
Estimation of Marine Macroalgal Biomass Using a Coverage Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
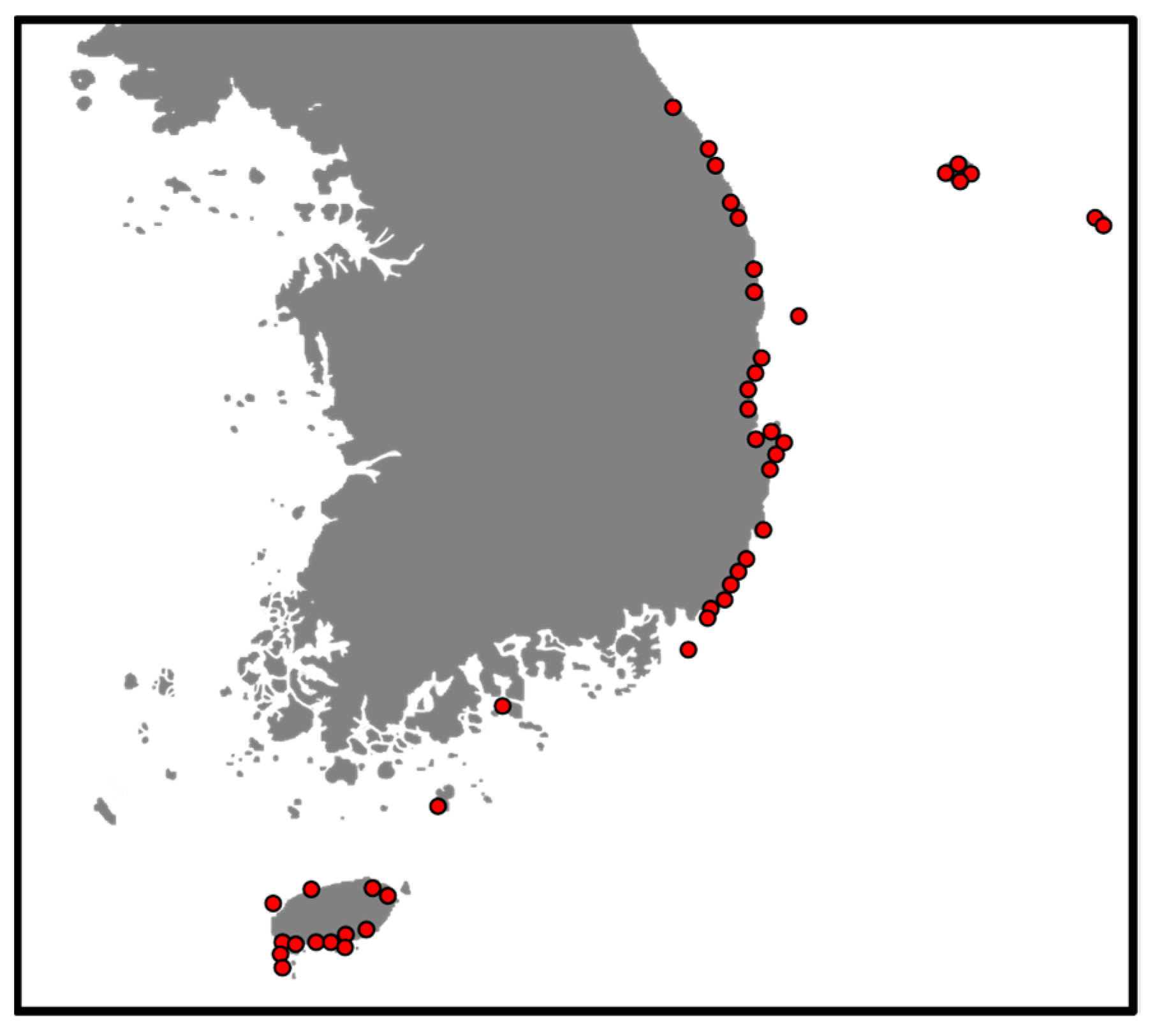
2. Materials and Methods
3. Results
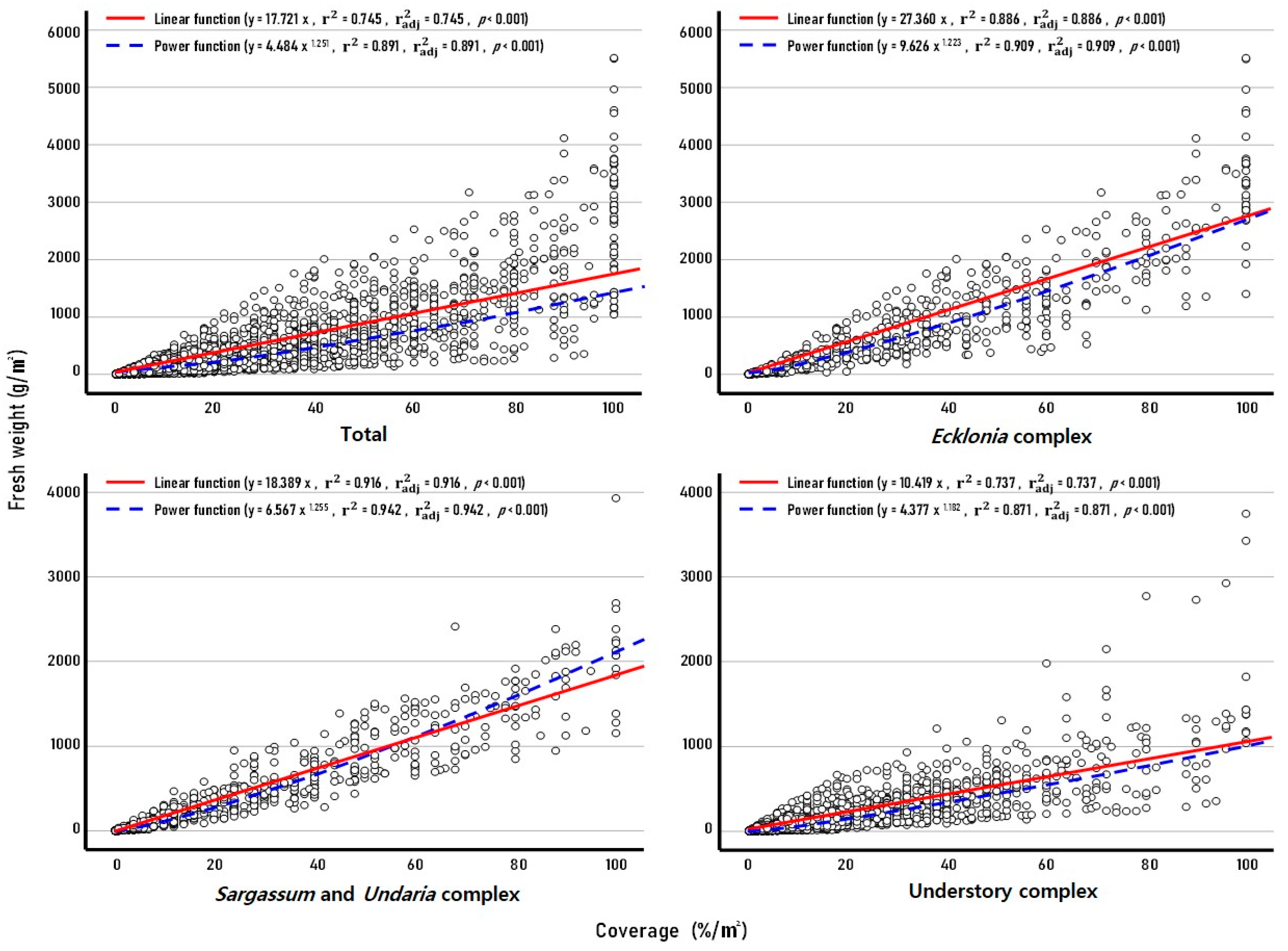
3.1. Regression of Marine Algae Data Set
3.2. Regression of Marine Algae Form (Complex)
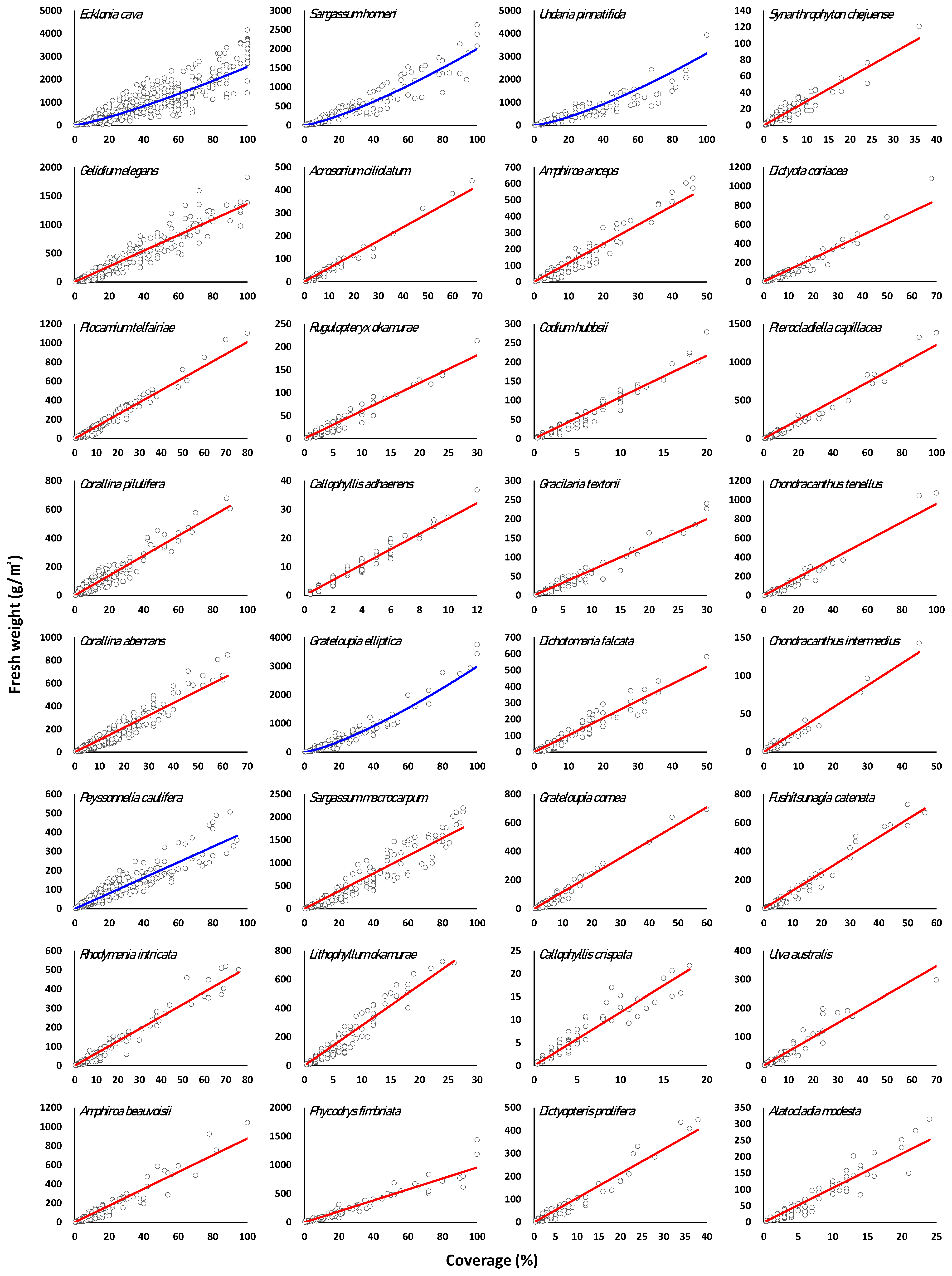
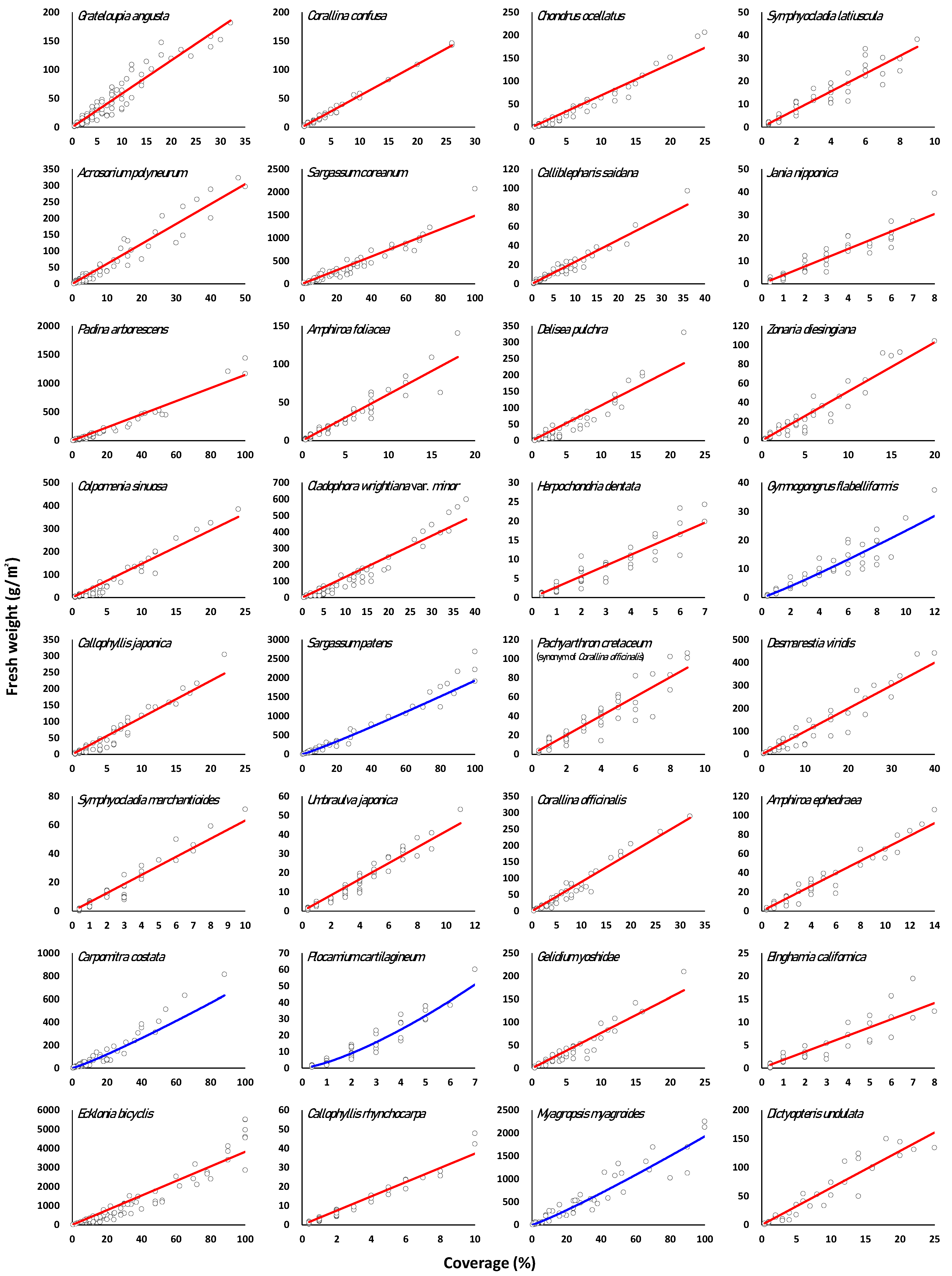
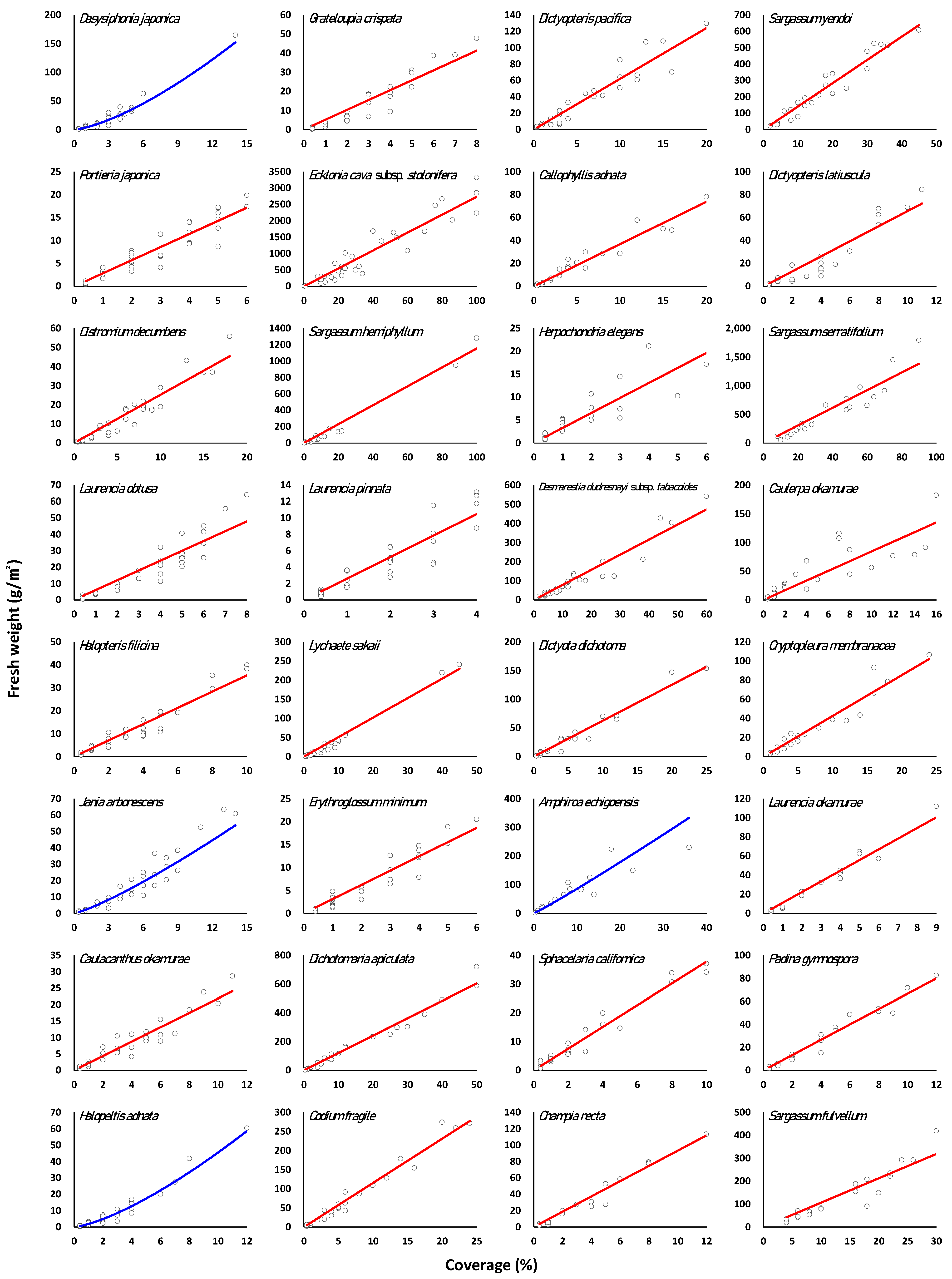
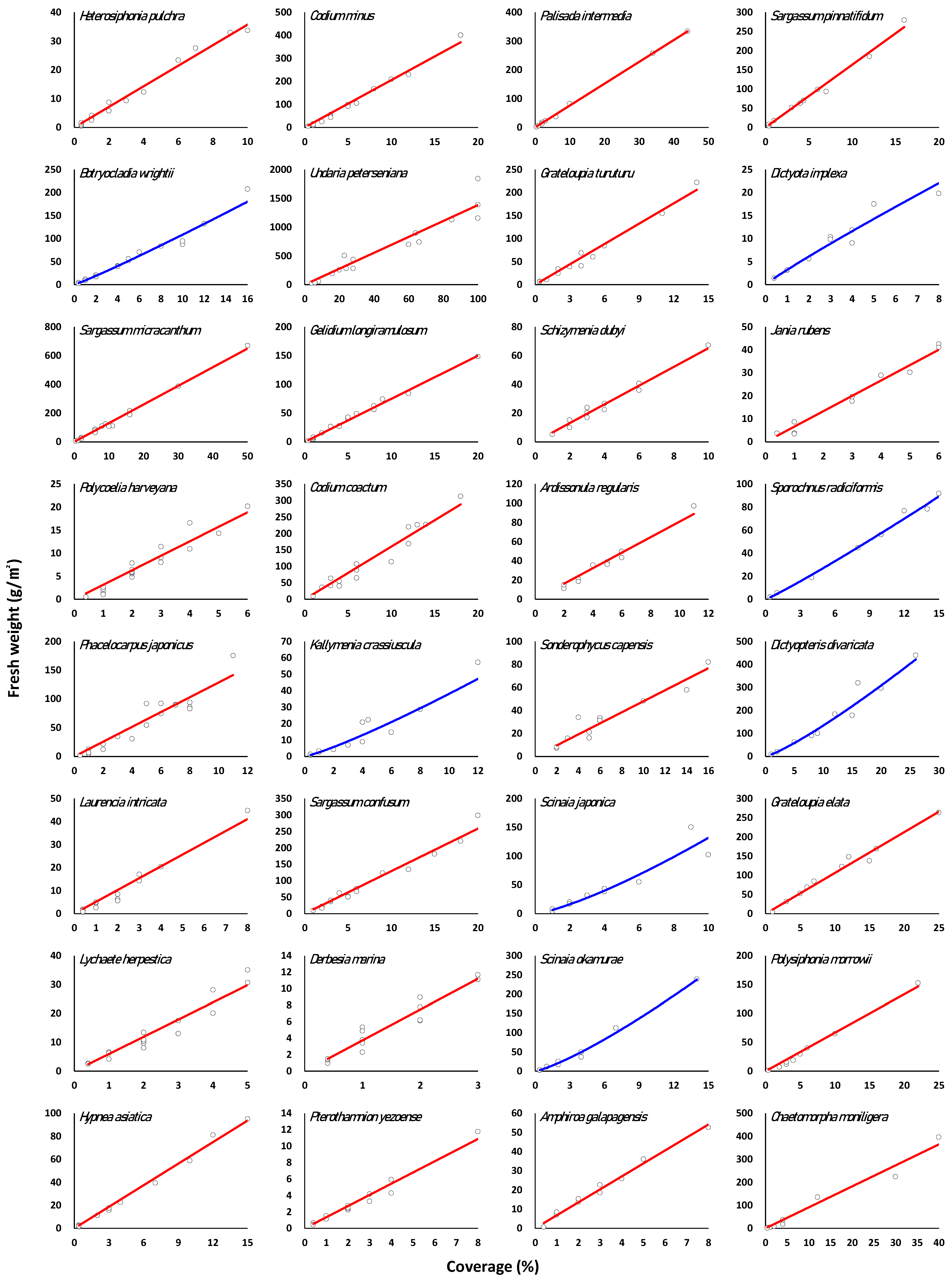
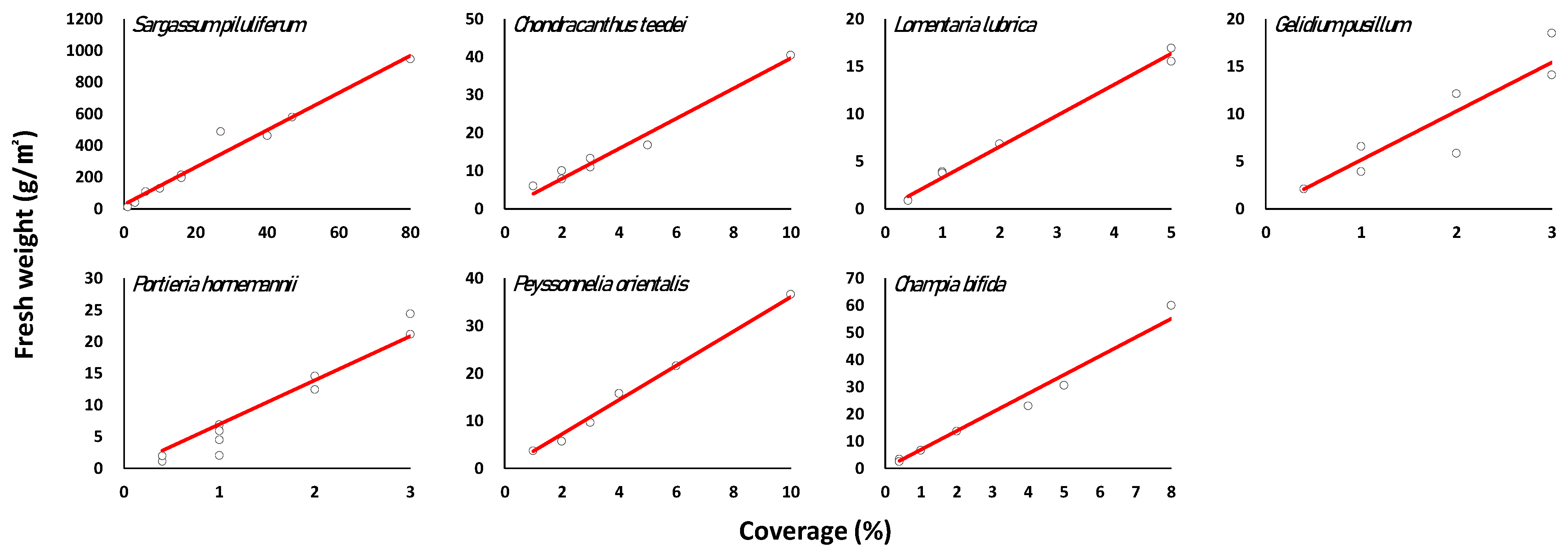
3.3. Regression of Marine Macroalgae Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Wreede, R.E. Destructive (harvest) sampling. In Handbook of Phycological Methods: Ecological Field Methods: Macroalgae; Littler, M.M., Littler, D.S., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 147–160. [Google Scholar]
- Littler, M.M.; Littler, D.S. Nondestructive sampling. In Handbook of Phycological Methods: Ecological Field Methods: Macroalgae; Cambridge University Press: Cambridge, UK, 1985; pp. 161–175. [Google Scholar]
- Gundersen, H.; Rinde, E.; Bekkby, T.; Hancke, K.; Gitmark, J.K.; Christie, H. Variation in population structure and standing stocks of kelp along multiple environmental gradients and implications for ecosystem services. Front. Mar. Sci. 2021, 8, 578629. [Google Scholar] [CrossRef]
