
Do Phytomer Turnover Models of Plant Morphology Describe Perennial Ryegrass Root Data from Field Swards?



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Field Plots
2.2. Mechanistic Modeling of Phytomer Root Production
2.3. Statistical Analysis
3. Results
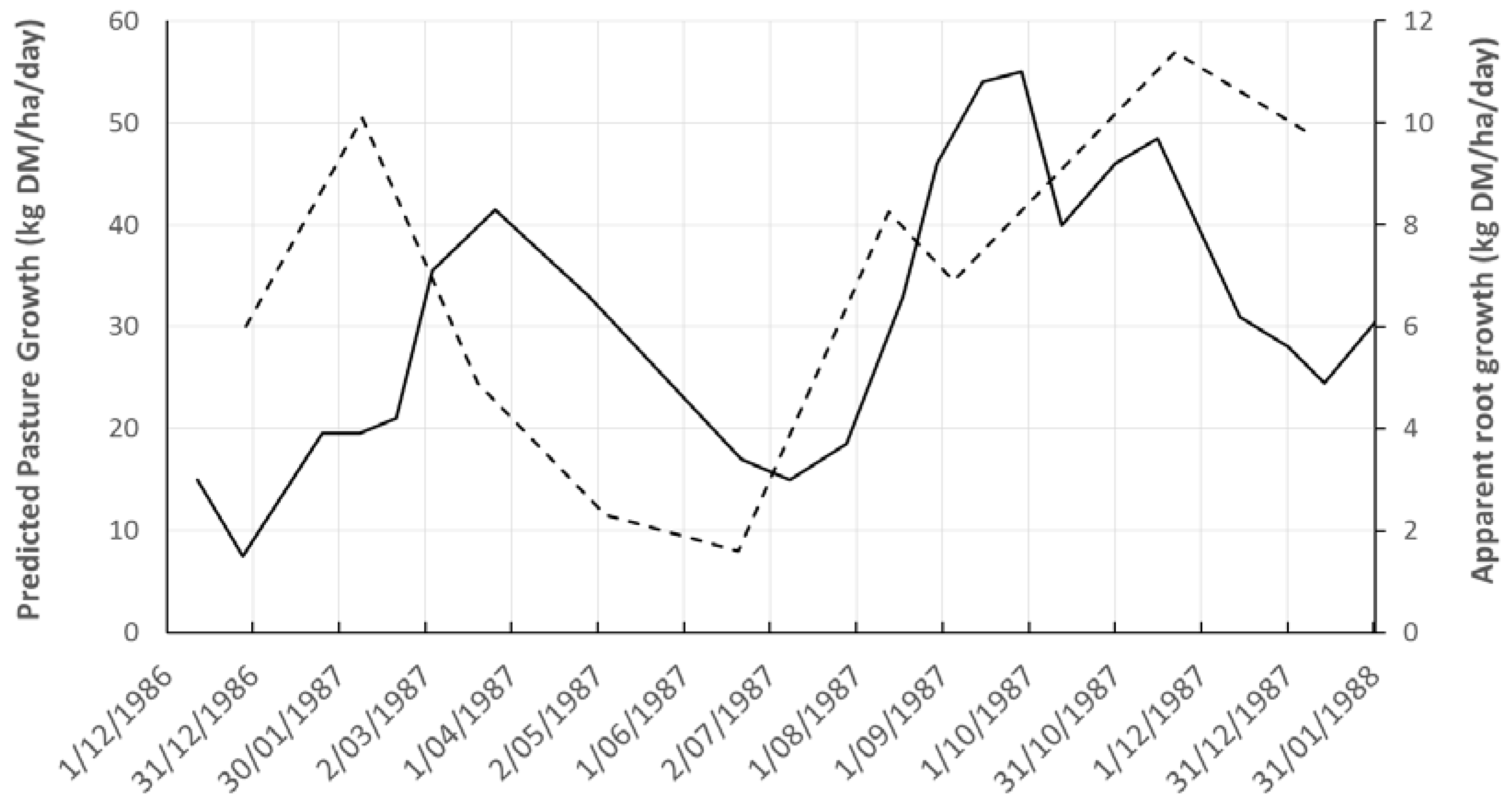
3.1. Root and Herbage Biomass Fluxes
3.2. Determination of Phyllochron (P) and Rainfall Pattern
3.3. Determination of Architectural Signal (AS), and Root Dry Weight Per Phytomer (Wr)
3.4. Relationship between Root Traits and Seasonal Change in Phyllochron (P) and Architectural Signal (AS)
4. Discussion
4.1. Ingrowth Core Data Provides Insight Complementary to that from Other Techniques
4.2. Signals Determining Seasonality of Root Deposition (Fr) and Root Diameter
4.3. Reconciliation of Fr Data from Field Swards with Phytomer Turnover Models of Root Formation
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Parsons, A.J.; Leafe, E.L.; Collett, B.; Stiles, W. The physiology of grass production under grazing. I. Characteristics of leaf and canopy photosynthesis of continuously-grazed swards. J. Appl. Ecol. 1983, 20, 117–126. [Google Scholar] [CrossRef]
- Parsons, A.J.; Leafe, E.L.; Collett, B.; Penning, P.D.; Lewis, J. The physiology of grass production under grazing. II. Photosynthesis, crop growth and animal intake of continuously-grazed swards. J. Appl. Ecol. 1983, 20, 127–139. [Google Scholar] [CrossRef]
- Korte, C.J. Tillering in ‘Grasslands Nui’perennial ryegrass swards 2. Seasonal pattern of tillering and age of flowering tillers with two mowing frequencies. N. Z. J. Agric. Res. 1986, 29, 629–638. [Google Scholar] [CrossRef]
- Garwood, E.A. Seasonal variation in appearance and growth of grass roots. Grass Forage Sci. 1967, 22, 121–129. [Google Scholar] [CrossRef]
- Chapman, D.F.; Lemaire, G. Morphogenetic and structural determinants of plant regrowth after defoliation. In Proceedings of the XVII International Grassland Congress, Palmerston North, New Zealand, 8–21 February 1993; pp. 95–104.
- Bahmani, I.; Hazard, L.; Varlet-Grancher, C.; Betin, M.; Lemaire, G.; Matthew, C.; Thom, E.R. Differences in tillering of long-and short-leaved perennial ryegrass genetic lines under full light and shade treatments. Crop Sci. 2000, 40, 1095–1102. [Google Scholar] [CrossRef]
- Van Loo, E.N. On the Relation between Tillering, Leaf Area Dynamics and Growth of Perennial Ryegrass (Lolium perenne L.). Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, May 1993. [Google Scholar]
- Sartie, A.M.; Matthew, C.; Easton, H.S.; Faville, M.J. Phenotypic and QTL analysis of herbage production-related traits in perennial ryegrass (Lolium perenne L.). Euphytica 2011, 182, 295–315. [Google Scholar] [CrossRef]
