
Effects of Miniaturization of the Summer Phytoplankton Community on the Marine Ecosystem in the Northern East China Sea

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cruises and Sampling
2.2. Dissolved Inorganic Nutrients
2.3. Phytoplankton Abundances and Dominant Species
2.4. Picophytoplankton Abundances
2.5. Chl-a Size Fractionation
2.6. Data Analyses
3. Results
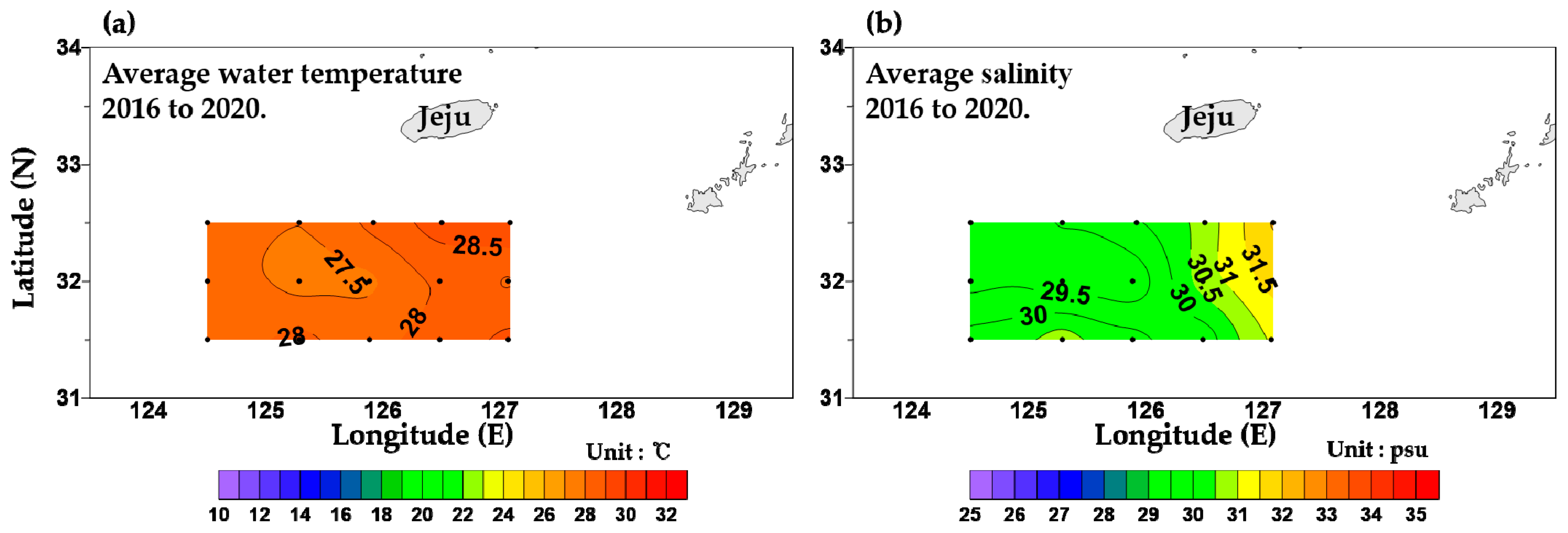
3.1. Physical Environments
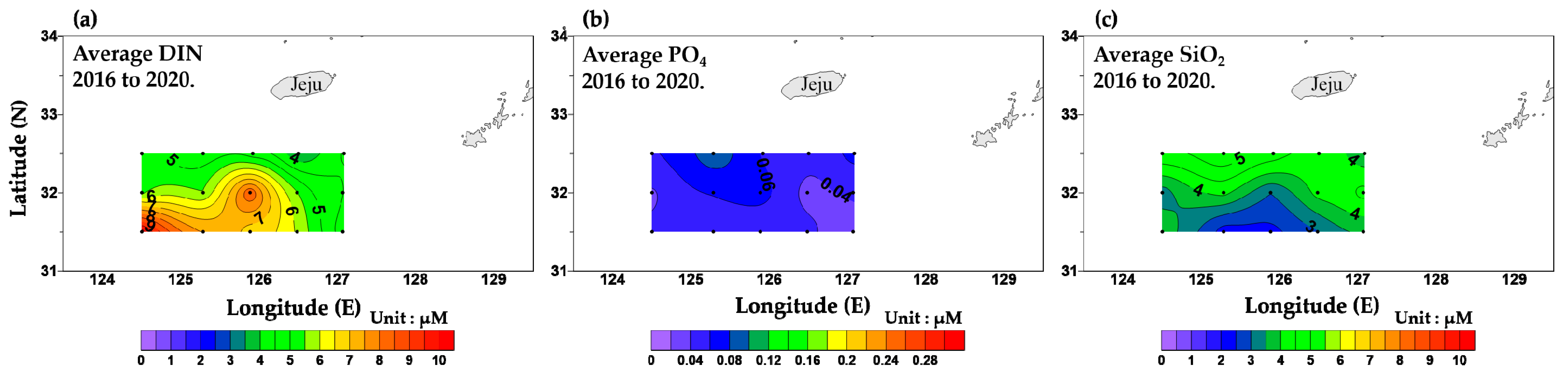
3.2. Dissolved Inorganic Nutrient Concentrations
3.3. Phytoplankton Abundance and Dominant Species
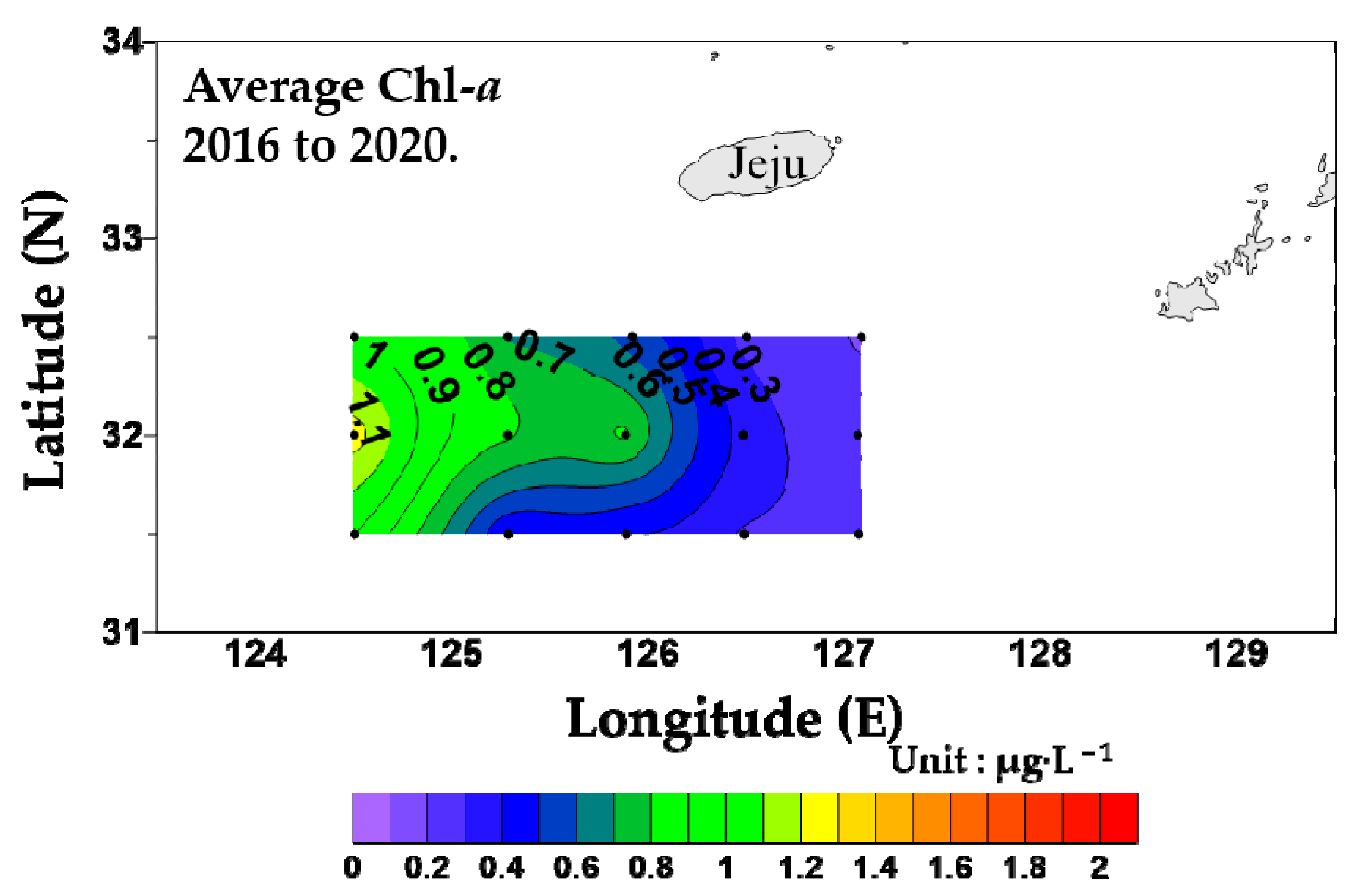
3.4. Relative Contributions of Size-Fractionated Chl-a to the Overall Chl-a Concentration
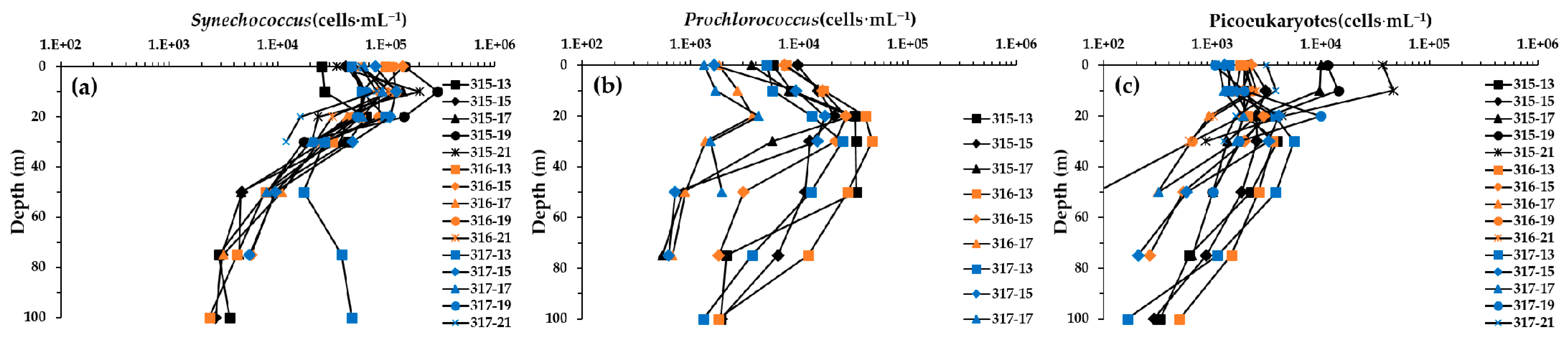
3.5. Picophytoplankton Cell Abundances
4. Discussion
4.1. Changes in Phytoplankton-Community Structures in Response to Changes in the Summer Marine Environment
4.2. Changes in Phytoplankton Community Sizes and Structures in the Northern East China Sea
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Q.L.; Weng, X.C. Analysis of water masses in the south Yellow Sea in spring. Oceanol. Limnol. Sin. 1996, 27, 421–428. [Google Scholar]
- Rho, H.K. Studies on Marine Environment of Fishing Grounds in the Waters around Cheju Island. Ph.D. Thesis, Tokyo University, Tokyo, Japan, 1985. [Google Scholar]
- Miyaji, K. Studies on the eddies associated with the meander of the Kuroshio in the waters off southwest coast of Kyushu and their effects on egg and larval transport. Bull. Seikai Nat. Fish. Res. Inst. 1991, 69, 1–77. [Google Scholar]
- Guo, Y.J.; Zhang, Y.S. Characteristics of phytoplankton distribution in Yellow Sea. J. Yellow Sea 1996, 2, 90–103. [Google Scholar]
- Simpson, J.H.; Pingree, R.D. Shallow sea fronts produced by tidal stirring. In Oceanic Fronts in Coastal Processes; Bowman, M.J., Esaias, W.E., Eds.; Springer: Berlin/Heidelberg, Germany, 1978; pp. 29–47. [Google Scholar]
- Beardsley, R.; Limebumer, R.; Yu, H.; Cannon, G.A. Discharge of the Changjiang (Yangtze River) into the East China Sea. Cont. Shelf Res. 1985, 4, 57–76. [Google Scholar] [CrossRef]
- Su, Y.S.; Weng, X.C. Water masses in China Seas. In Oceanography of China Seas; Zhou, D., Liang, Y.B., Zeng, C.K., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 3–16. [Google Scholar]
- Edmond, J.M.; Spivack, A.; Grant, B.C.; Chen, Z.; Chen, S.; Zeng, X. Chemical dynamics of the Changjiang Estuary. Cont. Shelf Res. 1985, 4, 17–36. [Google Scholar] [CrossRef]
- Hama, T.; Shin, K.H.; Handa, N. Spatial variability in the primary productivity in the East China Sea and its adjacent water. J. Oceanogr. 1997, 53, 41–51. [Google Scholar] [CrossRef]