- Mulas, M.; Silverman, J.; Rilov, G. Biomass calibration of nine dominant native and non-native Levantine seaweeds. Aquat. Bot. 2022, 178, 103496. [Google Scholar] [CrossRef]
- del Río, P.G.; Gomes-Dias, J.S.; Rocha, C.M.R.; Romaní, A.; Garrote, G.; Domingues, L. Recent trends on seaweed fractionation for liquid biofuels production. Bioresour. Technol. 2020, 299, 122613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, N.; Salama, E.-S.; Sharma, M.; Sharma, P.; Sharma, D.; Li, X. Efficient utilization and management of seaweed biomass for biogas production. Mater. Today Sustain. 2022, 18, 100120. [Google Scholar] [CrossRef]
- Ashour, M.; Alprol, A.E.; Heneash, A.M.M.; Saleh, H.; Abualnaja, K.M.; Alhashmialameer, D.; Mansour, A.T. Ammonia bioremediation from aquaculture wastewater effluents using Arthrospira platensis NIOF17/003: Impact of biodiesel residue and potential of ammonia-loaded biomass as Rotifer feed. Materials 2021, 14, 5460. [Google Scholar] [CrossRef]
- Figueira, M.M.; Volesky, B.; Ciminelli, V.S.T.; Roddick, F.A. Biosorption of metals in brown seaweed biomass. Water Res. 2000, 34, 196–204. [Google Scholar] [CrossRef]
