Lang, F.; Tarayre, C.; Destain, J.; Delvigne, F.; Druart, P.; Ongena, M.; Thonart, P. The effect of nutrients on the degradation of hydrocarbons in mangrove ecosystems by microorganisms. Int. J. Environ. Res. 2016, 10, 583–592. [Google Scholar]
- Vyas, T.K.; Dave, B. Effect of addition of nitrogen, phosphorus and potassium fertilizers on biodegradation of crude oil by marine bacteria. IJMS 2010, 39, 143–150. [Google Scholar]
- Harrison, S.J. Lessons from the Taylor Energy Oil Spill: History, Seasonality, and Nutrient Limitation; University of Georgia: Athens, GA, USA, 2017. [Google Scholar]
- Edwards, B.R.; Reddy, C.M.; Camilli, R.; Carmichael, C.A.; Longnecker, K.; Van Mooy, B.A. Rapid microbial respiration of oil from the Deepwater Horizon spill in offshore surface waters of the Gulf of Mexico. Environ. Res. Lett. 2011, 6, 035301. [Google Scholar] [CrossRef]
- Dubinsky, E.A.; Conrad, M.E.; Chakraborty, R.; Bill, M.; Borglin, S.E.; Hollibaugh, J.T.; Mason, O.U.; Piceno, M.Y.; Reid, F.C.; Stringfellow, W.T. Succession of hydrocarbon-degrading bacteria in the aftermath of the Deepwater Horizon oil spill in the Gulf of Mexico. Environ. Sci. Technol. 2013, 47, 10860–10867. [Google Scholar] [CrossRef]
- Elango, V.; Urbano, M.; Lemelle, K.R.; Pardue, J.H. Biodegradation of MC252 oil in oil: Sand aggregates in a coastal headland beach environment. Front. Microbiol. 2014, 5, 161. [Google Scholar] [CrossRef]
- Alabresm, A.; Chen, Y.P.; Decho, A.W.; Lead, J. A novel method for the synergistic remediation of oil-water mixtures using nanoparticles and oil-degrading bacteria. Sci. Total Environ. 2018, 630, 1292–1297. [Google Scholar] [CrossRef]
- Xu, J.; Du, J.; Li, L.; Zhang, Q.; Chen, Z. Fast-stimulating bioremediation of macro crude oil in soils using matching Fenton pre-oxidation. Chemosphere 2020, 252, 126622. [Google Scholar] [CrossRef] [PubMed]
- Zahid, M.R.; Khamaruddin, P.F.M.; Him, N.R.N.; Othman, N.H.; Yahya, E. Effect of Organic Nutrient Addition to the Biodegradation of Hydrocarbon Contaminated Marine Sediment in Malaysia. In Key Engineering Materials; Trans Tech Publications Ltd.: Kepellwag, Switzerland, 2019; pp. 74–83. [Google Scholar]
- Chaillan, F.; Le Flèche, A.; Bury, E.; Phantavong, Y.-h.; Grimont, P.; Saliot, A.; Oudot, J. Identification and biodegradation potential of tropical aerobic hydrocarbon-degrading microorganisms. Res. Microbiol. 2004, 155, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Oudot, J.; Merlin, F.; Pinvidic, P. Weathering rates of oil components in a bioremediation experiment in estuarine sediments. Mar. Environ. Res. 1998, 45, 113–125. [Google Scholar] [CrossRef]
- Chaineau, C.; Rougeux, G.; Yepremian, C.; Oudot, J. Effects of nutrient concentration on the biodegradation of crude oil and associated microbial populations in the soil. Soil Biol. Biochem. 2005, 37, 1490–1497. [Google Scholar] [CrossRef]
- Thavasi, R.; Jayalakshmi, S.; Banat, I.M. Application of biosurfactant produced from peanut oil cake by Lactobacillus delbrueckii in biodegradation of crude oil. Bioresour. Technol. 2011, 102, 3366–3372. [Google Scholar] [CrossRef]
- Farag, S.; Soliman, N.A.; Abdel-Fattah, Y.R. Statistical optimization of crude oil bio-degradation by a local marine bacterium isolate Pseudomonas sp. sp48. J. Genet. Eng. Biotechnol. 2018, 16, 409–420. [Google Scholar] [CrossRef]
- Souza, E.C.; Vessoni-Penna, T.C.; de Souza Oliveira, R.P. Biosurfactant-enhanced hydrocarbon bioremediation: An overview. Int. Biodeterior. Biodegrad. 2014, 89, 88–94. [Google Scholar] [CrossRef]
- Waikhom, D.; Ngasotter, S.; Soniya Devi, L.; Devi, S.; Singh, A.S. Role of Microbes in Petroleum Hydrocarbon Degradation in the Aquatic Environment: A Review. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 2890–2903. [Google Scholar] [CrossRef]
- Brakstad, O.G.; Lofthus, S.; Ribicic, D.; Netzer, R. Biodegradation of petroleum oil in cold marine environments. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Berlin/Heidelberg, Germany, 2017; pp. 613–644. [Google Scholar]
- Wurtsbaugh, W.A.; Paerl, H.W.; Dodds, W.K. Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum. Wiley Interdiscip. Rev. 2019, 6, e1373. [Google Scholar] [CrossRef]
- Huettel, M.; Berg, P.; Kostka, J.E. Benthic exchange and biogeochemical cycling in permeable sediments. Annu. Rev. Mar. Sci. 2014, 6, 23–51. [Google Scholar] [CrossRef]
- Joye, S.B.; Teske, A.P.; Kostka, J.E. Microbial dynamics following the Macondo oil well blowout across Gulf of Mexico environments. Bioscience 2014, 64, 766–777. [Google Scholar] [CrossRef] [Green Version]
- Hazen, T.C.; Dubinsky, E.A.; DeSantis, T.Z.; Andersen, G.L.; Piceno, Y.M.; Singh, N.; Jansson, J.K.; Probst, A.; Borglin, S.E.; Fortney, J.L. Deep-sea oil plume enriches indigenous oil-degrading bacteria. Science 2010, 330, 204–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walworth, J.; Pond, A.; Snape, I.; Rayner, J.; Ferguson, S.; Harvey, P. Nitrogen requirements for maximizing petroleum bioremediation in a sub-Antarctic soil. Cold Reg. Sci. Technol. 2007, 48, 84–91. [Google Scholar] [CrossRef]
