
Potential Geographical Distribution of Lagerstroemia excelsa under Climate Change
Abstract
:1. Introduction
2. Materials and Methods
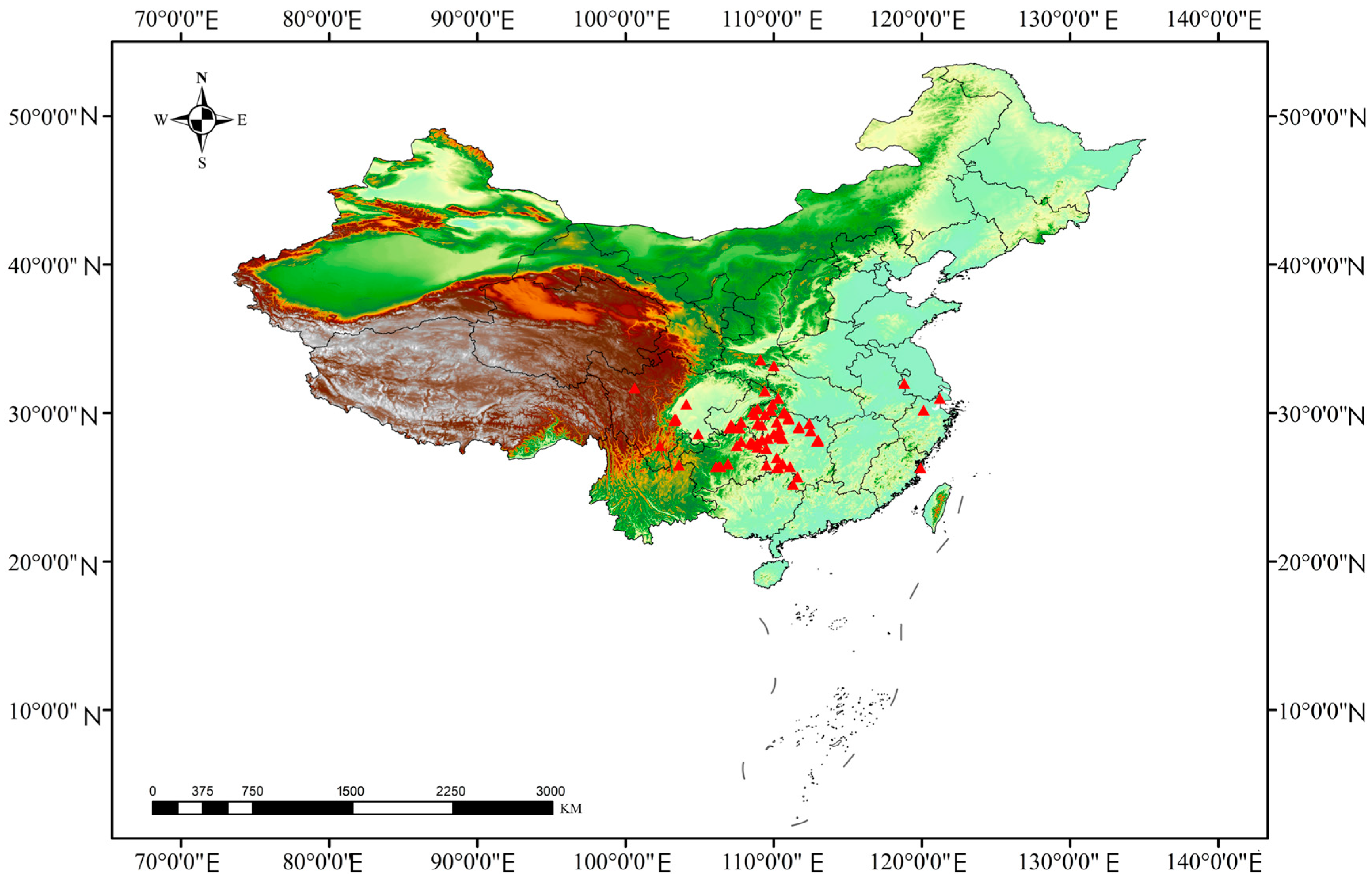
2.1. Species Occurrence Data Collection
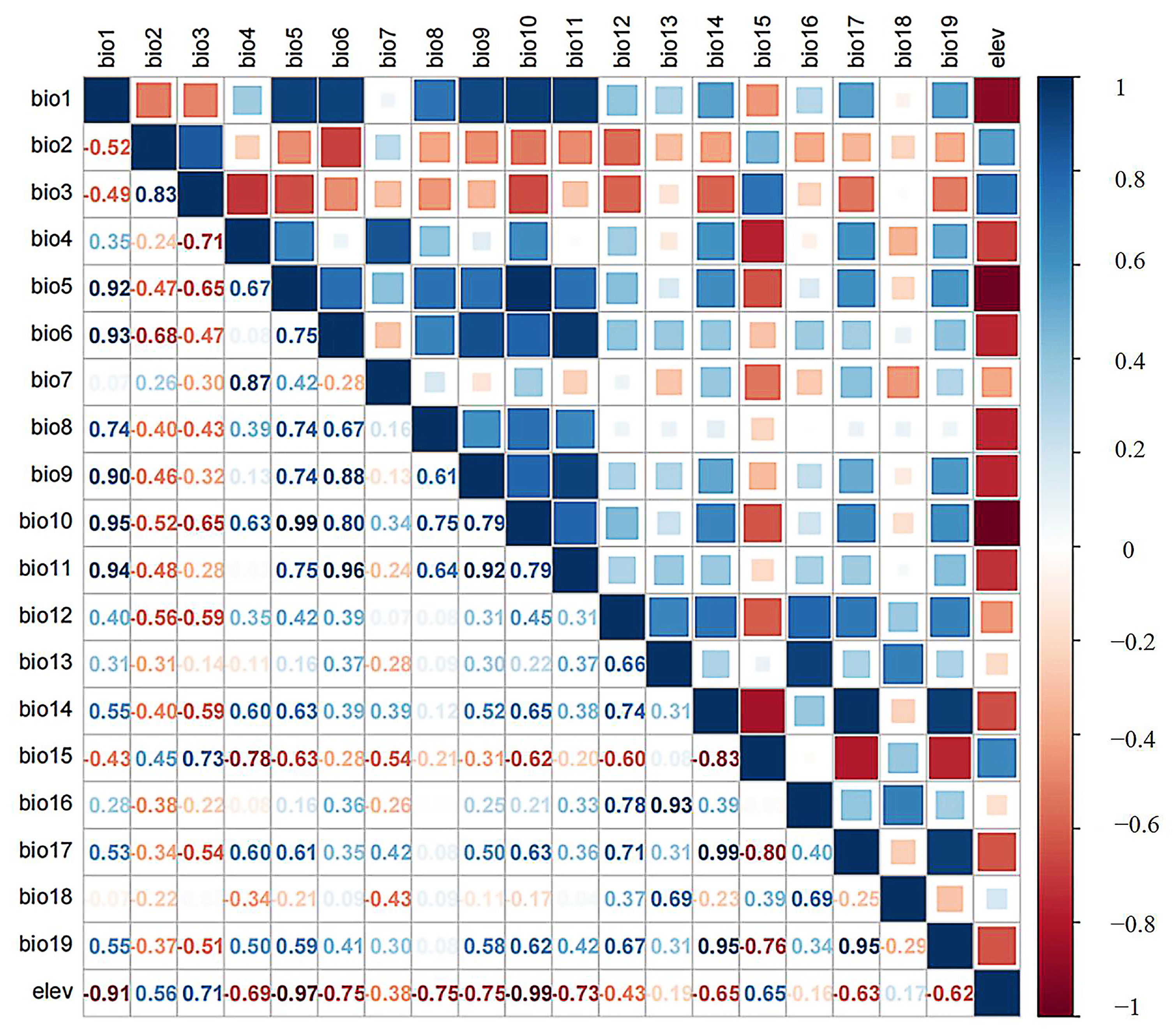
2.2. Environmental Attributes Collection and Selection
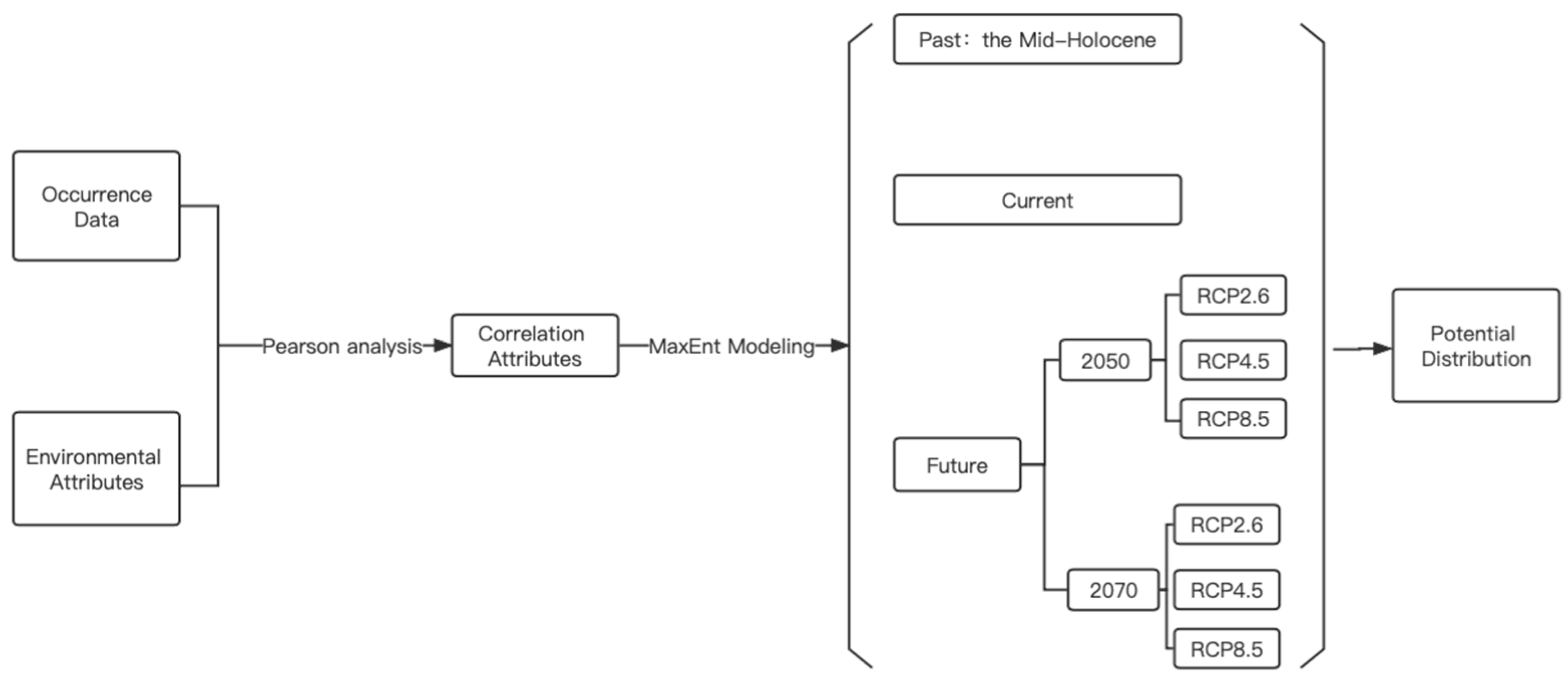
