
A Na+/H+-Exchanger Gene from Penaeus monodon: Molecular Characterization and Expression Analysis under Ammonia Nitrogen Stress
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Ammonia Nitrogen Stress
2.3. RNA Extraction and Sequencing
2.4. PmNHE cDNA Cloning
2.5. Bioinformatic Analysis
2.6. qRT-PCR Analysis of PmNHE mRNA Expression
2.7. RNA Interference
3. Results
3.1. PmNHE Sequence Display and Bioinformatics Analysis
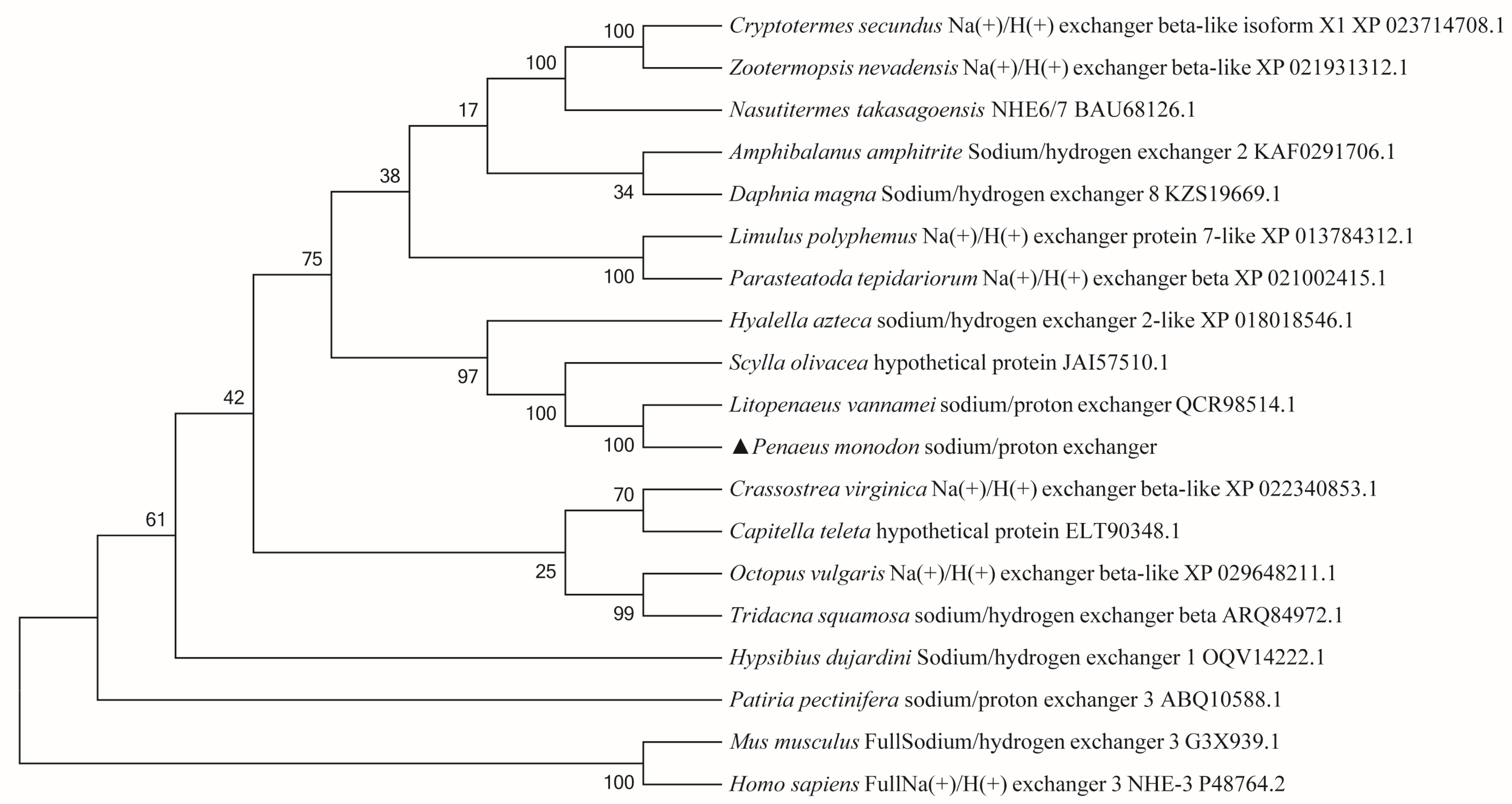
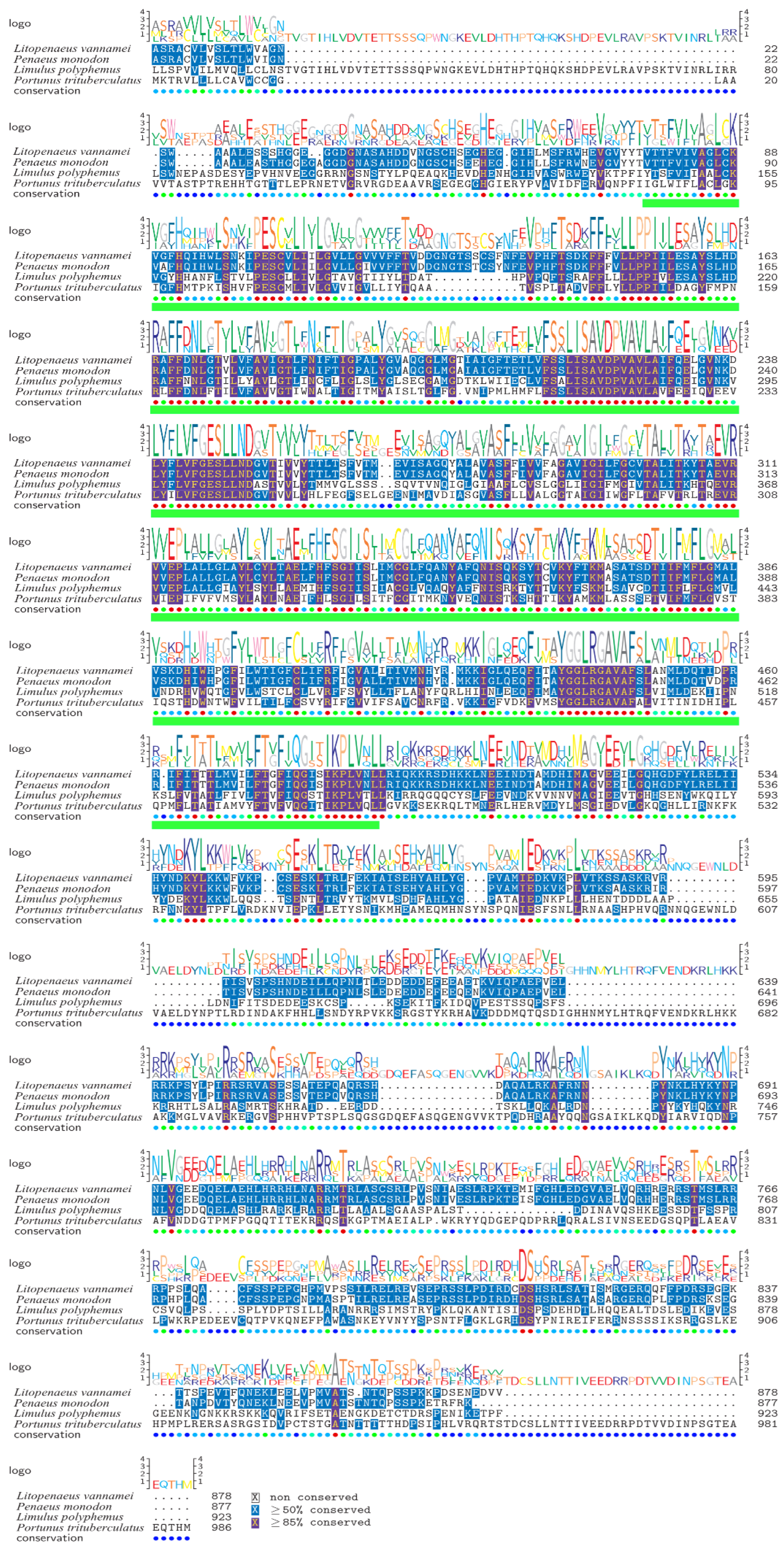
3.2. Phylogenetic Tree Analysis and Multiple Sequence Alignment
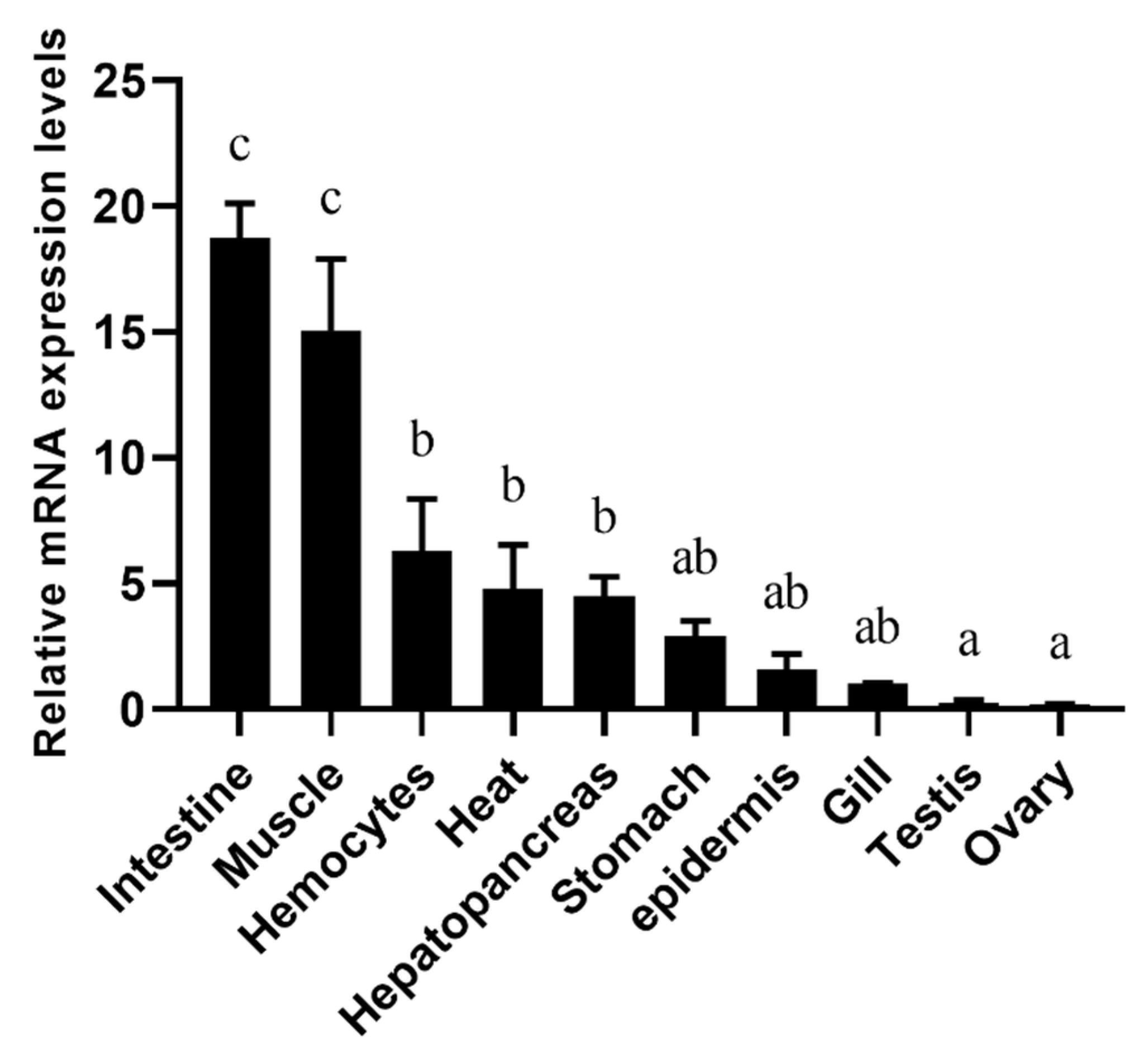
3.3. Expression Analysis of PmNHE mRNA in Different Tissues
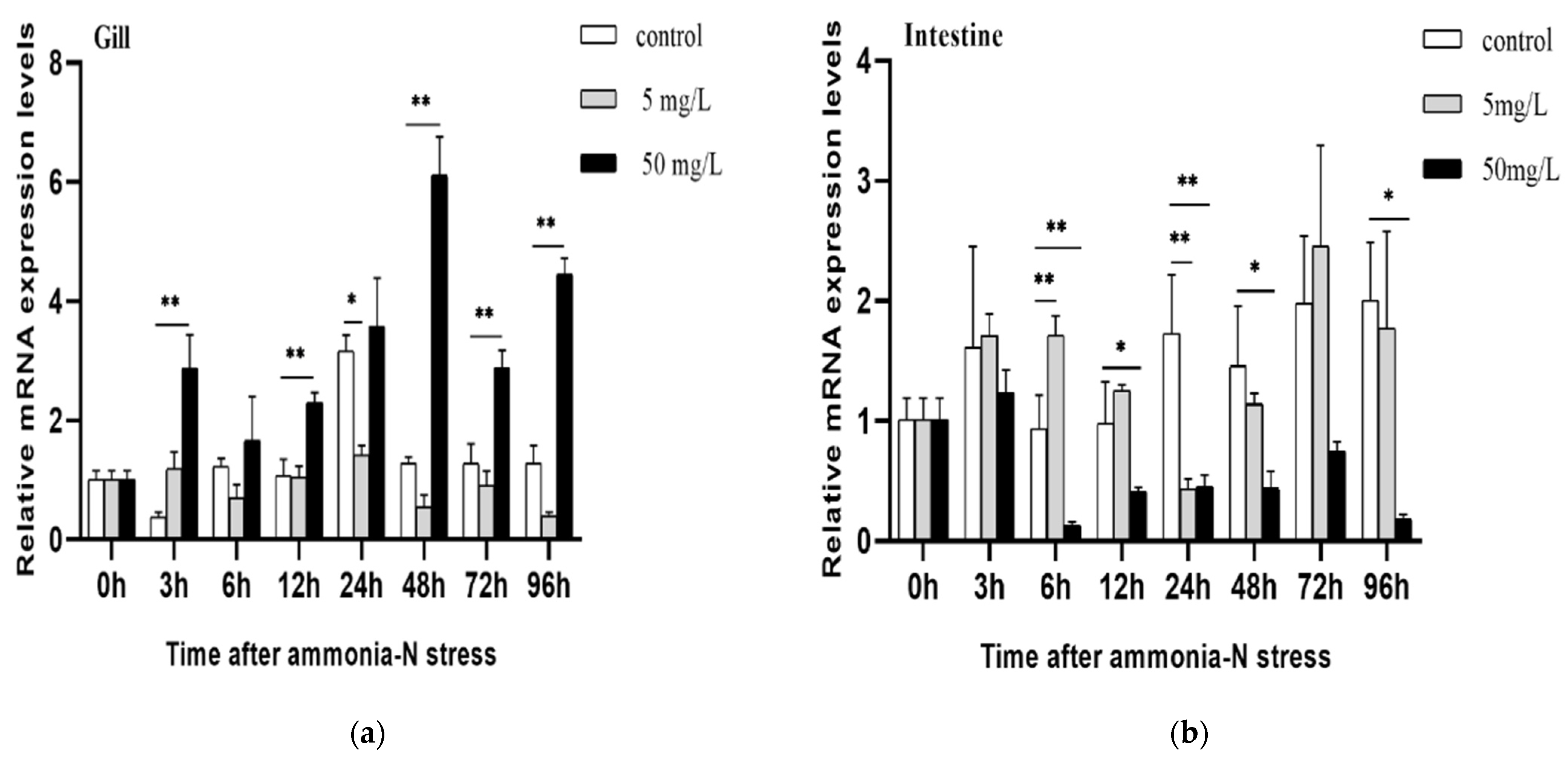
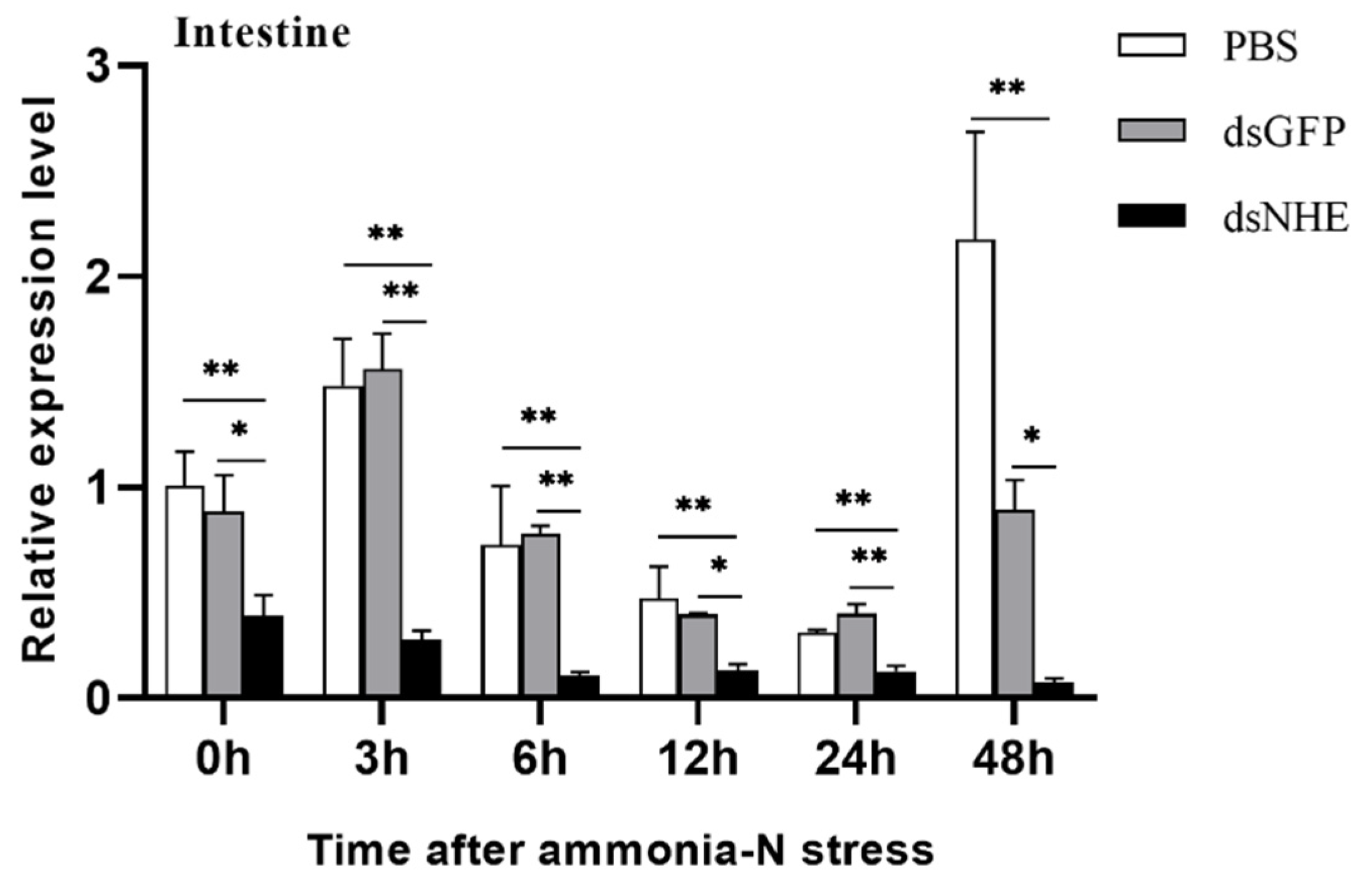
3.4. Expression Changes of PmNHE under Acute Ammonia Nitrogen Stress
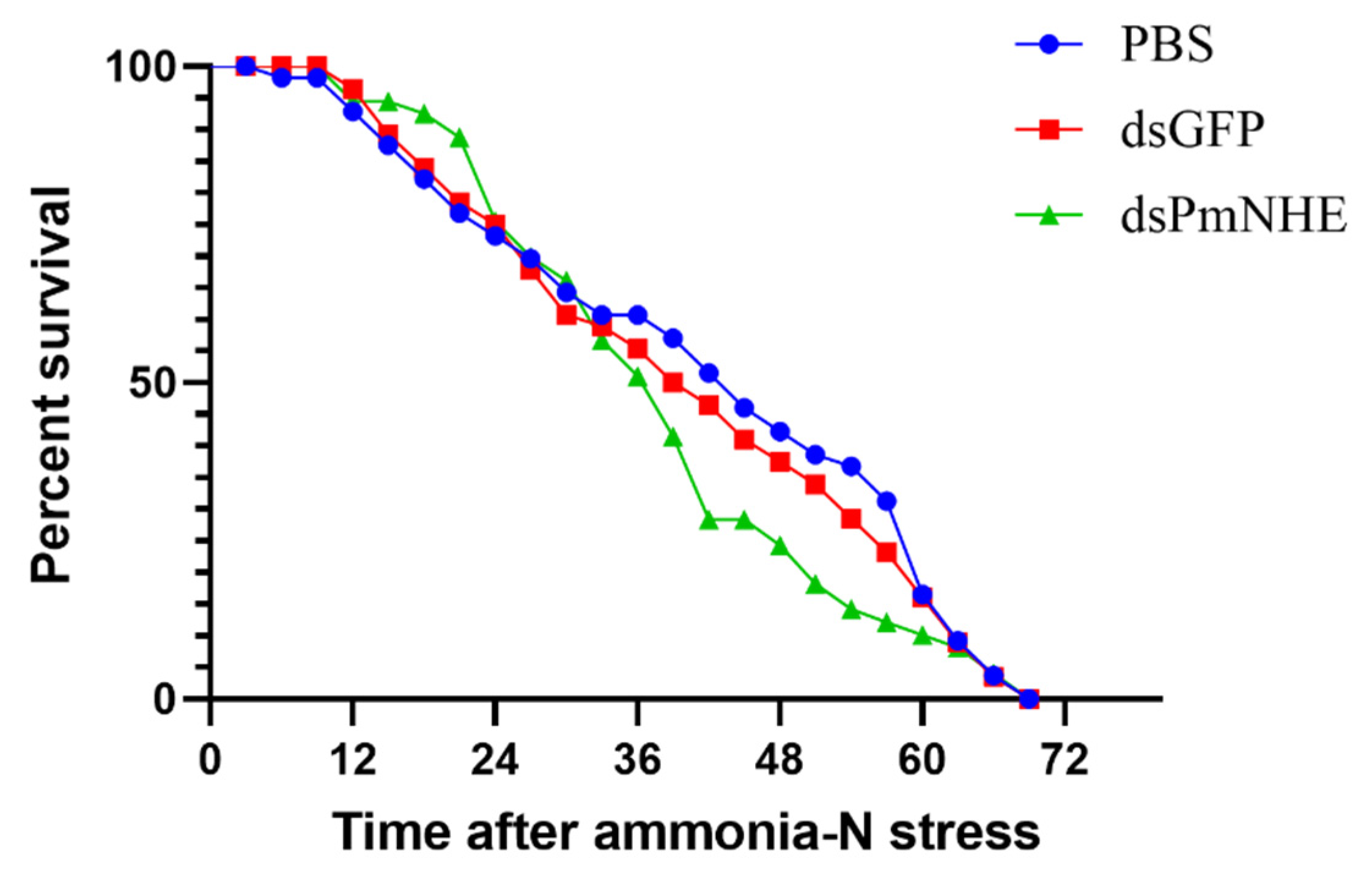
3.5. Mortality in High-Concentration Ammonia Nitrogen Environment after PmNHE Silencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hamm, L.L.; Nakhoul, N.; Hering-Smith, K.S. Acid-Base Homeostasis. Clin. J. Am. Soc. Nephrol. 