- Aeppli, C.; Carmichael, C.A.; Nelson, R.K.; Lemkau, K.L.; Graham, W.M.; Redmond, M.C.; Valentine, D.L.; Reddy, C.M. Oil weathering after the Deepwater Horizon disaster led to the formation of oxygenated residues. Environ. Sci. Technol. 2012, 46, 8799–8807. [Google Scholar] [CrossRef] [PubMed]
- Tarr, M.A.; Zito, P.; Overton, E.B.; Olson, G.M.; Adhikari, P.L.; Reddy, C.M. Weathering of oil spilled in the marine environment. Oceanography 2016, 29, 126–135. [Google Scholar] [CrossRef] [Green Version]
- King, S.M.; Leaf, P.A.; Olson, A.C.; Ray, P.Z.; Tarr, M.A. Photolytic and photocatalytic degradation of surface oil from the Deepwater Horizon spill. Chemosphere 2014, 95, 415–422. [Google Scholar] [CrossRef]
- D’Auria, M.; Emanuele, L.; Racioppi, R.; Velluzzi, V. Photochemical degradation of crude oil: Comparison between direct irradiation, photocatalysis, and photocatalysis on zeolite. J. Hazard. Mater. 2009, 164, 32–38. [Google Scholar] [CrossRef]
- Ward, C.P.; Overton, E.B. How the 2010 Deepwater Horizon spill reshaped our understanding of crude oil photochemical weathering at sea: A past, present, and future perspective. Environ. Sci. 2020, 22, 1125–1138. [Google Scholar] [CrossRef]
- Ziolli, R.L.; Jardim, W.F. Photochemical transformations of water-soluble fraction (WSF) of crude oil in marine waters: A comparison between photolysis and accelerated degradation with TiO2 using GC–MS and UVF. J. Photochem. Photobiol. A 2003, 155, 243–252. [Google Scholar] [CrossRef]
- Griffiths, M.T.; Da Campo, R.; O’Connor, P.B.; Barrow, M.P. Throwing light on petroleum: Simulated exposure of crude oil to sunlight and characterization using atmospheric pressure photoionization Fourier transform ion cyclotron resonance mass spectrometry. Anal. Chem. 2014, 86, 527–534. [Google Scholar] [CrossRef]
- Shankar, R.; Shim, W.J.; An, J.G.; Yim, U.H. A practical review on photooxidation of crude oil: Laboratory lamp setup and factors affecting it. Water Res. 2015, 68, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Hall, G.J.; Frysinger, G.S.; Aeppli, C.; Carmichael, C.A.; Gros, J.; Lemkau, K.L.; Nelson, R.K.; Reddy, C.M. Oxygenated weathering products of Deepwater Horizon oil come from surprising precursors. Mar. Pollut. Bull. 2013, 75, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Radović, J.R.; Aeppli, C.; Nelson, R.K.; Jimenez, N.; Reddy, C.M.; Bayona, J.M.; Albaiges, J. Assessment of photochemical processes in marine oil spill fingerprinting. Mar. Pollut. Bull. 2014, 79, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Dutta, T.K.; Harayama, S. Fate of crude oil by the combination of photooxidation and biodegradation. Environ. Sci. Technol. 2000, 34, 1500–1505. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Liu, Z.; Erdner, D.L. Natural sunlight shapes crude oil-degrading bacterial communities in Northern Gulf of Mexico surface waters. Front. Microbiol. 2015, 6, 1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, P.Z.; Tarr, M.A. Solar production of singlet oxygen from crude oil films on water. J. Photochem. Photobiol. A 2014, 286, 22–28. [Google Scholar] [CrossRef]
- Ray, P.Z.; Tarr, M.A. Petroleum films exposed to sunlight produce hydroxyl radical. Chemosphere 2014, 103, 220–227. [Google Scholar] [CrossRef]
- Plata, D.L.; Sharpless, C.M.; Reddy, C.M. Photochemical degradation of polycyclic aromatic hydrocarbons in oil films. Environ. Sci. Technol. 2008, 42, 2432–2438. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Kamalanathan, M.; Chiu, M.-H.; Tsai, S.-M.; Sun, L.; Labonté, J.M.; Schwehr, K.A.; Hala, D.; Santschi, P.H.; Chin, W.-C. Extracellular polymeric substances (EPS) producing and oil degrading bacteria isolated from the northern Gulf of Mexico. PLoS ONE 2018, 13, e0208406. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Chiu, M.-H.; Xu, C.; Lin, P.; Schwehr, K.A.; Bacosa, H.; Kamalanathan, M.; Quigg, A.; Chin, W.-C.; Santschi, P.H. The effects of sunlight on the composition of exopolymeric substances and subsequent aggregate formation during oil spills. Mar. Chem. 2018, 203, 49–54. [Google Scholar] [CrossRef]
- Passow, U.; Ziervogel, K.; Asper, V.; Diercks, A. Marine snow formation in the aftermath of the Deepwater Horizon oil spill in the Gulf of Mexico. Environ. Res. Lett. 2012, 7, 035301. [Google Scholar] [CrossRef] [Green Version]
- Bacosa, H.P.; Kamalanathan, M.; Cullen, J.; Shi, D.; Xu, C.; Schwehr, K.A.; Hala, D.; Wade, T.L.; Knap, A.H.; Santschi, P.H. Marine snow aggregates are enriched in polycyclic aromatic hydrocarbons (PAHs) in oil contaminated waters: Insights from a mesocosm study. J. Mar. Sci. Eng. 2020, 8, 781. [Google Scholar] [CrossRef]
- Shiu, R.-F.; Chiu, M.-H.; Vazquez, C.I.; Tsai, Y.-Y.; Le, A.; Kagiri, A.; Xu, C.; Kamalanathan, M.; Bacosa, H.P.; Doyle, S.M. Protein to carbohydrate (P/C) ratio changes in microbial extracellular polymeric substances induced by oil and Corexit. Mar. Chem. 2020, 223, 103789. [Google Scholar] [CrossRef]