- Gong, G.C.; Wen, Y.H.; Wang, B.W.; Liu, G.J. Seasonal variation of chlorophyll a concentration, primary production and environmental conditions in the subtropical East China Sea. Deep Sea Res. Part II 2003, 50, 1219–1236. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, H.Y.; Gong, G.C.; Lin, Y.H.; Jan, S.; Takahashi, M. Phytoplankton production during a summer coastal upwelling in the East China Sea. Cont. Shelf Res. 2004, 24, 1321–1338. [Google Scholar] [CrossRef]
- Koroji, K.; Matsusaki, M. Present activity on marine biology in Japan Meteorological Agency. Sokuko Jiho 1980, 47, 19–21. [Google Scholar]
- Chin, T.G.; Cheng, Z.M.; Lin, J.; Lin, S. Diatoms from the surface sediments of the east China Sea. Acta Oceanol. Sin. 1980, 2, 97–110. [Google Scholar]
- Huang, R. The influence of hydrography on the distribution of phytoplankton in the southern Taiwan strait. Estuar. Coast. Shelf Sci. 1988, 26, 643–656. [Google Scholar] [CrossRef]
- Koizumi, I. Pliocene and Pleistocene diatom datum levels related with Paleoceanography in the northwest Pacific. Mar. Micropaleontol. 1986, 10, 309–325. [Google Scholar] [CrossRef]
- Yu, J.L.; Li, R.X. Species composition and quantitative distribution of planktonic diatom in the Kuroshio current and its adjacent waters. In Selected Papers on the Investigation of Kuroshio; Su, J.L., Ed.; Ocean Press: Beijing, China, 1990; Volume 2, pp. 57–66. [Google Scholar]
- Guo, Y.J. The Kuroshio, part 2. Primary productivity and phytoplankton. Oceanogr. Mar. 1991, 29, 155–189. [Google Scholar]
- Huang, R. Phytoplankton distribution in the South China Sea and Kuroshio-flowing region of Taiwan. Acta Oceanogr. Taiwan. 1993, 31, 73–82. [Google Scholar]
- Matsuda, O.; Nishi, Y.; Yoon, Y.H.; Endo, T. Observation of thermohaline structure and phytoplankton biomass the shelf front of East China Sea during early summer. J. Fac. Appl. Biol. Sci. 1989, 28, 27–35. [Google Scholar]
- Kamiya, H. The correlation between appearance of phytoplankton and the sea condition. Umi Sora 1991, 67, 153–162. [Google Scholar]
- Ueno, S. Phytoplankton communities and their relation to the water types in the vicinity of the Kuroshio front in the East China. Sea J. Shimonoseki Univ. Fish. 1993, 41, 251–256. [Google Scholar]
- Meng, F.; Huang, F.; Zhaodang, M.; Qinliang, L. On the composition and distribution of zooplankton species in the Kuroshio region in the north of the East China Sea. In Selected Papers on the Investigation of Kuroshio; Su, J.L., Ed.; Ocean Press: Beijing, China, 1991; Volume 3, pp. 150–161. [Google Scholar]
- Zhang, H.Q. Relationship between zooplankton distribution and hydrographic characteristics of the southern Yellow Sea. Yellow Sea 1995, 1, 50–67. [Google Scholar]
- Choi, C.H.; Lee, C.R.; Kang, H.K.; Kang, K.A. Characteristics and variation of size-fractionated zooplankton biomass in the northern East China Sea. Ocean Polar Res. 2011, 33, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Oh, G.S. Characteristics and Variation of Zooplankton Community Structure in the Northern East China Sea. Master’s Thesis, Chonnam National University, Yeosu, Korea, 2012. [Google Scholar]