2015, 10, 2232–2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, I.D.; Verlander, J.W. Ammonia Transporters and Their Role in Acid-Base Balance. Physiol. Rev. 2017, 97, 465–494. [Google Scholar] [CrossRef] [PubMed]
- Aronson, P.S.; A Suhm, M.; Nee, J. Interaction of external H+ with the Na+-H+ exchanger in renal microvillus membrane vesicles. J. Biol. Chem. 1983, 258, 6767–6771. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, J.L.; Aronson, P.S. Interaction of NH4+ and Li+ with the renal microvillus membrane Na+-H+ exchanger. Am. J. Physiol. Cell Physiol. 1981, 241, C220–C226. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.D.; Zhou, F.L.; Huang, J.H.; Yang, L.S.; Jiang, S.; Yang, Q.B.; He, J.G.; Jiang, S.G. Transcriptome reveals involvement of immune defense, oxidative imbalance, and apoptosis in ammonia-stress response of the black tiger shrimp (Penaeus monodon). Fish Shellfish. Immunol. 2018, 83, 162–170. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, F.; Yang, Q.; Jiang, S.; Huang, J.; Yang, L.; Ma, Z.; Jiang, S. Single-Cell Sequencing Reveals Types of Hepatopancreatic Cells and Haemocytes in Black Tiger Shrimp (Penaeus monodon) and Their Molecular Responses to Ammonia Stress. Front. Immunol. 2022, 13, 883043. [Google Scholar] [CrossRef]