- Santschi, P.H.; Xu, C.; Schwehr, K.A.; Lin, P.; Sun, L.; Chin, W.-C.; Kamalanathan, M.; Bacosa, H.P.; Quigg, A. Can the protein/carbohydrate (P/C) ratio of exopolymeric substances (EPS) be used as a proxy for their ‘stickiness’ and aggregation propensity? Mar. Chem. 2020, 218, 103734. [Google Scholar] [CrossRef]
- Sun, L.; Xu, C.; Zhang, S.; Lin, P.; Schwehr, K.A.; Quigg, A.; Chiu, M.-H.; Chin, W.-C.; Santschi, P.H. Light-induced aggregation of microbial exopolymeric substances. Chemosphere 2017, 181, 675–681. [Google Scholar] [CrossRef]
- Venosa, A.D.; Zhu, X. Biodegradation of crude oil contaminating marine shorelines and freshwater wetlands. Spill Sci. Technol. Bull. 2003, 8, 163–178. [Google Scholar] [CrossRef]
- Meng, Q.; Wang, X.; Wang, X.; Shi, B.; Luo, X.; Zhang, L.; Lei, Y.; Jiang, C.; Liu, P. Gas geochemical evidences for biodegradation of shale gases in the Upper Triassic Yanchang Formation, Ordos Basin, China. Int. J. Coal Geol. 2017, 179, 139–152. [Google Scholar] [CrossRef]
- Sierra-Garcia, I.N.; de Oliveira, V.M. Microbial hydrocarbon degradation: Efforts to understand biodegradation in petroleum reservoirs. Biodegrad.-Eng. Technol. 2013, 10, 55920. [Google Scholar]
- McNally, D.L.; Mihelcic, J.R.; Lueking, D.R. Biodegradation of three-and four-ring polycyclic aromatic hydrocarbons under aerobic and denitrifying conditions. Environ. Sci. Technol. 1998, 32, 2633–2639. [Google Scholar] [CrossRef]
- Grishchenkov, V.; Townsend, R.; McDonald, T.; Autenrieth, R.; Bonner, J.; Boronin, A. Degradation of petroleum hydrocarbons by facultative anaerobic bacteria under aerobic and anaerobic conditions. Process Biochem. 2000, 35, 889–896. [Google Scholar] [CrossRef]
- Olajire, A.; Essien, J. Aerobic degradation of petroleum components by microbial consortia. J. Pet. Environ. Biotechnol. 2014, 5, 1. [Google Scholar] [CrossRef]
- Mille, G.; Mulyono, M.; El Jammal, T.; Bertrand, J.-C. Effects of oxygen on hydrocarbon degradation studies in vitro in surficial sediments. Estuar. Coast. Shelf Sci. 1988, 27, 283–295. [Google Scholar] [CrossRef]
- Kostka, J.E.; Joye, S.B.; Overholt, W.; Bubenheim, P.; Hackbusch, S.; Larter, S.R.; Liese, A.; Lincoln, S.A.; Marietou, A.; Müller, R. Biodegradation of petroleum hydrocarbons in the deep sea. In Deep Oil Spills; Springer: Berlin/Heidelberg, Germany, 2020; pp. 107–124. [Google Scholar]
- Nicholson, C.A.; Fathepure, B.Z. Biodegradation of benzene by halophilic and halotolerant bacteria under aerobic conditions. Appl. Environ. Microbiol. 2004, 70, 1222–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakimov, M.M.; Golyshin, P.N.; Lang, S.; Moore, E.R.; Abraham, W.-R.; Lünsdorf, H.; Timmis, K.N. Alcanivorax borkumensis gen. nov., sp. nov., a new, hydrocarbon-degrading and surfactant-producing marine bacterium. Int. J. Syst. Evol. Microbiol. 1998, 48, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Dyksterhouse, S.E.; Gray, J.P.; Herwig, R.P.; Lara, J.C.; Staley, J.T. Cycloclasticus pugetii gen. nov., sp. nov., an aromatic hydrocarbon-degrading bacterium from marine sediments. Int. J. Syst. Evol. Microbiol. 1995, 45, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Yakimov, M.M.; Timmis, K.N.; Golyshin, P.N. Obligate oil-degrading marine bacteria. Curr. Opin. Biotechnol. 2007, 18, 257–266. [Google Scholar] [CrossRef]
- Atlas, R.M. Petroleum biodegradation and oil spill bioremediation. Mar. Pollut. Bull. 1995, 31, 178–182. [Google Scholar] [CrossRef]
- Lee, W.; Morris, A.; Boatwright, D. Mexican oil spill: A toxicity study of oil accommodated in seawater on marine invertebrates. Mar. Pollut. Bull. 1980, 11, 231–234. [Google Scholar] [CrossRef]
- Teramoto, M.; Suzuki, M.; Okazaki, F.; Hatmanti, A.; Harayama, S. Oceanobacter-related bacteria are important for the degradation of petroleum aliphatic hydrocarbons in the tropical marine environment. Microbiology 2009, 155, 3362–3370. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, A.; Ishiwata, H.; Kitamura, K.; Sunamura, M.; Fujita, T.; Matsuo, M.; Higashihara, T. Dynamics of microbial populations and strong selection for Cycloclasticus pugetii following the Nakhodka oil spill. Microb. Ecol. 2003, 46, 442–453. [Google Scholar] [CrossRef]
- Jimeson, R.; Radosevich, M.; Stevens, R.; Giardino-Radosevich, G. Mixed Alcohol Fuels for Internal Combustion Engines, Furnaces, Boilers, Kilns and Gasifiers. U.S. Patent 8,277,522 B2, 2 October 2012. [Google Scholar]
- Chou, C.-C.; Riviere, J.E.; Monteiro-Riviere, N.A. Differential relationship between the carbon chain length of jet fuel aliphatic hydrocarbons and their ability to induce cytotoxicity vs. interleukin-8 release in human epidermal keratinocytes. Toxicol. Sci. 2002, 69, 226–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Istoto, E.H.; Saptadi, S. Production of Fuels From HDPE and LDPE Plastic Waste via Pyrolysis Methods. In E3S Web of Conferences; EDP Sciences: Ulis, France, 2019; p. 14011. [Google Scholar]
- Bayat, H.; Cheng, F.; Dehghanizadeh, M.; Soliz, N.; Brewer, C.E.; Jena, U. Hydrothermal Liquefaction of Food Waste: Bio-crude oil Characterization, Mass and Energy Balance. In 2019 ASABE Annual International Meeting; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2019. [Google Scholar]