- Matthew, C.; Xia, J.; Chu, A.; Mackay, A.; Hodgson, J. Relationship between root production and tiller appearance rates in perennial ryegrass (Lolium perenne L.). In Plant Root Growth—An Ecological Perspective; Atkinson, D., Ed.; British Ecological Society Special Publication, Blackwell Scientific Publications: London, UK, 1991; pp. 281–290. [Google Scholar]
- Xia, J.X. The Effects of Defoliation on Tissue Turnover and Pasture Production in Perennial Ryegrass, Prairie Grass and Smooth Bromegrass Pasture. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 1991. [Google Scholar]
- Matthew, C. A Study of Seasonal Root and Tiller Dynamics in Swards of Perennial Ryegrass (Lolium perenne L.). Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 1993. Available online: http://mro.massey.ac.nz/xmlui/bitstream/handle/10179/3289/02_whole.pdf (accessed on 11 June 2016). [Google Scholar]
- Davidson, R.L. Root systems—The forgotten component of pastures. In Plant Relations in Pastures; Wilson, J.R., Ed.; CSIRO: Melbourne, Australia, 1978; pp. 86–94. [Google Scholar]
- Troughton, A. The Underground Organs of Herbage Grasses; Bulletin No. 44; Commonwealth Bureau of Pastures and Field Crops: Hurley, UK, 1957. [Google Scholar]
- Crush, J.R.; Nichols, S.N.; Easton, H.S.; Ouyang, L.; Hume, D.E. Comparisons between wild populations and bred perennial ryegrasses for root growth and root/shoot partitioning. N. Z. J. Agric. Res. 2009, 52, 161–169. [Google Scholar] [CrossRef]
- Crush, J.R.; Easton, H.S.; Waller, J.E.; Hume, D.E.; Faville, M.J. Genotypic variation in patterns of root distribution, nitrate interception and response to moisture stress of a perennial ryegrass (Lolium perenne L.) mapping population. Grass For. Sci. 2007, 62, 265–273. [Google Scholar] [CrossRef]
- Crush, J.R.; Popay, A.J.; Waller, J. Effect of different Neotyphodium endophytes on root distribution of a perennial ryegrass (Lolium perenne L.) cultivar. N. Z. J. Agric. Res. 2004, 47, 345–349. [Google Scholar] [CrossRef]
- Cartes, P.; Jara, A.A.; Pinilla, L.; Rosas, A.; Mora, M.L. Selenium improves the antioxidant ability against aluminium-induced oxidative stress in ryegrass roots. Ann. Appl. Biol. 2010, 156, 297–307. [Google Scholar] [CrossRef]
- Xu, W.H.; Liu, H.; Ma, Q.F.; Xiong, Z.T. Root exudates, rhizosphere Zn fractions, and Zn accumulation of ryegrass at different soil Zn levels. Pedosphere 2007, 17, 389–396. [Google Scholar] [CrossRef]
- Bircham, J.S.; Hodgson, J. The influence of sward condition on rates of herbage growth and senescence in mixed swards under continuous stocking management. Grass For. Sci. 1983, 38, 323–331. [Google Scholar] [CrossRef]