- Znad, H.; Awual, M.R.; Martini, S. The utilization of algae and seaweed biomass for bioremediation of heavy metal-contaminated wastewater. Molecules 2022, 27, 1275. [Google Scholar] [CrossRef]
- Lerat, Y.; Cornish, M.L.; Critchley, A.T. Applications of algal biomass in global food and feed markets: From traditional usage to the potential for functional products. In Blue Biotechnology: Production and Use of Marine Molecules; Bare., S.L., Bates, S.S., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2018; pp. 143–189. [Google Scholar]
- Cotas, J.; Leandro, A.; Pacheco, D.; Gonçalves, A.M.M.; Pereira, L. A comprehensive review of the nutraceutical and therapeutic applications of red seaweeds (Rhodophyta). Life 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Abdelrhman, A.M.; Ashour, M.; Al-Zahaby, M.A.; Sharawy, Z.Z.; Nazmi, H.; Zaki, M.A.A.; Ahmed, N.H.; Ahmed, S.R.; El-Haroun, E.; Doan, H.V.; et al. Effect of polysaccharides derived from brown macroalgae Sargassum dentifolium on growth performance, serum biochemical, digestive histology and enzyme activity of hybrid red tilapia. Aquac. Rep. 2022, 25, 101212. [Google Scholar] [CrossRef]
- Ko, Y.W.; Sung, G.H.; Kim, J.H. Estimation for seaweed biomass using regression: A methodological approach. Algae 2008, 23, 289–294. [Google Scholar] [CrossRef]