2.3. Potential Geographical Distribution Prediction
2.4. Research Framework
3. Results
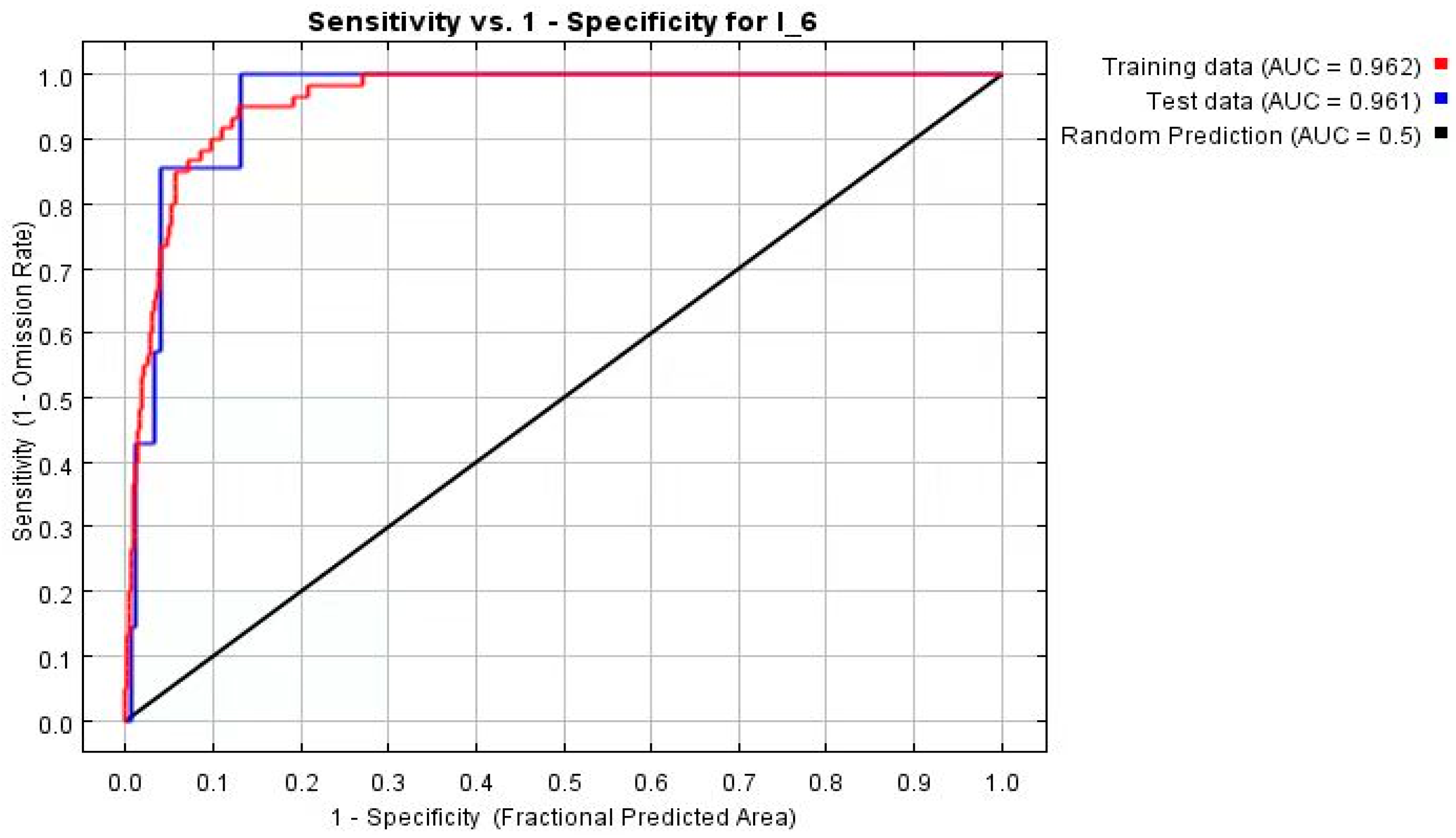
3.1. Model Evaluation
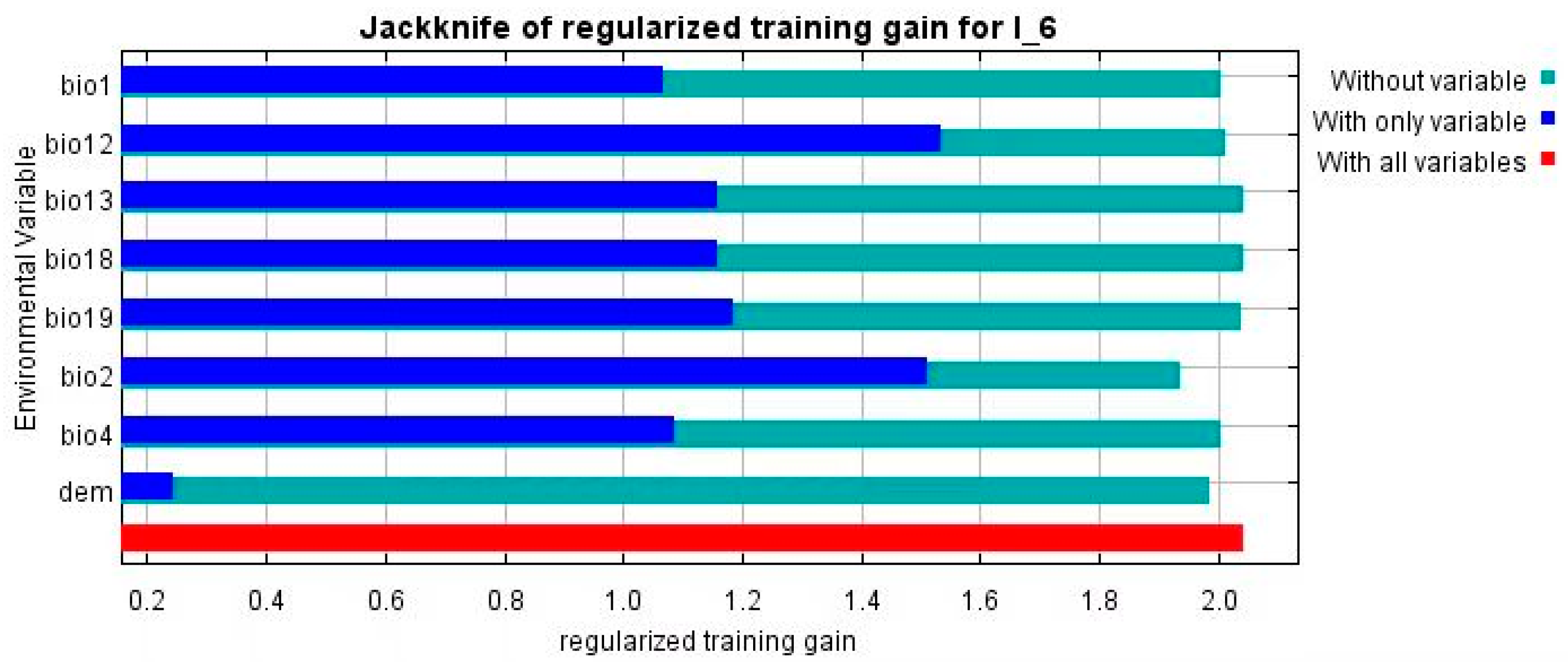
3.2. Attribute Contribution
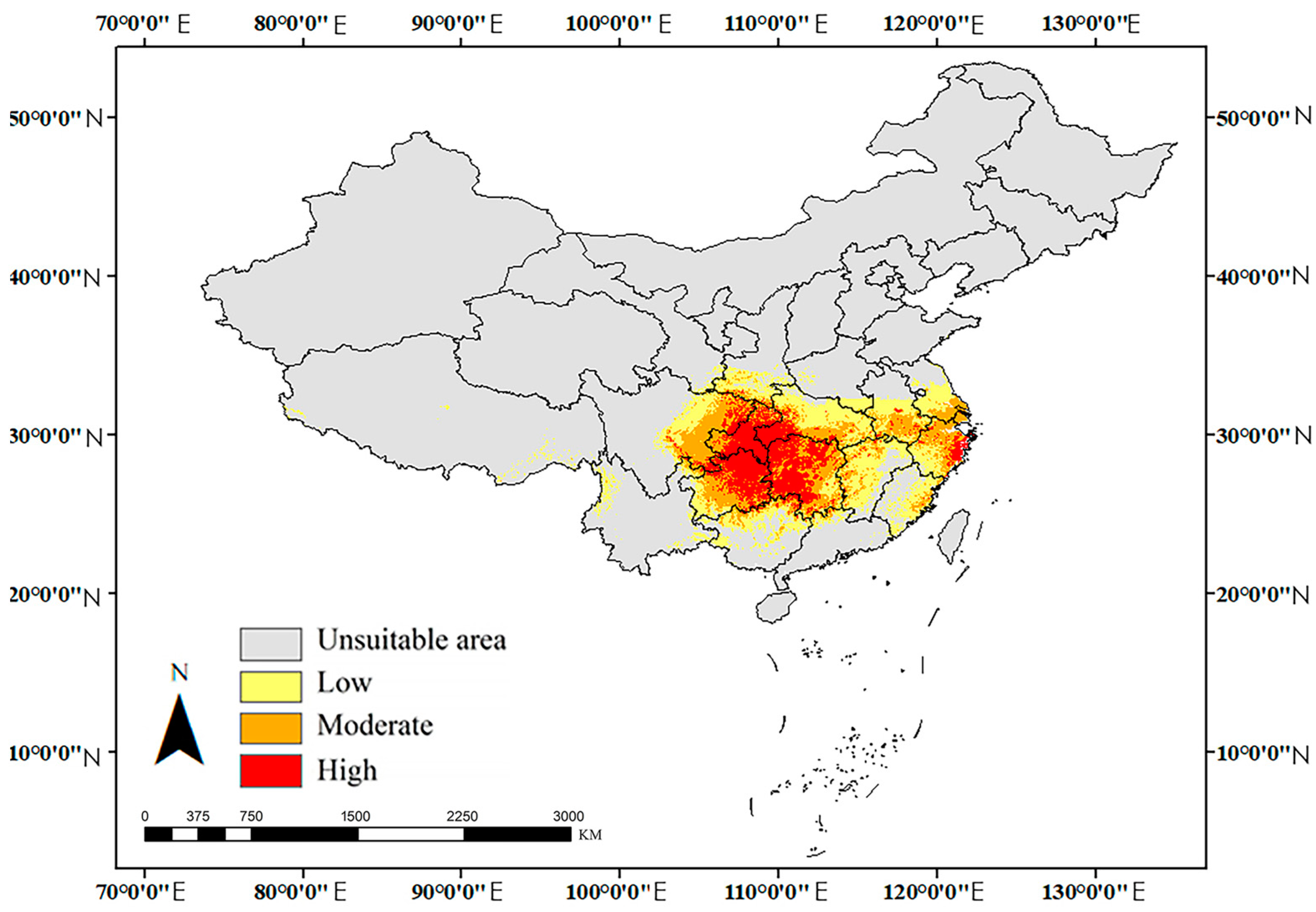
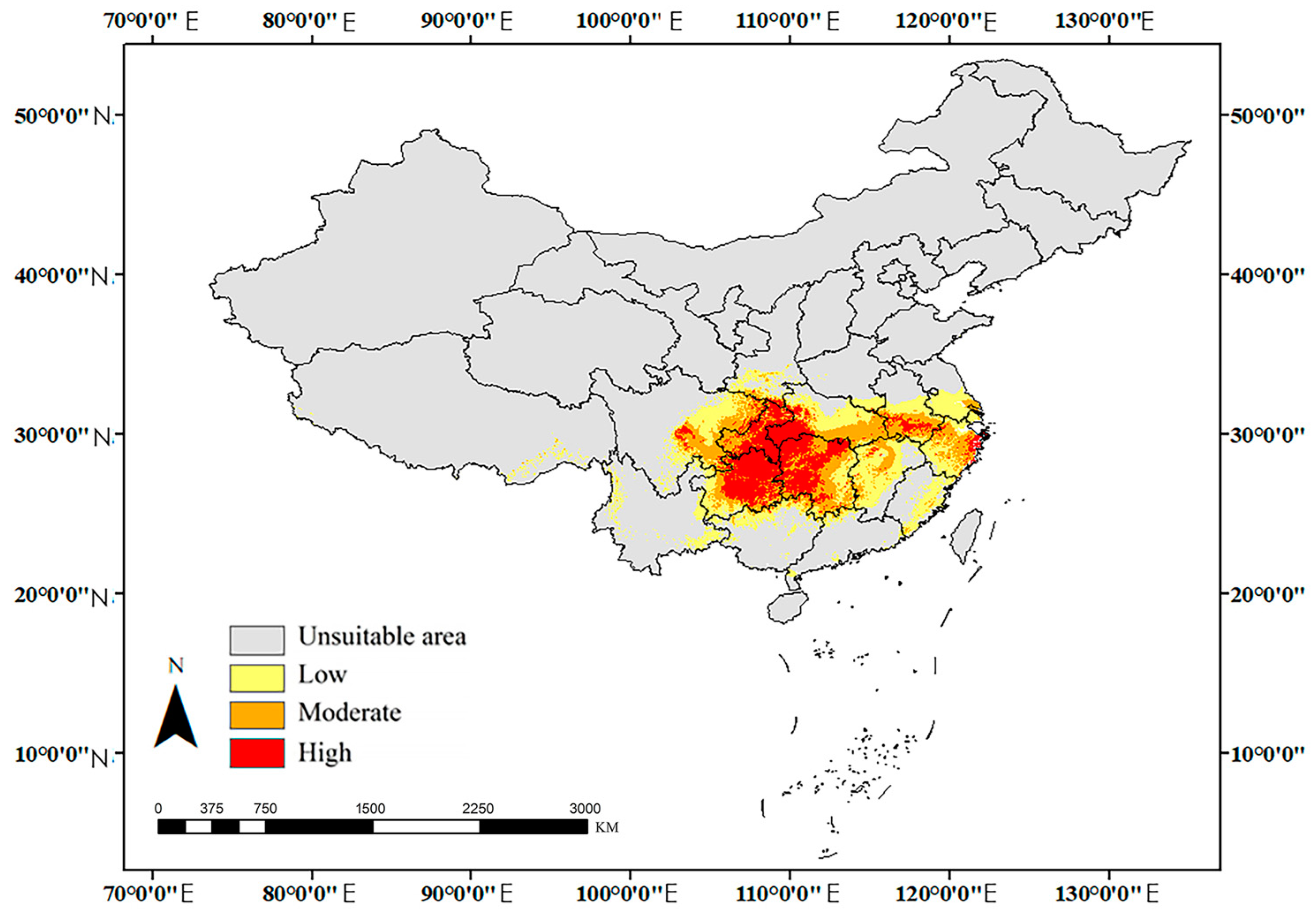
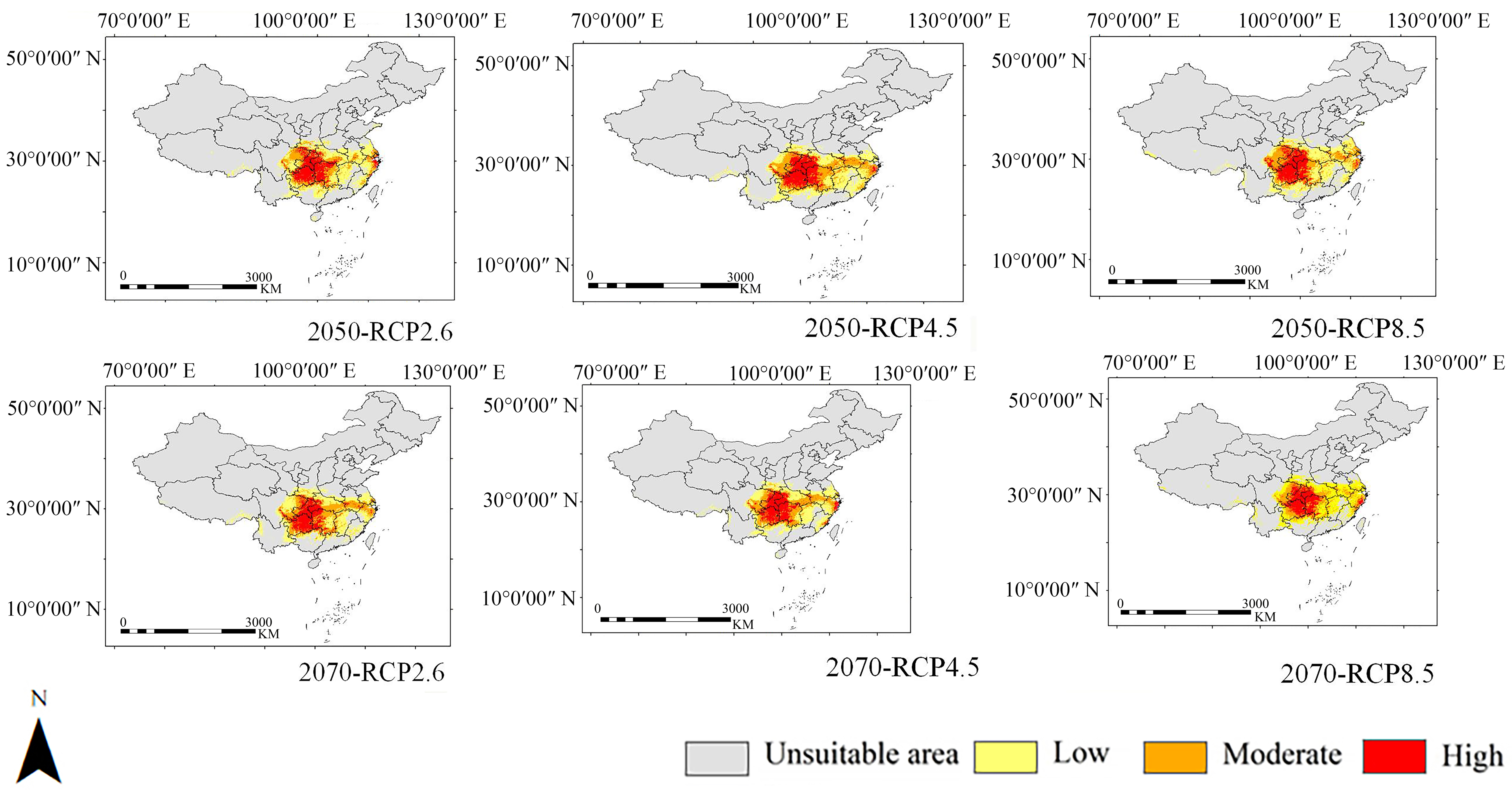