- Lucu, Č.; Devescovi, M.; Siebers, D. Do amiloride and ouabain affect ammonia fluxes in perfused Carcinus gill epithelia? J. Exp. Zool. 1989, 249, 1–5. [Google Scholar] [CrossRef]
- Weihrauch, D.; Becker, W.; Postel, U.; Luck-Kopp, S.; Siebers, D. Potential of active excretion of ammonia in three different haline species of crabs. J. Comp. Physiol. B 1999, 169, 25–37. [Google Scholar] [CrossRef]
- Towle, D.W.; E Rushton, M.; Heidysch, D.; Magnani, J.J.; Rose, M.J.; Amstutz, A.; Jordan, M.K.; Shearer, D.W.; Wu, W.S. Sodium/proton antiporter in the euryhaline crab Carcinus maenas: Molecular cloning, expression and tissue distribution. J. Exp. Biol. 1997, 200, 1003–1014. [Google Scholar] [CrossRef]
- Li, Y.D.; Zhou, F.L.; Tang, Y.P.; Huang, J.H.; Yang, L.S.; Jiang, S.; Yang, Q.B.; Jiang, S.G. Variation in bacterial communities among stress-sensitive and stress-tolerant black tiger shrimp (Penaeus monodon) individuals. Aquac. Res. 2020, 52, 2146–2159. [Google Scholar] [CrossRef]