- Choi, C.G.; Ohno, M.; Sohn, C.H. Algal succession on different substrata covering the artificial iron reef at Ikata in Shikoku, Japan. Algae 2006, 21, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Wernberg, T.; Coleman, M.A.; Babcock, R.C.; Bell, S.Y.; Bolton, J.J.; Connell, S.D.; Hurd, C.L.; Johnson, C.R.; Marzinelli, E.M.; Shears, N.T.; et al. Biology and ecology of the globally significant kelp Ecklonia radiata. Oceanogr. In Oceanography and Marine Biology; Oxford University Press: Oxford, UK, 2019; Volume 57, pp. 265–324. [Google Scholar]
- Layton, C.; Coleman, M.A.; Marzinelli, E.M.; Steinberg, P.D.; Swearer, S.E.; Vergés, A.; Wernberg, T.; Johnson, C.R. Kelp forest restoration in Australia. Front. Mar. Sci. 2020, 7, 74. [Google Scholar] [CrossRef] [Green Version]
- Cebrian, E.; Tamburello, L.; Verdura, J.; Guarnieri, G.; Medrano, A.; Linares, C.; Hereu, B.; Garrabou, J.; Cerrano, C.; Galobart, C.; et al. A roadmap for the restoration of mediterranean macroalgal forests. Front. Mar. Sci. 2021, 8, 709219. [Google Scholar] [CrossRef]
- Fragkopoulou, E.; Serrão, E.A.; De Clerck, O.; Costello, M.J.; Araújo, M.B.; Duarte, C.M.; Krause-Jensen, D.; Assis, J. Global biodiversity patterns of marine forests of brown macroalgae. Glob. Ecol. Biogeogr. 2022, 31, 636–648. [Google Scholar] [CrossRef]