- Donaghy, D.J.; Fulkerson, W.J. Plant-soluble carbohydrate reserves and senescence—Key criteria for developing an effective grazing management system for ryegrass-based pastures: A review. Aust. J. Exp. Agric. 2001, 41, 261–275. [Google Scholar]
- Robin, A.H.K.; Matthew, C.; Crush, J. Time course of root initiation and development in perennial ryegrass—A new perspective. Proc. N. Z. Grassl. Assoc. 2010, 72, 233–239. [Google Scholar]
- Robin, A.H.K.; Uddin, M.J.; Bayazid, K.N. Polyethylene glycol (PEG)-treated hydroponic culture reduces length and diameter of root hairs of wheat varieties. Agronomy 2015, 5, 506–518. [Google Scholar] [CrossRef]
- Robin, A.H.K.; Uddin, M.J.; Afrin, S.; Paul, P.R. Genotypic variations in root traits of wheat varieties at phytomer level. J. Bangladesh Agric. Univ. 2014, 12, 45–54. [Google Scholar] [CrossRef]
- Robin, A.H.K.; Matthew, C.; Uddin, M.J.; Bayazid, K.N. Salinity-induced reduction in root surface area and changes in major root and shoot traits at the phytomer level in wheat. J. Exp. Bot. 2016. [Google Scholar] [CrossRef] [PubMed]
- Steen, E. Variation in root growth in a grass ley studied with a mesh bag technique. Swed. J. Agric. Res. 1984, 14, 93–97. [Google Scholar]
- Matthew, C.; Yang, J.Z.; Potter, J.F. Determination of tiller and root appearance in perennial ryegrass (Lolium perenne) swards by observation of the tiller axis, and potential application in mechanistic modelling. N. Z. J. Agric. Res. 1998, 41, 1–10. [Google Scholar] [CrossRef]
- Bos, H.J.; Neuteboom, J.H. Morphological analysis of leaf and tiller number dynamics of wheat (Triticum aestivum L.): Responses to temperature and light Intensity. Ann. Bot. 1998, 81, 131–139. [Google Scholar] [CrossRef]
- Hitch, P.A.; Sharman, B.C. Vascular pattern of festucoid grass axes, with particular reference to nodal plexi. Bot. Gaz. 1971, 132, 38–56. [Google Scholar] [CrossRef]
- Etter, A.G. How Kentucky bluegrass grows. Ann. MO. Bot. Gard. 1951, 38, 293–375. [Google Scholar] [CrossRef]
- Henessy, D.; O’Donovan, M.; French, P.; Laidlaw, A.S. Factors influencing tissue turnover during winter in perennial ryegrass-dominated swards. Grass For. Sci. 2008, 63, 202–211. [Google Scholar] [CrossRef]
- Caradus, J.R.; Evans, P.S. Seasonal root formation of white clover, ryegrass and cocksfoot in New Zealand. N. Z. J. Agric. Res. 1977, 20, 337–342. [Google Scholar] [CrossRef]
- Jacques, W.A. Root development in some common New Zealand pasture plants. IX. The root replacement pattern in perennial ryegrass (Lolium perenne). N. Z. J. Sci. Technol. A 1956, 38, 160–165. [Google Scholar]
- Matthew, C.; Van Loo, E.N.; Thom, E.R.; Dawson, L.A.; Care, D.A. Understanding shoot and root development. In Proceedings of the XIX International Grassland Congress, Sao Pedro, Brazil, 11–21 February 2001; pp. 19–27.