- Cerniglia, C. Microbial degradation of polycyclic aromatic hydrocarbons (PAH) in the aquatic environment. Metab. Polycycl. Aromat. Hydrocarb. Aquat. Environ. 1989, 38, 41–68. [Google Scholar]
- Solano-Serena, F.; Marchal, R.; Casarégola, S.; Vasnier, C.; Lebeault, J.-M.; Vandecasteele, J.-P. A Mycobacterium strain with extended capacities for degradation of gasoline hydrocarbons. Appl. Environ. Microbiol. 2000, 66, 2392–2399. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, L.B.; van Zelm, R.; Elshout, P.M.; Hendriks, A.J.; Leuven, R.S.; Struijs, J.; de Zwart, D.; Huijbregts, M.A. Species richness–phosphorus relationships for lakes and streams worldwide. Glob. Ecol. Biogeogr. 2013, 22, 1304–1314. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Gu, X.; Zhang, R.; Chen, L.; Lu, J. Microbial degradation of diesel oil and heavy oil in the presence of modified clay. Energy Sources Part A Recovery Util. Environ. Eff. 2017, 39, 326–331. [Google Scholar] [CrossRef]
- Adebusoye, S.A.; Ilori, M.O.; Amund, O.O.; Teniola, O.D.; Olatope, S. Microbial degradation of petroleum hydrocarbons in a polluted tropical stream. World J. Microbiol. Biotechnol. 2007, 23, 1149–1159. [Google Scholar] [CrossRef]
- Mandri, T.; Lin, J. Isolation and characterization of engine oil degrading indigenous microrganisms in Kwazulu-Natal, South Africa. Afr. J. Biotechnol. 2007, 6, 23–27. [Google Scholar]
- Darvishi, P.; Mowla, D.; Ayatollahi, S.; Niazi, A. Biodegradation of heavy crude oil in wastewater by an efficient strain, ERCPPI-1. Desalination Water Treat. 2011, 28, 46–54. [Google Scholar] [CrossRef]
- Rajasekar, A.; Maruthamuthu, S.; Ting, Y.-P.; Balasubramanian, R.; Rahman, P.K. Bacterial degradation of petroleum hydrocarbons. In Microbial Degradation of Xenobiotics; Springer: Berlin/Heidelberg, Germany, 2012; pp. 339–369. [Google Scholar]
- Bacosa, H.P.; Mabuhay-Omar, J.A.; Balisco, R.A.T.; Omar, D.M.; Inoue, C. Biodegradation of binary mixtures of octane with benzene, toluene, ethylbenzene or xylene (BTEX): Insights on the potential of Burkholderia, Pseudomonas and Cupriavidus isolates. World J. Microbiol. Biotechnol. 2021, 37, 122. [Google Scholar] [CrossRef]
- Rahman, K.; Thahira-Rahman, J.; Lakshmanaperumalsamy, P.; Banat, I.M. Towards efficient crude oil degradation by a mixed bacterial consortium. Bioresour. Technol. 2002, 85, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Tzarkova, E.; Groudeva, V. Bioremediation of Petroleum—Polluted Soils by Adsorptive Immobilized Corynebacterium Sp. RB—96. Biotechnol. Biotechnol. Equip. 2000, 14, 72–75. [Google Scholar] [CrossRef]
- Sathishkumar, M.; Binupriya, A.R.; Baik, S.H.; Yun, S.E. Biodegradation of crude oil by individual bacterial strains and a mixed bacterial consortium isolated from hydrocarbon contaminated areas. CLEAN—Soil Air Water 2008, 36, 92–96. [Google Scholar] [CrossRef]
- Nguyen, U.T.; Lincoln, S.A.; Valladares Juárez, A.G.; Schedler, M.; Macalady, J.L.; Müller, R.; Freeman, K.H. The influence of pressure on crude oil biodegradation in shallow and deep Gulf of Mexico sediments. PLoS ONE 2018, 13, e0199784. [Google Scholar] [CrossRef] [PubMed]
- Schedler, M.; Hiessl, R.; Juárez, A.G.V.; Gust, G.; Müller, R. Effect of high pressure on hydrocarbon-degrading bacteria. AMB Express 2014, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Scoma, A.; Boon, N. Osmotic stress confers enhanced cell integrity to hydrostatic pressure but impairs growth in Alcanivorax borkumensis SK2. Front. Microbiol. 2016, 7, 729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scoma, A.; Yakimov, M.M.; Boon, N. Challenging oil bioremediation at deep-sea hydrostatic pressure. Front. Microbiol. 2016, 7, 1203. [Google Scholar] [CrossRef] [Green Version]
- Hackbusch, S.; Noirungsee, N.; Viamonte, J.; Sun, X.; Bubenheim, P.; Kostka, J.E.; Müller, R.; Liese, A. Influence of pressure and dispersant on oil biodegradation by a newly isolated Rhodococcus strain from deep-sea sediments of the Gulf of Mexico. Mar. Pollut. Bull. 2020, 150, 110683. [Google Scholar] [CrossRef]
- Dutta, A.; Peoples, L.M.; Gupta, A.; Bartlett, D.H.; Sar, P. Exploring the piezotolerant/piezophilic microbial community and genomic basis of piezotolerance within the deep subsurface Deccan traps. Extremophiles 2019, 23, 421–433. [Google Scholar] [CrossRef]
- Yayanos, A.A. Microbiology to 10,500 meters in the deep sea. Annu. Rev. Microbiol. 1995, 49, 777–805. [Google Scholar] [CrossRef]
- Oger, P.M.; Jebbar, M. The many ways of coping with pressure. Res. Microbiol. 2010, 161, 799–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, O.U.; Han, J.; Woyke, T.; Jansson, J.K. Single-cell genomics reveals features of a Colwellia species that was dominant during the Deepwater Horizon oil spill. Front. Microbiol. 2014, 5, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marietou, A.; Chastain, R.; Beulig, F.; Scoma, A.; Hazen, T.C.; Bartlett, D.H. The effect of hydrostatic pressure on enrichments of hydrocarbon degrading microbes from the Gulf of Mexico following the Deepwater Horizon oil spill. Front. Microbiol. 2018, 9, 808. [Google Scholar] [CrossRef] [PubMed]