- Li, Y.D.; Zhou, F.L.; Huang, J.H.; Yang, L.S.; Jiang, S.; Yang, Q.B.; Jiang, S.G. Transcriptome and miRNA Profiles of Black Tiger Shrimp, Penaeus monodon, Under Different Salinity Conditions. Front. Mar. Sci. 2020, 7, 579381. [Google Scholar] [CrossRef]
- Fan, H.; Li, Y.; Yang, Q.; Jiang, S.; Yang, L.; Huang, J.; Jiang, S.; Zhou, F. Isolation and characterization of a MAPKK gene from Penaeus monodon in response to bacterial infection and low-salinity challenge. Aquac. Rep. 2021, 20, 100671. [Google Scholar] [CrossRef]
- Si, M.-R.; Li, Y.-D.; Jiang, S.-G.; Yang, Q.-B.; Jiang, S.; Yang, L.-S.; Huang, J.-H.; Chen, X.; Zhou, F.-L. A CSDE1/Unr gene from Penaeus monodon: Molecular characterization, expression and association with tolerance to low salt stress. Aquaculture 2022, 561, 738660. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Q.; Su, T.; Zhou, F.; Yang, L.; Huang, J. The toxicity of ammonia-N on Penaeus monodon and immune parameters. J. Shanghai Ocean. Univ. 2012, 21, 358–362. [Google Scholar] [CrossRef]