- Dong, H.; Ren, D.; Li, B. The distribution and influential factors of oxygen and phosphate in Kuroshio area of the East China Sea north to 30 degree N in summer and winter 1987. In Selected Papers on the Investigation of Kuroshio; Su, J.L., Ed.; Ocean Press: Beijing, China, 1990; Volume 2, pp. 208–217. [Google Scholar]
- Noh, J.H. A Study on the Phytoplankton Distribution in the Yellow Sea and the East China Sea. Master’s Thesis, Inha University, Incheon, Korea, 1995. [Google Scholar]
- Yoon, Y.H.; Park, J.S.; Soh, H.Y.; Hwang, D.J. Spatial distribution of phytoplankton community and red tide of Dinoflagellate, Prorocentrum donghaiense in the East China Sea during early summer. Korean J. Environ. Biol. 2003, 21, 132–141. [Google Scholar]
- Yoon, Y.H.; Park, J.S.; Park, Y.G.; Soh, H.Y.; Hwang, D.J. A characteristics of thermohaline structure and phytoplankton community from southwestern parts of the East China Sea during early summer, 2004. Bull. Korean Soc. Fish. 2005, 41, 129–139. [Google Scholar]
- Noh, J.H.; Yoo, S.J.; Lee, J.A.; Kim, H.C.; Lee, J.H. Phytoplankton in the waters of the Ieodo Ocean Research Station determined by microscopy, flow cytometry, HPLC pigment data and remote sensing. Ocean Polar Res. 2005, 27, 397–417. [Google Scholar]
- Kim, D.S.; Shim, J.H.; Lee, J.A.; Kang, Y.C. The distribution of nutrients and chlorophyll in the northern East China Sea during the spring and summer. Ocean Polar Res. 2005, 27, 251–263. [Google Scholar] [CrossRef]
- Kim, D.S.; Kim, K.H.; Shim, J.H.; Yoo, S.J. The distribution and interannual variation in nutrients, chlorophyll-a, and suspended solids in the northern East China Sea during the summer. Ocean Polar Res. 2007, 29, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Son, Y.B.; Ryu, J.H.; Noh, J.H.; Ju, S.J.; Kim, S.H. Climatological variability of satellite-derived sea surface temperature and chlorophyll in the south sea of Korea and East China Sea. Ocean Polar Res. 2012, 34, 201–218. [Google Scholar] [CrossRef] [Green Version]
- Gong, G.C.; Chang, J.; Chiang, K.P.; Hsiung, T.M.; Hung, C.C.; Duan, S.W.; Codispti, L.A. Reduction of primary production and changing of nutrient ratio in the East China Sea: Effect of the Three Gorges Dam? Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef] [Green Version]
- Jiaoa, N.; Zhang, Y.; Zeng, Y.; Gardner, W.D.; Mishonov, A.V.; Richardson, M.J.; Hong, N.; Pan, D.; Yan, X.H.; Jo, Y.H.; et al. Ecological anomalies in the East China Sea: Impacts of the Three Gorges Dam? Water Res. 2007, 41, 1287–1293. [Google Scholar] [CrossRef]