- Bohm, W. Methods of studying root systems. In Ecological Studies—Analysis and Synthesis; Billings, W.D., Golley, F., Lange, O.L., Olson, J.S., Eds.; Springer-Verlag: Berlin, Germany, 1979; Volume 33. [Google Scholar]
- Tennant, D. A test of a modified line intersect method of estimating root length. J. Ecol. 1975, 63, 995–1001. [Google Scholar] [CrossRef]
- Mitchell, K.J.; Glenday, A.C. The tiller population of pasture. N. Z. J. Agric. Res. 1958, 1, 305–318. [Google Scholar]
- Butler, B.M.; Matthew, C.; Heerdegen, R. The greenhouse effect—What consequences for seasonality of pasture production. Weather Clim. 1990, 10, 55–60. [Google Scholar]
- Hoglund, J.H.; Crush, J.R.; Brock, J.L.; Carran, R.A. Nitrogen fixation in pasture. 12. General discussion. N. Z. J. Exp. Agric. 1979, 1, 45–51. [Google Scholar]
- Moot, D.J.; Scott, W.R.; Roy, A.M.; Nicholls, A.C. Base temperature and thermal time requirements for germination and emergence of temperate pasture species. N. Z. J. Agric. Res. 2000, 43, 15–25. [Google Scholar] [CrossRef]
- Black, A.D.; Moot, D.J.; Lucas, R.J. Seedling development and growth of white clover, Caucasian clover, and perennial ryegrass sown in field and controlled environments. Proc. N. Z. Grassl. Assoc. 2002, 64, 197–204. [Google Scholar]
- Yang, J.Z.; Matthew, C.; Rowland, R.E. Tiller axis observations for perennial ryegrass (Lolium perenne) and tall rescue (Festuca arundinacea): Number of active phytomers, probability of tiller appearance, and frequency of root appearance per phytomer for three cutting heights. N. Z. J. Agric. Res. 1998, 41, 11–18. [Google Scholar] [CrossRef]
- Matthew, C.; van der Linden, A.; Hussain, S.; Easton, H.S.; Hatier, J.-H.B.; Horne, D.J. Which way forward in the quest for drought tolerance in perennial ryegrass. Proc. N. Z. Grassl. Assoc. 2012, 74, 195–200. [Google Scholar]
- Sackville Hamilton, C.A.G.; Cherrtt, J.M. The development of clover and ryegrass root systems in a pasture and their interactions with the soil fauna. In Plant Root Growth—An Ecological Perspective; Atkinson, D., Ed.; British Ecological Society Special Publication, Blackwell Scientific Publications: London, UK, 1991; pp. 291–301. [Google Scholar]
- Evans, P.S. The effect of repeated defoliation to three different levels on root growth of five pasture species. N. Z. J. Agric. Res. 1973, 16, 31–34. [Google Scholar] [CrossRef]
- Bland, W.L. Estimating root length density by the core break method. Soil Sci. Soc. Am. J. 1989, 53, 1595–1597. [Google Scholar] [CrossRef]
- De Willigen, P.; van Noordwijk, M. Roots, Plant Production and Nutrient Use Efficiency. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 1987. [Google Scholar]