- Grossi, V.; Yakimov, M.M.; Al Ali, B.; Tapilatu, Y.; Cuny, P.; Goutx, M.; La Cono, V.; Giuliano, L.; Tamburini, C. Hydrostatic pressure affects membrane and storage lipid compositions of the piezotolerant hydrocarbon-degrading Marinobacter hydrocarbonoclasticus strain# 5. Environ. Microbiol. 2010, 12, 2020–2033. [Google Scholar] [PubMed]
- Perez Calderon, L.J.; Gontikaki, E.; Potts, L.D.; Shaw, S.; Gallego, A.; Anderson, J.A.; Witte, U. Pressure and temperature effects on deep-sea hydrocarbon-degrading microbial communities in subarctic sediments. Microbiologyopen 2019, 8, e00768. [Google Scholar] [CrossRef]
- ZoBell, C.E.; Johnson, F.H. The influence of hydrostatic pressure on the growth and viability of terrestrial and marine bacteria. J. Bacteriol. 1949, 57, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Prince, R.C.; Lessard, R.R. Crude oil releases to the environment: Natural fate and remediation options. Encycl. Energy 2004, 1, 727–736. [Google Scholar]
- Scoma, A.; Barbato, M.; Hernandez-Sanabria, E.; Mapelli, F.; Daffonchio, D.; Borin, S.; Boon, N. Microbial oil-degradation under mild hydrostatic pressure (10 MPa): Which pathways are impacted in piezosensitive hydrocarbonoclastic bacteria? Sci. Rep. 2016, 6, 23526. [Google Scholar] [CrossRef] [Green Version]
- Kimes, N.E.; Callaghan, A.V.; Aktas, D.F.; Smith, W.L.; Sunner, J.; Golding, B.T.; Drozdowska, M.; Hazen, T.C.; Suflita, J.M.; Morris, P.J. Metagenomic analysis and metabolite profiling of deep–sea sediments from the Gulf of Mexico following the Deepwater Horizon oil spill. Front. Microbiol. 2013, 4, 50. [Google Scholar] [CrossRef] [Green Version]
- Prince, R.C.; Nash, G.W.; Hill, S.J. The biodegradation of crude oil in the deep ocean. Mar. Pollut. Bull. 2016, 111, 354–357. [Google Scholar] [CrossRef]
- Aulenta, F.; Palma, E.; Marzocchi, U.; Cruz Viggi, C.; Rossetti, S.; Scoma, A. Enhanced Hydrocarbons Biodegradation at Deep-Sea Hydrostatic Pressure with Microbial Electrochemical Snorkels. Catalysts 2021, 11, 263. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F. Biodegradation and bioremediation of hydrocarbons in extreme environments. Appl. Microbiol. Biotechnol. 2001, 56, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Louvado, A.; Gomes, N.; Simões, M.M.; Almeida, A.; Cleary, D.F.; Cunha, A. Polycyclic aromatic hydrocarbons in deep sea sediments: Microbe–pollutant interactions in a remote environment. Sci. Total Environ. 2015, 526, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.; Liu, J.; Bacosa, H.; Rosenheim, B.E.; Liu, Z. Petroleum hydrocarbon persistence following the Deepwater Horizon oil spill as a function of shoreline energy. Mar. Pollut. Bull. 2017, 115, 47–56. [Google Scholar] [CrossRef]
- Michel, J.; Hayes, M.O. Weathering patterns of oil residues eight years after the Exxon Valdez oil spill. Mar. Pollut. Bull. 1999, 38, 855–863. [Google Scholar] [CrossRef]
- Sauer, T.C.; Michel, J.; Hayes, M.O.; Aurand, D.V. Hydrocarbon characterization and weathering of oiled intertidal sediments along the Saudi Arabian Coast two years after the Gulf War oil spill. Environ. Int. 1998, 24, 43–60. [Google Scholar] [CrossRef]
- Peterson, C.H.; Rice, S.D.; Short, J.W.; Esler, D.; Bodkin, J.L.; Ballachey, B.E.; Irons, D.B. Long-term ecosystem response to the Exxon Valdez oil spill. Science 2003, 302, 2082–2086. [Google Scholar] [CrossRef] [Green Version]
- Owens, E.H.; Taylor, E.; Humphrey, B. The persistence and character of stranded oil on coarse-sediment beaches. Mar. Pollut. Bull. 2008, 56, 14–26. [Google Scholar] [CrossRef]
- Hayes, M.O.; Gundlach, E.R.; D‘Ozouville, L. Role of dynamic coastal processes in the impact and dispersal of the Amoco Cadiz oil spill (March 1978) Brittany, France. In International Oil Spill Conference; American Petroleum Institute: Washington, DC, USA, 1979; pp. 193–198. [Google Scholar]
- Hayes, M.O. An exposure index for oiled shorelines. Spill Sci. Technol. Bull. 1996, 3, 139–147. [Google Scholar] [CrossRef]
- Pendergraft, M.A.; Rosenheim, B.E. Varying relative degradation rates of oil in different forms and environments revealed by ramped pyrolysis. Environ. Sci. Technol. 2014, 48, 10966–10974. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M. Degradation of Bunker C oil under different coastal environments of Chedabucto Bay, Nova Scotia. Estuar. Coast. Mar. Sci. 1974, 2, 137–144. [Google Scholar] [CrossRef]
- Cui, F.; Daskiran, C.; King, T.; Robinson, B.; Lee, K.; Katz, J.; Boufadel, M.C. Modeling oil dispersion under breaking waves. Part I: Wave hydrodynamics. Environ. Fluid Mech. 2020, 20, 1527–1551. [Google Scholar] [CrossRef]
- Tkalich, P.; Chan, E.S. Vertical mixing of oil droplets by breaking waves. Mar. Pollut. Bull. 2002, 44, 1219–1229. [Google Scholar] [CrossRef]
- Adzigbli, L.; Bacosa, H.P.; Deng, Y. Response of microbial communities to oil spill in the Gulf of Mexico: A review. Afr. J. Microbiol. Res. 2018, 12, 536–545. [Google Scholar]