- Smayda, T.J. Biogeographical meaning indicators. In Phytoplankton Manual; Sournia, A., Ed.; United Nations Educational, Scientific, and Cultural Organization: Paris, France, 1978; pp. 225–229. [Google Scholar]
- Rines, J.E.B.; Hargraves, P.E. The Chaetoceros Ehrenberg (Bacillariophyceae) Flora of Narragansett Bay; Schweizebart Science: Stuttgart, Germany, 1988. [Google Scholar]
- Round, F.E.; Crawfor, R.M.; Mann, D.G. The Diatoms; Cambridge: New York, NY, USA, 1990. [Google Scholar]
- Shim, J.H. Flora and fauna of Korea. In Marine Phytoplankton; Ministry of Education: Seoul, Korea, 1994; Volume 34. [Google Scholar]
- Tomas, C.R. Identifying Marine Phytoplankton; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Biological and Chemical Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984. [Google Scholar]
- Oh, H.J. Ecological Characteristics of Phytoplankton in the Northern Part of East China Sea. Ph.D. Thesis, Pusan National University, Busan, Korea, 1998. [Google Scholar]
- Lin, C.; Ning, X.; Su, J.; Lin, Y.; Xu, B. Environmental changes and the responses of the ecosystems of the Yellow Sea during 1976–2000. J. Mar. Syst. 2005, 55, 223–234. [Google Scholar] [CrossRef]
- Li, M.K.; Xu, K.; Watanabe, M.; Chen, Z. Long-term variations in dissolved silicate, nitrogen, and phosphorus flux from the Changjiang River into the East China Sea and impacts on estuarine ecosystem. Estuar. Coast. Shelf Sci. 2007, 71, 3–12. [Google Scholar] [CrossRef]
- Li, H.M.; Tang, H.J.; Shi, X.Y.; Zhang, C.S.; Wang, X.L. Increased nutrient loads from the Changjiang (Yangtze) River have led to increased Harmful Algal Blooms. Harmful Algae 2014, 39, 92–101. [Google Scholar] [CrossRef]
- Ministry of Land, Transport and Maritime Affairs. The Study of Oceanographic Environmental Impact in the South Sea (East China Sea) due to the Three Gorges Dam; Ministry of Land, Transport and Maritime Affairs: Seoul, Korea, 2009; pp. 7–175.
- Jiang, Z.; Liu, J.; Chen, J.; Chen, Q.; Yan, X.; Xuan, J.; Zeng, J. Responses of summer phytoplankton community to drastic environmental changes in the 664 Changjiang (Yangtze River) estuary during the past 50 years. Water Res. 2014, 54, 1–11. [Google Scholar] [CrossRef]
- Youn, S.C.; Suk, H.Y.; Whang, J.D.; Suh, Y.S.; Yoon, Y.Y. Long-term variation in ocean environmental conditions of the northern East China Sea. J. Korean Soc. Mar. Environ. Energy 2015, 18, 189–206. [Google Scholar] [CrossRef]
- Zhou, M.J.; Shen, Z.L.; Yu, R.C. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River. Cont. Shelf Res. 2008, 28, 1483–1489. [Google Scholar] [CrossRef]
- Lee, Y.J. Phytoplankton Dynamics and Primary Production in the Yellow Sea during Winter and Summer. Ph.D. Thesis, Inha University, Incheon, Korea, 2012. [Google Scholar]
- Dortch, Q.; Whitledge, T.E. Does nitrogen or silicon limit phytoplankton production in the Mississippi River plume and nearby regions? Cont. Shelf Res. 1992, 12, 1293–1309. [Google Scholar] [CrossRef]