- Gastal, F.; Saugier, B. Relationships between nitrogen uptake and carbon assimilation in whole plants of tall fescue. Plant Cell Environ. 1989, 12, 407–418. [Google Scholar] [CrossRef]
- Wilson, J.B. A review of evidence on the control of root:shoot ratio in relation to models. Ann. Bot. 1988, 61, 433–449. [Google Scholar]
- Davies, A. Structure of the grass sward. In Animal Production from Temperate Grassland; Gilsenan, B., Ed.; Irish Grassland and Animal Production Association: Dublin, Ireland, 1977; pp. 36–44. [Google Scholar]
- Troughton, A. Further studies on the relationship between root and shoot systems of grasses. J. Br. Grassl. Soc. 1960, 15, 4–47. [Google Scholar] [CrossRef]
- Robin, A.H.K.; Saha, P.S. Morphology of lateral roots of twelve rice cultivars of Bangladesh: Dimension increase and diameter reduction in progressive root branching at the vegetative stage. Plant Root 2015, 9, 34–42. [Google Scholar] [CrossRef]
- Troughton, A. Production of root axes and leaf elongation in perennial ryegrass in relation to dryness of the upper soil layer. J. Agric. Sci. 1980, 95, 533–538. [Google Scholar] [CrossRef]
- Stuckey, I.H. Seasonal growth of grass roots. Am. J. Bot. 1941, 28, 486–491. [Google Scholar] [CrossRef]
- Edwards, G.R.; Chapman, D.F. Plant responses to defoliation and relationships with pasture persistence. In Pasture Persistence Symposium; Mercer, C.F., Ed.; New Zealand Grassland Association: Dunedin, New Zeaalnd, 2011; pp. 99–108. [Google Scholar]
- Matthew, C.; Lemaire, G.; Sackville Hamilton, N.R.; Hernandez-Garay, A. A modified self-thinning equation to describe size/density relationships for defoliated swards. Ann. Bot. 1995, 76, 579–587. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Harvest Number and Median Date of Root Ingrowth Core Capture | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait | Grazing | Soil Depth | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | MEAN | S.E. 1 | S.E. 2 |
| (mm) | 25 December 1986 | 3 February 1987 | 13 March 1987 | 1 May 1987 | 10 June 1987 | 2 August 1987 | 26 September 1987 | 14 November 1987 | 29 December 1987 | |||||
| Root deposition (kg DM/ha/day) | L | 0–70 | 4.3 | 7.6 | 3.1 | 2.3 | 1.2 | 6.1 | 4.2 | 6.8 | 5.9 | 4.6 | 0.4 | 0.9 |
| 70–250 | 1.8 | 3.3 | 1.4 | 0.9 | - | 2 | 2 | 4.6 | 2.8 | 2.2 | 0.3 | 0.6 | ||
| 250–600 | 1.1 | 0.6 | 0.4 | 0.1 | - 3 | - | - | 2.4 | 1.4 | 1.1 | 0.3 | 0.4 | ||
| 0–600 | 7.2 | 11.5 | 4.9 | 3.3 | 1.8 | 8.1 | 6.2 | 13.8 | 10.1 | |||||
| H | 0–70 | 3.8 | 6 | 3.6 | 0.9 | 0.9 | 5.8 | 4.8 | 7.8 | 7.7 | 4.6 | 0.4 | 0.9 | |
| 70–250 | 2.1 | 3.4 | 1.8 | 0.4 | 0.4 | 2.6 | 2.8 | 3.5 | 3.1 | 2.2 | 0.3 | 0.6 | ||
| 250–600 | 1.9 | 0.8 | 0.6 | 0.1 | - 3 | - | - | 1.8 | 3 | 1.4 | 0.3 | 0.4 | ||