- Zhou, K.; Zhou, F.; Huang, J.; Yang, Q.; Jiang, S.; Qiu, L.; Yang, L.; Zhu, C.; Jiang, S. Characterization and expression analysis of a chitinase gene (PmChi-4) from black tiger shrimp (Penaeus monodon) under pathogen infection and ambient ammonia nitrogen stress. Fish Shellfish. Immunol. 2017, 62, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, F.; Fan, H.; Jiang, S.; Yang, Q.; Huang, J.; Yang, L.; Chen, X.; Zhang, W.; Jiang, S. Molecular Technology for Isolation and Characterization of Mitogen-Activated Protein Kinase Kinase 4 from Penaeus monodon, and the Response to Bacterial Infection and Low-Salinity Challenge. J. Mar. Sci. Eng. 2022, 10, 1642. [Google Scholar] [CrossRef]
- Si, M.-R.; Li, Y.-D.; Jiang, S.-G.; Yang, Q.-B.; Jiang, S.; Yang, L.-S.; Huang, J.-H.; Chen, X.; Zhou, F.-L. Identification of multifunctionality of the PmE74 gene and development of SNPs associated with low salt tolerance in Penaeus monodon. Fish Shellfish Immunol. 2022, 128, 7–18. [Google Scholar] [CrossRef]
- Tseng, Y.; Yan, J.; Furukawa, F.; Hwang, P. Did Acidic Stress Resistance in Vertebrates Evolve as Na+/H+ Exchanger-Mediated Ammonia Excretion in Fish? BioEssays 2020, 42, e1900161. [Google Scholar] [CrossRef]