- Chen, Y.L.; Lu, H.; Shiah, F.; Gong, G.; Liu, K.; Kanda, J. New production and f-ratio on the continental shelf of the East China Sea: Comparisons between nitrate inputs from the subsurface Kuroshio Current and the Changjiang River. Estuar. Coast. Shelf Sci. 1999, 48, 59–75. [Google Scholar] [CrossRef]
- Magazzu, G.; Decembrini, F. Primary production, biomass and abundance of phototrophic picoplankton in the Mediterranean Sea: A review. Aquat. Microb. Ecol. 1995, 9, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Uitz, J.; Stramski, D.; Gentili, B.; D’Ortenzio, F.; Claustre, H. Estimates of phytoplankton class-specific and total primary production in the Mediterranean Sea from satellite ocean color observations. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Duarte, C.M.; Agusti, S. Nutrient and temperature control of the contribution of picoplankton to phytoplankton biomass and production. Limnol. Oceanogr. 2000, 45, 591–600. [Google Scholar] [CrossRef]
- Zohary, T.; Brenner, S.; Krom, M.D.; Angel, D.L.; Kress, N.; Li, W.K.W.; Neori, A.; Yacobi, Y.Z. Buildup of microbial biomass during deep winter mixing in a Mediterranean warm-core eddy. Mar. Ecol. Prog. Ser. 1998, 167, 47–57. [Google Scholar] [CrossRef]
- Decembrini, F.; Caroppo, C.; Azzaro, M. Size structure and production of phytoplankton community and carbon pathways channeling in the southern Tyrrhenian Sea (western Mediterranean). Deep Sea Res. Part II 2009, 56, 687–699. [Google Scholar] [CrossRef]
- Kwak, J.H.; Lee, S.H.; Hwan, J.; Suh, Y.-S.; Park, H.J.; Chang, K.-I.; Kim, K.-R.; Kang, C.K. Summer primary productivity and phytoplankton community composition driven by different hydrographic structures in the East/Japan Sea and the Western Subarctic Pacific. J. Geophys. Res. Oceans 2014, 119, 4505–4519. [Google Scholar] [CrossRef]
- Revelante, N.; Gilmartin, M. The relative increase of larger phytoplankton in a subsurface chlorophyll maximum of the northern Adriatic Sea. J. Plankton Res. 1995, 17, 1535–1562. [Google Scholar] [CrossRef]
- Marañón, E.; Holligan, P.M.; Barciela, R.; González, N.; Mouriño, B.; Pazó, M.J.; Varela, M. Patterns of phytoplankton size structure and productivity in contrasting open-ocean environments. Mar. Ecol. Prog. Ser. 2001, 216, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Morán, X.A.G.; Taupier-Letage, I.; Vázquez-Domínguez, E.; Ruiz, S.; Arin, L.; Raimbault, P.; Estrada, M. Physical-biological coupling in the Algerian Basin (SW Mediterranean): Influence of mesoscale instabilities on the biomass and production of phytoplankton and bacterioplankton. Deep Sea Res. Part I 2001, 48, 405–437. [Google Scholar] [CrossRef]