| 0–600 | 7.8 | 10.2 | 6 | 1.4 | 1.3 | 8.4 | 7.6 | 13.1 | 13.8 | |||||
| Mean | 0–70 | 4 | 6.8 | 3.3 | 1.6 | 1.1 | 6 | 4.5 | 7.3 | 6.8 | 4.6 | 0.3 | 0.6 | |
| 70–250 | 1.9 | 3.4 | 1.6 | 0.6 | 0.5 | 2.3 | 2.4 | 4 | 2.9 | 2.2 | 0.2 | 0.4 | ||
| 250–600 | 1.3 | 0.6 | 0.5 | 0.1 | - 3 | - | - | 2.2 | 2.2 | 1.2 | 0.2 | 0.3 | ||
| Mean diameter (mm) | Mean | 0.24 | 0.25 | 0.27 | 0.22 | 0.3 | 0.31 | 0.21 | 0.18 | 0.22 | 0.24 | 0.011 | 0.011 | |
| Relative root length 4 | Mean | 0.81 | 1.15 | 0.49 | 0.32 | 0.12 | 0.59 | 1.09 | 2.7 | 1.74 | 1 | n.d. 5 | n.d. | |
| Tiller population density (Tillers·m−2) | L | 4183 | 8257 | 5615 | 4624 | 3853 | 5284 | 4624 | 4404 | 6606 | 5270 | 622 | 289 | |
| H | 5725 | 11,009 | 7927 | 7266 | 6606 | 6606 | 6606 | 7486 | 11,119 | 7820 | ||||
| Grazing | Harvest Number and Median Date of Root Ingrowth Core Capture | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | MEAN | ||
| 25 December 1986 | 3 February 1987 | 13 March 1987 | 1 May 1987 | 10 June 1987 | 2 August 1987 | 26 September 1987 | 14 November 1987 | 29 December 1987 | |||
| P 1 (days) | 6.4 | 6.6 | 7.4 | 9.4 | 13.1 | 13.2 | 10.5 | 7.6 | 6.6 | 8.8a | |
| AS | 1.03 | 0.97 | 0.89 | 0.79 | 0.72 | 0.99 | 1.26 | 1.38 | 1.15 | ||
| Wr | L | 1.1 | 0.9 | 0.6 | 0.6 | 0.6 | 1.9 | 1.4 | 2.3 | 1.0 | 1.2 |
| H | 0.8 | 0.6 | 0.5 | 0.2 | 0.2 | 1.6 | 1.2 | 1.3 | 0.8 | 0.8 | |
| Fr(0–70) | Fr(70–250) | Fr(250–600) | Fr(0–600) | Diameter | |
|---|---|---|---|---|---|
| P | −0.409 | −0.475 | −0.690 | −0.521 | 0.587 |
| (0.275) | (0.196) | (0.040) | (0.150) | (0.097) | |
| 1/P | 0.443 | 0.485 | 0.708 | 0.547 | −0.476 |
| (0.232) | (0.186) | (0.033) | (0.127) | (0.195) | |
| AS | 0.758 | 0.830 | 0.620 | 0.804 | −0.700 |
| (0.018) | (0.006) | (0.075) | (0.009) | (0.036) | |
| 1/AS | −0.816 | −0.856 | −0.609 | −0.842 | 0.642 |
| (0.007) | (0.003) | (0.002) | (0.004) | (0.062) | |
| Cos day of year 1 | 0.660 | 0.720 | 0.807 | 0.760 | −0.614 |
| (0.053) | (0.029) | (0.009) | (0.018) | (0.078) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matthew, C.; Mackay, A.D.; Robin, A.H.K. Do Phytomer Turnover Models of Plant Morphology Describe Perennial Ryegrass Root Data from Field Swards? Agriculture 2016, 6, 28. https://doi.org/10.3390/agriculture6030028
Matthew C, Mackay AD, Robin AHK. Do Phytomer Turnover Models of Plant Morphology Describe Perennial Ryegrass Root Data from Field Swards? Agriculture. 2016; 6(3):28. https://doi.org/10.3390/agriculture6030028
Chicago/Turabian StyleMatthew, Cory, Alec D. Mackay, and Arif Hasan Khan Robin. 2016. "Do Phytomer Turnover Models of Plant Morphology Describe Perennial Ryegrass Root Data from Field Swards?" Agriculture 6, no. 3: 28. https://doi.org/10.3390/agriculture6030028