- Mansour, A.T.; Ashour, M.; Alprol, A.E.; Alsaqufi, A.S. Aquatic plants and aquatic animals in the context of sustainability: Cultivation techniques, integration, and blue revolution. Sustainability 2022, 14, 3257. [Google Scholar] [CrossRef]
- Preskitt, L.B.; Vroom, P.S.; Smith, C.M. A rapid ecological assessment (REA) quantitative survey method for benthic algae using photoquadrats with scuba. Pac. Sci. 2004, 58, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Sedano, F.; Pavón-Paneque, A.; Navarro-Barranco, C.; Guerra-García, J.M.; Digenis, M.; Sempere-Valverde, J.; Espinosa, F. Coastal armouring affects intertidal biodiversity across the Alboran Sea (Western Mediterranean Sea). Mar. Environ. Res. 2021, 171, 105475. [Google Scholar] [CrossRef]
- Ko, Y.W.; Choi, H.-G.; Lee, D.S.; Kim, J.H. 30 years revisit survey for long-term changes in the Antarctic subtidal algal assemblage. Sci. Rep. 2020, 10, 8481. [Google Scholar] [CrossRef]
- Bravo, G.; Livore, J.P.; Bigatti, G. Monitoring rocky reef biodiversity by underwater geo-referenced photoquadrats. Underw. Technol. 2021, 38, 17–24. [Google Scholar] [CrossRef]