- Ning, X.; Chai, F.; Xue, H.; Cai, Y.; Liu, C.; Shi, J. Physical-biological oceanographic coupling influencing phytoplankton and primary production in the South China Sea. J. Geophys. Res. 2005, 109, C10005. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Joo, H.T.; Lee, J.H.; Lee, J.H.; Kang, J.J.; Lee, H.W.; Lee, D.B.; Kang, C.K. Carbon uptake rates of phytoplankton in the Northern East/Japan Sea. Deep Sea Res. Part II 2017, 143, 45–53. [Google Scholar] [CrossRef]
- Furuya, K.; Hayashi, M.; Yabushita, Y.; Ishikawa, A. Phytoplankton dynamics in the East China Sea in spring and summer as revealed by HPLC derived pigment signatures. Deep Sea Res. Part II 2003, 50, 367–387. [Google Scholar] [CrossRef]
- Raven, J.R. The twelfth tansley lecture. Small is beautiful: The picophytoplankton. Funct. Ecol. 1998, 12, 503–513. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A.; Schofield, O.M.; Falkowski, P.G. The role of functional traits and trade-offs in structuring phytoplankton communities: Scaling from cellular to ecosystem level. Ecol. Lett. 2007, 10, 1170–1181. [Google Scholar] [CrossRef]
- Longhurst, A.R. Ecological Geography of the Sea; Academic Press: London, UK, 2010. [Google Scholar]
- Joo, H.T.; Son, S.H.; Park, J.W.; Kang, J.J.; Jeong, J.Y.; Lee, C.I.; Kang, C.K.; Lee, S.H. Long-term pattern of primary productivity in the East/Japan Sea based on ocean color data derived from MODIS-aqua. Remote Sens. 2016, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.J. In hot water: Zooplankton and climate change. ICES J. Mar. Sci. 2008, 65, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.R.; Kang, H.K.; Choi, K.H. Latitudinal distribution of mesozooplankton community in the northwestern Pacific Ocean. Ocean Polar Res. 2011, 33, 337–347. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Latitude | Longitude | Bottom Depth (m) |
|---|---|---|---|
| 315-13 | 32.5 | 127.0 | 125 |
| 315-15 | 32.5 | 126.5 | 107 |
| 315-17 | 32.5 | 125.9 | 91 |
| 315-19 | 32.5 | 125.3 | 68 |
| 315-21 | 32.5 | 124.5 | 46 |
| 316-13 | 32.0 | 127.0 | 119 |
| 316-15 | 32.0 | 126.5 | 99 |
| 316-17 | 32.0 | 125.9 | 76 |
| 316-19 | 32.0 | 125.3 | 55 |
| 316-21 | 32.0 | 124.5 | 39 |
| 317-13 | 31.5 | 127.0 | 105 |
| 317-15 | 31.5 | 126.5 | 88 |
| 317-17 | 31.5 | 125.9 | 67 |
| 317-19 | 31.5 | 125.3 | 54 |
| 317-21 | 31.5 | 124.5 | 46 |
| Dominance | 2016 | 2017 | 2018 | 2019 | 2020 |
|---|---|---|---|---|---|
| 1st | Nanoflagellates (<20 µm; 50.5%) | Nanoflagellates (<20 µm; 50.1%) | Nanoflagellates (<20 µm; 43.1%) | Nanoflagellates (<20 µm; 68.6%) | Nanoflagellates (<20 µm; 69.7%) |
| 2nd | Scrippsiella acuminata (15.2%) | Chaetoceros lorenzianus (14.3%) | Paralia sulcata (10.2%) | Alexandrium spp. (8.4%) | Paralia sulcata (6.5%) |
| 3rd | Guinardia flaccida (9.4%) | Diploneis spp. (11.0%) | Chaetoceros spp. (6.0%) |
| 315 Line | Chl-a Size Composition (%) | 316 Line | Chl-a Size Composition (%) | 317 Line | Chl-a Size Composition (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| St.* | Depth (m) | M | N | P | St. | Depth (m) | M | N | P | St. | Depth (m) | M | N | P |