- Dibrov, P.; Fliegel, L. Comparative molecular analysis of Na+/H+ exchangers: A unified model for Na+/H+ antiport? FEBS Lett. 1998, 424, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Ren, C.; Jiang, X.; Cheng, C.; Ruan, Y.; Zhang, X.; Huang, W.; Chen, T.; Hu, C.; Qiu, G.F. Na+/H+ exchanger (NHE) in Pacific white shrimp (Litopenaeus vannamei): Molecular cloning, transcriptional response to acidity stress, and physiological roles in pH homeostasis. PLoS ONE 2019, 14, e0212887. [Google Scholar] [CrossRef]
- Freire, C.A.; Onken, H.; McNamara, J.C. A structure–function analysis of ion transport in crustacean gills and excretory organs. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 272–304. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ren, C.; Wang, Y.; Gao, Y.; Wong, N.-K.; Zhang, L.; Hu, C. Crustacean cardioactive peptide (CCAP) of the Pacific white shrimp (Litopenaeus vannamei): Molecular characterization and its potential roles in osmoregulation and freshwater tolerance. Aquaculture 2016, 451, 405–412. [Google Scholar] [CrossRef]
- Mcfarland, W.; Lee, B.D. Osmotic and ionic concentrations of penaeidean shrimps of the Texas coast. Bull. Mar. Sci. 1963, 13, 391–417. [Google Scholar]