- Chennu, A.; Färber, P.; Death, G.; de Beer, D.; Fabricius, K.E. A diver-operated hyperspectral imaging and topographic surveying system for automated mapping of benthic habitats. Sci. Rep. 2017, 7, 7122. [Google Scholar] [CrossRef]
- Robbins, B.D.; Boese, B.L. Macroalgal volume: A surrogate for biomass in some green algae. Bot. Mar. 2002, 45, 586–588. [Google Scholar] [CrossRef]
- Kim, S.; Youn, S.-H. Estimating the individual dry weight of sheet form macroalgae for laboratory studies. J. Korean Soc. Mar. Environ. Saf. 2019, 25, 244–250. [Google Scholar] [CrossRef]
- Kim, S.; Kang, Y.H.; Kim, T.-H.; Lee, H.J.; Park, S.R. Use of morphological characteristics for calculating individual biomass in the kelp Ecklonia cava. J. Appl. Phycol. 2017, 29, 2587–2593. [Google Scholar] [CrossRef]
- Campbell, J.; Starko, S. Allometric models effectively predict Saccharina latissima (Laminariales, Phaeophyceae) fresh weight at local scales. J. Appl. Phycol. 2020, 33, 491–500. [Google Scholar] [CrossRef]
- Scrosati, R.A.; MacDonald, H.L.; Córdova, C.A.; Casas, G.N. Length and biomass data for Atlantic and Pacific seaweeds from both hemispheres. Front. Mar. Sci. 2020, 7, 592675. [Google Scholar] [CrossRef]