| 13 | 0 | 12.0 | 18.4 | 69.6 | 13 | 0 | 12.6 | 14.0 | 73.4 | 13 | 0 | 20.2 | 15.0 | 64.8 |
| 10 | 10.7 | 17.4 | 71.9 | 10 | 9.3 | 13.7 | 77.0 | 10 | 16.8 | 17.0 | 66.2 | |||
| 20 | 5.2 | 18.6 | 76.2 | 20 | 17.0 | 16.1 | 66.9 | 20 | 16.6 | 18.9 | 64.5 | |||
| 30 | 10.5 | 21.2 | 68.3 | 30 | 10.6 | 28.8 | 60.6 | 30 | 12.6 | 17.0 | 70.4 | |||
| 50 | 13.2 | 21.9 | 64.9 | 50 | 11.3 | 19.8 | 68.9 | 50 | 17.8 | 25.3 | 56.9 | |||
| 75 | 18.4 | 31.5 | 50.1 | 75 | 13.7 | 27.6 | 58.7 | 75 | 18.6 | 30.3 | 51.1 | |||
| 100 | 18.2 | 44.5 | 37.3 | 100 | 16.7 | 42.0 | 41.3 | 100 | 23.9 | 42.3 | 33.8 | |||
| 15 | 0 | 14.3 | 32.6 | 53.1 | 15 | 0 | 17.9 | 22.3 | 59.8 | 15 | 0 | 12.4 | 22.9 | 64.7 |
| 10 | 8.6 | 23.9 | 67.5 | 10 | 11.9 | 21.0 | 67.1 | 10 | 24.3 | 17.3 | 58.4 | |||
| 20 | 14.2 | 18.5 | 67.3 | 20 | 2.9 | 22.2 | 74.9 | 20 | 20.5 | 21.7 | 57.8 | |||
| 30 | 25.8 | 21.0 | 53.2 | 30 | 15.0 | 29.5 | 55.5 | 30 | 23.2 | 27.5 | 49.3 | |||
| 50 | 12.1 | 22.6 | 65.3 | 50 | 19.9 | 47.2 | 32.9 | 50 | 13.5 | 41.7 | 44.8 | |||
| 75 | 10.0 | 33.9 | 56.1 | 75 | 13.0 | 58.2 | 28.8 | 75 | 23.1 | 50.9 | 26.0 | |||
| 100 | 12.7 | 52.5 | 34.8 | 100 | 17.7 | 58.9 | 23.4 | □ | □ | □ | ||||
| 17 | 0 | 17.5 | 20.0 | 62.5 | 17 | 0 | 40.8 | 18.5 | 40.7 | 17 | 0 | 30.6 | 14.5 | 54.9 |
| 10 | 19.8 | 21.1 | 59.1 | 10 | 32.5 | 17.3 | 50.2 | 10 | 26.0 | 18.3 | 55.7 | |||
| 20 | 10.0 | 29.6 | 60.4 | 20 | 24.1 | 20.9 | 55 | 20 | 40.7 | 16.7 | 42.6 | |||
| 30 | 13.7 | 35.8 | 50.5 | 30 | 27.3 | 30.9 | 41.8 | 30 | 40.7 | 22.3 | 37.0 | |||
| 50 | 9.9 | 47.8 | 42.3 | 50 | 24.8 | 51.2 | 24.0 | 50 | 24.3 | 45.6 | 30.1 | |||
| 75 | 26.9 | 52.8 | 20.3 | 75 | 23.7 | 59.9 | 16.4 | □ | □ | □ | ||||
| 19 | 0 | 23.8 | 19.4 | 56.8 | 19 | 0 | 37.2 | 23.1 | 39.7 | 19 | 0 | 30.9 | 21.2 | 47.9 |
| 10 | 20.7 | 21.4 | 57.9 | 10 | 34.7 | 21.2 | 44.1 | 10 | 27.3 | 18.3 | 54.4 | |||
| 20 | 23.4 | 27.5 | 49.1 | 20 | 27.1 | 28.3 | 44.6 | 20 | 28.8 | 19.9 | 51.3 | |||
| 30 | 8.9 | 40.6 | 50.5 | 30 | 37.1 | 40.3 | 22.6 | 30 | 44.2 | 30.5 | 25.3 | |||
| 50 | 20.8 | 58.5 | 20.7 | 50 | 25.6 | 55.7 | 18.7 | 50 | 19.6 | 47.0 | 33.4 | |||
| 21 | 0 | 29.9 | 15.8 | 54.3 | 21 | 0 | 53.6 | 18.1 | 28.3 | 21 | 0 | 34.4 | 29.6 | 36.0 |
| 10 | 41.9 | 19.7 | 38.4 | 10 | 45.8 | 17.7 | 36.5 | 10 | 30.6 | 31.9 | 37.5 | |||
| 20 | 40.4 | 32.3 | 27.3 | 20 | 47.1 | 23.1 | 29.8 | 20 | 54.1 | 16.8 | 29.1 | |||
| 30 | 51.4 | 29.6 | 19.0 | 30 | 35.5 | 34.5 | 30.0 | 30 | 42.8 | 25.0 | 32.2 | |||
| Mean | 18.8 | 29.3 | 51.9 | Mean | 24.4 | 30.4 | 45.2 | Mean | 26.6 | 26.1 | 47.3 | |||
| SD | 10.9 | 12.1 | 16.0 | SD | 12.9 | 14.7 | 18.3 | SD | 10.6 | 10.7 | 13.7 | |||
| Parameters | August 1998 | August 2016–2020 | |
|---|---|---|---|