- Tremblay, J.; Yergeau, E.; Fortin, N.; Cobanli, S.; Elias, M.; King, T.L.; Lee, K.; Greer, C.W. Chemical dispersants enhance the activity of oil-and gas condensate-degrading marine bacteria. ISME J. 2017, 11, 2793–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaerun, S.K.; Tazaki, K.; Asada, R.; Kogure, K. Interaction between clay minerals and hydrocarbon-utilizing indigenous microorganisms in high concentrations of heavy oil: Implications for bioremediation. Clay Miner. 2005, 40, 105–114. [Google Scholar] [CrossRef]
- Varjani, S.J.; Upasani, V.N. A new look on factors affecting microbial degradation of petroleum hydrocarbon pollutants. Int. Biodeterior. Biodegrad. 2017, 120, 71–83. [Google Scholar] [CrossRef]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum hydrocarbon-degrading bacteria for the remediation of oil pollution under aerobic conditions: A perspective analysis. Front. Microbiol. 2018, 9, 2885. [Google Scholar] [CrossRef]
- Deppe, U.; Richnow, H.-H.; Michaelis, W.; Antranikian, G. Degradation of crude oil by an arctic microbial consortium. Extremophiles 2005, 9, 461–470. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Inoue, C. Heavy oil degrading Burkholderia and Pseudomonas strains: Insights on the degradation potential of isolates and microbial consortia. Palawan Sci. 2020, 12, 74–89. [Google Scholar]
- Tang, X.; He, L.; Tao, X.; Dang, Z.; Guo, C.; Lu, G.; Yi, X. Construction of an artificial microalgal-bacterial consortium that efficiently degrades crude oil. J. Hazard. Mater. 2010, 181, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Pelz, O.; Tesar, M.; Wittich, R.M.; Moore, E.R.; Timmis, K.N.; Abraham, W.R. Towards elucidation of microbial community metabolic pathways: Unravelling the network of carbon sharing in a pollutant-degrading bacterial consortium by immunocapture and isotopic ratio mass spectrometry. Environ. Microbiol. 1999, 1, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, F.A.; Boufadel, M.C.; Johnson, R.; Lee, K.; Graan, T.P.; Bejarano, A.C.; Zhu, Z.; Waterman, D.; Capone, D.M.; Hayter, E. Oil-Particle Interactions and Submergence from Crude Oil Spills in Marine and Freshwater Environments: Review of the Science and Future Science Needs; U.S. Geological Survey: Reston, VA, USA, 2015.
- Adhikari, P.; Maiti, K.; Bam, W. Fate of particle-bound polycyclic aromatic hydrocarbons in the river-influenced continental margin of the northern Gulf of Mexico. Mar. Pollut. Bull. 2019, 141, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, P.L.; Maiti, K.; Overton, E.B. Vertical fluxes of polycyclic aromatic hydrocarbons in the northern Gulf of Mexico. Mar. Chem. 2015, 168, 60–68. [Google Scholar] [CrossRef]
- Adhikari, P.L.; Maiti, K.; Bosu, S.; Jones, P.R. 234Th as a tracer of vertical transport of polycyclic aromatic hydrocarbons in the northern Gulf of Mexico. Mar. Pollut. Bull. 2016, 107, 179–187. [Google Scholar] [CrossRef]
- Bragg, J.R.; Owens, E.H. Shoreline cleansing by interactions between oil and fine mineral particles. In International Oil Spill Conference; American Petroleum Institute: Columbia, WA, USA, 1995; pp. 219–227. [Google Scholar]
- Lee, K.; Weise, A.M.; St-Pierre, S. Enhanced oil biodegradation with mineral fine interaction. Spill Sci. Technol. Bull. 1996, 3, 263–267. [Google Scholar] [CrossRef]
- Owens, E.H. The interaction of fine particles with stranded oil. Pure Appl. Chem. 1999, 71, 83–93. [Google Scholar] [CrossRef]
- Daly, K.L.; Passow, U.; Chanton, J.; Hollander, D. Assessing the impacts of oil-associated marine snow formation and sedimentation during and after the Deepwater Horizon oil spill. Anthropocene 2016, 13, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Warr, L.N.; Friese, A.; Schwarz, F.; Schauer, F.; Portier, R.J.; Basirico, L.M.; Olson, G.M. Bioremediating oil spills in nutrient poor ocean waters using fertilized clay mineral flakes: Some experimental constraints. Biotechnol. Res. Int. 2013, 2013, 704806. [Google Scholar] [CrossRef]
- Loh, A.; Yim, U.H. A review of the effects of particle types on oil-suspended particulate matter aggregate formation. Ocean Sci. J. 2016, 51, 535–548. [Google Scholar] [CrossRef]
- Wang, W.; Zheng, Y.; Li, Z.; Lee, K. PIV investigation of oil–mineral interaction for an oil spill application. Chem. Eng. J. 2011, 170, 241–249. [Google Scholar] [CrossRef]
- Giani, M.; Rinaldi, A.; Degobbis, D. Mucilages in the Adriatic and Tyrrhenian Sea: An introduction. Sci. Total Environ. 2005, 353, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Alldredge, A.L.; Silver, M.W. Characteristics, dynamics and significance of marine snow. Prog. Oceanogr. 1988, 20, 41–82. [Google Scholar] [CrossRef]
- Patton, J.S.; Rigler, M.W.; Boehm, P.D.; Fiest, D.L. Ixtoc 1 oil spill: Flaking of surface mousse in the Gulf of Mexico. Nature 1981, 290, 235–238. [Google Scholar] [CrossRef]
- Dell’Amore, C. “Sea Snot” Explosion Caused by Gulf Oil Spill? 2010. Available online: https://www.nationalgeographic.com/science/article/100916-sea-snot-gulf-bp-oil-spill-marine-snow-science-environment (accessed on 30 January 2022).