- Van Son, T.C.; Nikolioudakis, N.; Steen, H.; Albretsen, J.; Furevik, B.R.; Elvenes, S.; Moy, F.; Norderhaug, K.M. Achieving reliable estimates of the spatial distribution of kelp biomass. Front. Mar. Sci. 2020, 7, 107. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Zhang, J.; Cui, T.; Gong, J.; Liu, R.; Chen, X.; Liang, X. Remote sensing estimation of the biomass of floating Ulva prolifera and analysis of the main factors driving the interannual variability of the biomass in the Yellow Sea. Mar. Pollut. Bull. 2019, 140, 330–340. [Google Scholar] [CrossRef]
- Che, S.; Du, G.; Wang, N.; He, K.; Mo, Z.; Sun, B.; Chen, Y.; Cao, Y.; Wang, J.; Mao, Y. Biomass estimation of cultivated red algae Pyropia using unmanned aerial platform based multispectral imaging. Plant Methods 2021, 17, 12. [Google Scholar] [CrossRef]
- Dayton, P.K.; Currie, V.; Gerrodette, T.; Keller, B.D.; Rosenthal, R.; Ven Tresca, D. Patch dynamics and stability of some California kelp communities. Ecol. Monogr. 1984, 54, 253–289. [Google Scholar] [CrossRef]
- Benes, K.M.; Carpenter, R.C. Kelp canopy facilitates understory algal assemblage via competitive release during early stages of secondary succession. Ecology 2015, 96, 241–251. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication. National University of Ireland: Galway, Ireland. Available online: https://www.algaebase.org (accessed on 16 February 2022).