- Zimmer, A.M.; Nawata, C.M.; Wood, C.M. Physiological and molecular analysis of the interactive effects of feeding and high environmental ammonia on branchial ammonia excretion and Na+ uptake in freshwater rainbow trout. J. Comp. Physiol. B 2010, 180, 1191–1204. [Google Scholar] [CrossRef]
- Silva, J.M.; Coimbra, J.; Wilson, J. Weatherloach (Misgurnus anguillicaudatus) actively excretes ammonia through NHE. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 150, S57. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Nucleotide Sequence (5′ → 3′) | Purpose |
|---|---|---|
| 3′race GSP | TACCTCTGCTACCTCACGGCGGAACT | 3′RACE |
| 3′race NGSP | GTAGAGCTGAGAAGGAAGCCCTCGTA | 3′RACE |
| 5′race GSP | TCCGTGCTGCCCTAGAATCTCC | 5′RACE |
| 5′race NGSP | TCCGCCGTGAGGTAGCAGAGGT | 5′RACE |
| qNHE-F | TGGCTTCACAGAGACCTT | RT-PCR |
| qNHE-R | CACGCATCCGAACAGAAT | RT-PCR |
| qEF-1a-F | AAGCCAGGTATGGTTGTCAACTTT | RT-PCR |
| qEF-1a-R | CGTGGTGCATCTCCACAGACT | RT-PCR |
| dsNHE-F | TAATACGACTCACTATAGGGGCACAAGGAGGTCTAATGGG | dsRNA |
| dsNHE-R | TAATACGACTCACTATAGGGGAGGTAGCAGAGGTACGCAAG | dsRNA |