- Ramachandran, S.D.; Sweezey, M.J.; Hodson, P.V.; Boudreau, M.; Courtenay, S.C.; Lee, K.; King, T.; Dixon, J.A. Influence of salinity and fish species on PAH uptake from dispersed crude oil. Mar. Pollut. Bull. 2006, 52, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Thavasi, R.; Jayalakshmi, S.; Balasubramanian, T.; Banat, I.M. Effect of salinity, temperature, pH and crude oil concentration on biodegradation of crude oil by Pseudomonas aeruginosa. J. Biol. Environ. Sci. 2007, 1, 51–57. [Google Scholar]
- Si-Zhong, Y.; Hui-Jun, J.; Zhi, W.; Rui-Xia, H.; Yan-Jun, J.; Xiu-Mei, L.; Shao-Peng, Y. Bioremediation of oil spills in cold environments: A review. Pedosphere 2009, 19, 371–381. [Google Scholar]
- Cao, Y.; Zhang, B.; Zhu, Z.; Song, X.; Cai, Q.; Chen, B.; Dong, G.; Ye, X. Microbial eco-physiological strategies for salinity-mediated crude oil biodegradation. Sci. Total Environ. 2020, 727, 138723. [Google Scholar] [CrossRef]
- Jamal, M.T.; Pugazhendi, A. Degradation of petroleum hydrocarbons and treatment of refinery wastewater under saline condition by a halophilic bacterial consortium enriched from marine environment (Red Sea), Jeddah, Saudi Arabia. 3 Biotech 2018, 8, 276. [Google Scholar] [CrossRef]
- Duran, R.; Cravo-Laureau, C. Role of environmental factors and microorganisms in determining the fate of polycyclic aromatic hydrocarbons in the marine environment. FEMS Microbiol. Rev. 2016, 40, 814–830. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Bai, X.; Sheng, H.; Jiao, L.; Zhou, H.; Shao, Z. Distribution of PAHs and the PAH-degrading bacteria in the deep-sea sediments of the high-latitude Arctic Ocean. Biogeosciences 2015, 12, 2163–2177. [Google Scholar] [CrossRef] [Green Version]
- Al-Awadhi, H.; Dashti, N.; Kansour, M.; Sorkhoh, N.; Radwan, S. Hydrocarbon-utilizing bacteria associated with biofouling materials from offshore waters of the Arabian Gulf. Int. Biodeterior. Biodegrad. 2012, 69, 10–16. [Google Scholar] [CrossRef]
- Somee, M.R.; Shavandi, M.; Dastgheib, S.M.M.; Amoozegar, M.A. Bioremediation of oil-based drill cuttings by a halophilic consortium isolated from oil-contaminated saline soil. 3 Biotech 2018, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Mnif, S.; Chamkha, M.; Sayadi, S. Isolation and characterization of Halomonas sp. strain C2SS100, a hydrocarbon-degrading bacterium under hypersaline conditions. J. Appl. Microbiol. 2009, 107, 785–794. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, H.; Mao, X.; Li, R. Biodegradation of phenanthrene by a halophilic bacterial consortium under aerobic conditions. Curr. Microbiol. 2009, 58, 205–210. [Google Scholar] [CrossRef]
- Dastgheib, S.M.M.; Amoozegar, M.A.; Khajeh, K.; Shavandi, M.; Ventosa, A. Biodegradation of polycyclic aromatic hydrocarbons by a halophilic microbial consortium. Appl. Microbiol. Biotechnol. 2012, 95, 789–798. [Google Scholar] [CrossRef]
- Al-Mailem, D.; Sorkhoh, N.; Al-Awadhi, H.; Eliyas, M.; Radwan, S. Biodegradation of crude oil and pure hydrocarbons by extreme halophilic archaea from hypersaline coasts of the Arabian Gulf. Extremophiles 2010, 14, 321–328. [Google Scholar] [CrossRef]
- Khemili-Talbi, S.; Kebbouche-Gana, S.; Akmoussi-Toumi, S.; Angar, Y.; Gana, M.L. Isolation of an extremely halophilic arhaeon Natrialba sp. C21 able to degrade aromatic compounds and to produce stable biosurfactant at high salinity. Extremophiles 2015, 19, 1109–1120. [Google Scholar]
- Okpokwasili, G.; Odokuma, L. Effect of salinity on biodegradation of oil spill dispersants. Waste Manag. 1990, 10, 141–146. [Google Scholar] [CrossRef]
- Sun, J.; Zheng, X. A review of oil-suspended particulate matter aggregation—a natural process of cleansing spilled oil in the aquatic environment. J. Environ. Monit. 2009, 11, 1801–1809. [Google Scholar] [CrossRef]
- Wells, P.G.; Butler, J.N.; Hughes, J.S. Exxon Valdez Oil Spill: Fate and Effects in Alaskan Waters; ASTM: West Conshohocken, PA, USA, 1995. [Google Scholar]
- Díaz, M.P.; Boyd, K.G.; Grigson, S.J.; Burgess, J.G. Biodegradation of crude oil across a wide range of salinities by an extremely halotolerant bacterial consortium MPD-M, immobilized onto polypropylene fibers. Biotechnol. Bioeng. 2002, 79, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Megharaj, M.; Venkateswarlu, K.; Naidu, R. Bioremediation. In Encyclopedia of Toxicology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 1, pp. 485–489. [Google Scholar]