- Webster, T.; MacDonald, C.; McGuigan, K.; Crowell, N.; Lauzon-Guay, J.-S.; Collins, K. Calculating macroalgal height and biomass using bathymetric LiDAR and a comparison with surface area derived from satellite data in Nova Scotia, Canada. Bot. Mar. 2019, 63, 43–59. [Google Scholar] [CrossRef]
- Johnson, M.P. Estimating intertidal seaweed biomass at larger scales from quadrat surveys. Mar. Environ. Res. 2020, 156, 104906. [Google Scholar] [CrossRef]
- Montes-Herrera, J.C.; Cimoli, E.; Cummings, V.; Hill, N.; Lucieer, A.; Lucieer, V. Underwater hyperspectral imaging (UHI): A review of systems and applications for proximal seafloor ecosystem studies. Remote Sens. 2021, 13, 3451. [Google Scholar] [CrossRef]
- Kain (Jones), J.M. Morphology and growth of the giant kelp Macrocystis pyrifera in New Zealand and California. Mar. Biol. 1982, 67, 143–157. [Google Scholar] [CrossRef]
- Steneck, R.S.; Dethier, M.N. A functional group approach to the structure of algal-dominated communities. Oikos 1994, 69, 476–498. [Google Scholar] [CrossRef] [Green Version]
- Hwang, E.-K.; Gong, Y.-G.; Ha, D.-S.; Park, C.-S. Nursery and main culture conditions for mass cultivation of the brown alga, Ecklonia cava Kjellman. Korean J. Fish. Aquat. Sci. 2010, 43, 687–692. [Google Scholar] [CrossRef] [Green Version]
- Kawai, H.; Akita, S.; Hashimoto, K.; Hanyuda, T. A multigene molecular phylogeny of Eisenia reveals evidence for a new species, Eisenia nipponica (Laminariales), from Japan. Eur. J. Phycol. 2020, 55, 234–241. [Google Scholar] [CrossRef]
- Duarte, C.M.; Wu, J.; Xiao, X.; Bruhn, A.; Krause-Jensen, D. Can seaweed farming play a role in climate change mitigation and adaptation? Front. Mar. Sci. 2017, 4, 100. [Google Scholar] [CrossRef] [Green Version]
- Krause-Jensen, D.; Lavery, P.; Serrano, O.; Marbà, N.; Masque, P.; Duarte, C.M. Sequestration of macroalgal carbon: The elephant in the blue carbon room. Biol. Lett. 2018, 14, 20180236. [Google Scholar] [CrossRef] [Green Version]
- Raven, J. Blue carbon: Past, present and future, with emphasis on macroalgae. Biol. Lett. 2018, 14, 20180336. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Littler, M.M.; Littler, D.S. Nutrient availability to marine macroalgae in siliciclastic versus carbonate-rich coastal waters. Estuaries 1992, 15, 75–82. [Google Scholar] [CrossRef]
- Gevaert, F.; Davoult, D.; Creach, A.; Kling, R.; Janquin, M.-A.; Seuront, L.; Lemoine, Y. Carbon and nitrogen content of Laminaria saccharina in the eastern English Channel: Biometrics and seasonal variations. J. Mar. Biol. Assoc. UK 2001, 81, 727–734. [Google Scholar] [CrossRef]
- Choi, S.K.; Oh, H.-J.; Yun, S.-H.; Lee, H.J.; Lee, K.; Han, Y.S.; Kim, S.; Park, S.R. Population dynamics of the ‘golden tides’ seaweed, Sargassum horneri, on the Southwestern coast of Korea: The extent and formation of golden tides. Sustainability 2020, 12, 2903. [Google Scholar] [CrossRef] [Green Version]
- Olsson, J.; Toth, G.B.; Albers, E. Biochemical composition of red, green and brown seaweeds on the Swedish west coast. J. Appl. Phycol. 2020, 32, 3305–3317. [Google Scholar] [CrossRef]
- Filbee-Dexter, K.; Wernberg, T. Substantial blue carbon in overlooked Australian kelp forests. Sci. Rep. 2020, 10, 12341. [Google Scholar] [CrossRef] [PubMed]
- Laurens, L.M.L.; Lane, M.; Nelson, R.S. Sustainable seaweed biotechnology solutions for carbon capture, composition, and deconstruction. Trends Biotechnol. 2020, 38, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Wernberg, T.; Krumhansl, K.; Filbee-Dexter, K.; Pedersen, M.F. Status and trends for the world’s kelp forests. In World Seas: An Environmental Evaluation, 2nd ed.; Sheppard, C., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 57–78. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, S.W.; Choi, C.G. Estimation of Marine Macroalgal Biomass Using a Coverage Analysis. J. Mar. Sci. Eng. 2022, 10, 1676. https://doi.org/10.3390/jmse10111676
Jung SW, Choi CG. Estimation of Marine Macroalgal Biomass Using a Coverage Analysis. Journal of Marine Science and Engineering. 2022; 10(11):1676. https://doi.org/10.3390/jmse10111676
Chicago/Turabian StyleJung, Seung Wook, and Chang Geun Choi. 2022. "Estimation of Marine Macroalgal Biomass Using a Coverage Analysis" Journal of Marine Science and Engineering 10, no. 11: 1676. https://doi.org/10.3390/jmse10111676