| dsGFP-F | TAATACGACTCACTATAGGGATGGCTAGCAAAGGAGAAGAACTTT | dsRNA |
| dsGFP-R | TAATACGACTCACTATAGGGAACGGGAAAAGCATTGAACACCA | dsRNA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Jiang, S.; Fan, H.; Yang, Q.; Jiang, S.; Huang, J.; Yang, L.; Zhang, W.; Chen, X.; Zhou, F. A Na+/H+-Exchanger Gene from Penaeus monodon: Molecular Characterization and Expression Analysis under Ammonia Nitrogen Stress. J. Mar. Sci. Eng. 2022, 10, 1897. https://doi.org/10.3390/jmse10121897
Li Y, Jiang S, Fan H, Yang Q, Jiang S, Huang J, Yang L, Zhang W, Chen X, Zhou F. A Na+/H+-Exchanger Gene from Penaeus monodon: Molecular Characterization and Expression Analysis under Ammonia Nitrogen Stress. Journal of Marine Science and Engineering. 2022; 10(12):1897. https://doi.org/10.3390/jmse10121897
Chicago/Turabian StyleLi, Yundong, Shigui Jiang, Hongdi Fan, Qibin Yang, Song Jiang, Jianhua Huang, Lishi Yang, Wenwen Zhang, Xu Chen, and Falin Zhou. 2022. "A Na+/H+-Exchanger Gene from Penaeus monodon: Molecular Characterization and Expression Analysis under Ammonia Nitrogen Stress" Journal of Marine Science and Engineering 10, no. 12: 1897. https://doi.org/10.3390/jmse10121897