- Megharaj, M.; Naidu, R. Soil and brownfield bioremediation. Microb. Biotechnol. 2017, 10, 1244–1249. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, S.; Thavamani, P.; Megharaj, M.; Lee, Y.B.; Naidu, R. Isolation and characterization of polycyclic aromatic hydrocarbons (PAHs) degrading, pH tolerant, N-fixing and P-solubilizing novel bacteria from manufactured gas plant (MGP) site soils. Environ. Technol. Innov. 2016, 6, 204–219. [Google Scholar] [CrossRef]
- Piontek, J.; Lunau, M.; Händel, N.; Borchard, C.; Wurst, M.; Engel, A. Acidification increases microbial polysaccharide degradation in the ocean. Biogeosciences 2010, 7, 1615–1624. [Google Scholar] [CrossRef] [Green Version]
- Radwan, S.; Al-Hasan, R.; Ali, N.; Salamah, S.; Khanafer, M. Oil-consuming microbial consortia floating in the Arabian Gulf. Int. Biodeterior. Biodegrad. 2005, 56, 28–33. [Google Scholar] [CrossRef]
- Hui, L.; Zhang, Y.; Kravchenko, I.; Hui, X.; Zhang, C.-g. Dynamic changes in microbial activity and community structure during biodegradation of petroleum compounds: A laboratory experiment. J. Environ. Sci. 2007, 19, 1003–1013. [Google Scholar]
- Okoh, A.I. Biodegradation alternative in the cleanup of petroleum hydrocarbon pollutants. Biotechnol. Mol. Biol. Rev. 2006, 1, 38–50. [Google Scholar]
- Fathepure, B.Z. Recent studies in microbial degradation of petroleum hydrocarbons in hypersaline environments. Front. Microbiol. 2014, 5, 173. [Google Scholar] [CrossRef] [Green Version]
- Pawar, R.M. The effect of soil pH on bioremediation of polycyclic aromatic hydrocarbons (PAHS). J. Bioremediat. Biodegrad. 2015, 6, 291–304. [Google Scholar] [CrossRef]
- Obahiagbon, K.O.; Amenaghawon, A.N.; Agbonghae, E.O. Effect of initial pH on the bioremediation of crude oil polluted water using a consortium of microbes. Pasific J. Sci. Technol. 2014, 15, 452–457. [Google Scholar]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reactor | Redox Potential (mV) | Biodegradation (%) | ||
|---|---|---|---|---|
| Extractable Organic Matter (EOM) | Saturated Fraction (FS) | Aromatic Fraction (FA) | ||
| 3 (8 ppm O2, stirring) | 150 | 64 | 70 | 59 |
| 4 (8 ppm O2, without stirring) | 90 | 55 | 61 | 52 |
| 5 (2–3 ppm O2) | 30 | 11 | 20 | 17 |
| 6 (0.2–0.3 ppm O2) | −190 | NS | NS | NS |
| Oil | Carbon Composition | Oil Concentration | % of Oil Biodegraded | Dominant Bacteria | Isolate or Consortium | References |
|---|---|---|---|---|---|---|
| Light oil | C4–C13 | 7% | 75% | Pseudomonas sp. | Isolate | [118] |
| Gasoline | C1–C10 | 10% | 86% | Mycobacterium austroafricanum | Isolate | [116] |
| Kerosene | C10–C18 | 1% | 20% | Achromobacter, Alcaligenes, Cupriavidus, Burkholderia, Rhodanobacter | Consortium | [16] |
| Diesel | >C12 | 1% | 85% | Pseudomonas aeruginosa, Acinetobacter sp., Corynebacterium sp. | Consortium | [119] |
| Engine oil/Lubricating oil | C9–C16 | 10% | 80–90% | Flavobacterium sp., Acinetobacterium calcoaceticum, and Pseudomonas aeruginosa | Consortium | [120] |
| Heavy oil | C20–C50 | 1% | 21–30% | Pseudomonas spp. Burkholderia spp. | Consortium | [17] |
| Enterobacteriaceae | [121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bacosa, H.P.; Ancla, S.M.B.; Arcadio, C.G.L.A.; Dalogdog, J.R.A.; Ellos, D.M.C.; Hayag, H.D.A.; Jarabe, J.G.P.; Karim, A.J.T.; Navarro, C.K.P.; Palma, M.P.I.; et al. From Surface Water to the Deep Sea: A Review on Factors Affecting the Biodegradation of Spilled Oil in Marine Environment. J. Mar. Sci. Eng. 2022, 10, 426. https://doi.org/10.3390/jmse10030426
Bacosa HP, Ancla SMB, Arcadio CGLA, Dalogdog JRA, Ellos DMC, Hayag HDA, Jarabe JGP, Karim AJT, Navarro CKP, Palma MPI, et al. From Surface Water to the Deep Sea: A Review on Factors Affecting the Biodegradation of Spilled Oil in Marine Environment. Journal of Marine Science and Engineering. 2022; 10(3):426. https://doi.org/10.3390/jmse10030426
Chicago/Turabian StyleBacosa, Hernando Pactao, Sheila Mae B. Ancla, Cris Gel Loui A. Arcadio, John Russel A. Dalogdog, Dioniela Mae C. Ellos, Heather Dale A. Hayag, Jiza Gay P. Jarabe, Ahl Jimhar T. Karim, Carl Kenneth P. Navarro, Mae Princess I. Palma, and et al. 2022. "From Surface Water to the Deep Sea: A Review on Factors Affecting the Biodegradation of Spilled Oil in Marine Environment" Journal of Marine Science and Engineering 10, no. 3: 426. https://doi.org/10.3390/jmse10030426