| Temperature (°C) | Surface | 27.4–29.4 (28.7 ± 0.5) | 25.4–31.3 (28.0 ± 1.4) |
| Vertical | 12.7–29.4 (21.5 ± 5.9) | 11.8–31.3 (23.1 ± 5.5) | |
| Salinity | Surface | 26.7–29.7 (28.3 ± 0.9) | 26.2–34.0 (30.1 ± 1.9) |
| Vertical | 27.5–34.7 (31.2 ± 2.5) | 26.2–34.6 (31.7 ± 1.9) | |
| Nitrite (µM) | Surface | 0.07–0.36 (0.21 ± 0.08) | 0.01–0.16 (0.05 ± 0.04) |
| Vertical | 0.07–0.57 (0.19 ± 0.14) | 0.01–1.61 (0.15 ± 0.26) | |
| Nitrate (µM) | Surface | 1.26–3.54 (2.51 ± 0.66) | 0.12–11.37 (2.77 ± 2.70) |
| Vertical | 0.87–3.54 (2.05 ± 0.74) | 0.09–35.40 (5.87 ± 5.54) | |
| Phosphate (µM) | Surface | 0.16–0.45 (0.25 ± 0.09) | ND–0.16 (0.05 ± 0.04) |
| Vertical | 0.16–1.03 (0.39 ± 0.23) | ND–0.82 (0.21 ± 0.24) | |
| Silicate (µM) | Surface | 8.95–13.77 (9.69 ± 1.45) | 0.01–11.46 (4.23 ± 3.10) |
| Vertical | 8.95–13.77 (9.59 ± 0.88) | 0.01–40.17 (7.87 ± 5.71) | |
| Dominant species | Pseudonitzschia pungens Prorocentrum dentatum Skeleonema costatum | Nanoflagellates (<20 µm) Scrippsiella acuminata Paralia sulcata |
| Relative Ratio (%) | |||||
|---|---|---|---|---|---|
| Area | Date | Pico Size | Nano Size | Micro Size | References |
| Northen East China Sea | 2018–2020/ seasonally | 45.6 | 31.2 | 23.2 | This study |
| Blanes Bay | 1997/summer | >50 | [55] | ||
| Levantine Basin | March 1992 | 54.3–64.2 | [56] | ||
| Tyrrhenian Sea (South) | July 2005 | 44–81 | [57] | ||
| December 2005 | 76–90 | ||||
| Western Subarctic Pacific | 23–29 June 2010 | 63 | [58] | ||
| Japan Basin | 5–11 July 2010 | 56 | |||
| Yamato Basin | 18–20 July 2010 | 56 | |||
| Ulleung Basin | 22–24 July 2010 | 38 | |||
| Mediterranean Sea | 31–92 | [53] | |||
| Adriatic Sea (North) | August 1986 and 1988; July 1987 | 10–23 | [59] | ||
| Atlantic Meridional Transect (Oligortophic) | April, October 1996 April, October 1997 | 80 | 16 | 4 | [60] |
| Algerian Basin | October 1996 | 42–62 | 38–58 | [61] | |
| South China Sea | summer 1998 | 63 | 22 | 16 | [62] |
| winter 1998 | 51 | 14 | 36 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, K.-W.; Oh, H.-J.; Moon, S.-Y.; Yoo, M.-H.; Youn, S.-H. Effects of Miniaturization of the Summer Phytoplankton Community on the Marine Ecosystem in the Northern East China Sea. J. Mar. Sci. Eng. 2022, 10, 315. https://doi.org/10.3390/jmse10030315
Park K-W, Oh H-J, Moon S-Y, Yoo M-H, Youn S-H. Effects of Miniaturization of the Summer Phytoplankton Community on the Marine Ecosystem in the Northern East China Sea. Journal of Marine Science and Engineering. 2022; 10(3):315. https://doi.org/10.3390/jmse10030315
Chicago/Turabian StylePark, Kyung-Woo, Hyun-Ju Oh, Su-Yeon Moon, Man-Ho Yoo, and Seok-Hyun Youn. 2022. "Effects of Miniaturization of the Summer Phytoplankton Community on the Marine Ecosystem in the Northern East China Sea" Journal of Marine Science and Engineering 10, no. 3: 315. https://doi.org/10.3390/jmse10030315