3.3. Potential Distributions
4. Discussion
4.1. Environmental Attributes Influencing the Distribution of Lagerstroemia excelsa
4.2. Response of Lagerstroemia excelsa Distribution to Climate Change
4.3. Conservation and Utilization of Lagerstroemia excelsa
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bhadra, P.; Maitra, S.; Shankar, T.; Hossain, A.; Praharaj, S.; Aftab, T. Climate change impact on plants: Plant responses and adaptations. In Plant Perspectives to Global Climate Changes; Academic Press: Cambridge, MA, USA, 2022; pp. 1–24. [Google Scholar] [CrossRef]
- Alizadeh, B.; Hitchmough, J. How will climate change affect future urban naturalistic herbaceous planting? The role of plant origin and fitness. Urban For. Urban Green. 2020, 54, 126786. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusià, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; et al. Impacts of global change on Mediterranean forests and their services. Forests 2017, 8, 463. [Google Scholar] [CrossRef]
- Allen, J.M.; Folk, R.A.; Soltis, P.S.; Soltis, D.E.; Guralnick, R.P. Biodiversity synthesis across the green branches of the tree of life. Nat. Plants 2019, 5, 11–13. [Google Scholar] [CrossRef]
- Fischer, F.M.; Wrigh, A.J.; Eisenhauer, N.; Ebeling, A.; Roscher, C.; Wagg, C.; Weigelt, A.; Weisser, W.W.; Pillar, V.D. Plant species richness and functional traits affect community stability after a flood event. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150276. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Farzana, R.; Sihalath, S.; Rattanavong, S.; Vongsouvath, M.; Sands, K.; Newton, P.N.; Dance, D.A.; Hassan, B.; Walsh, T. A one-health sampling strategy to explore the dissemination and relationship between colistin resistance in human, animal, and environmental sectors in Laos. Engineering 2022, 15, 45–56. [Google Scholar] [CrossRef]
- Ngarega, B.K.; Chaibva, P.; Masocha, V.F.; Saina, J.K.; Khine, P.K.; Schneider, H. Application of MaxEnt modeling to evaluate the climate change effects on the geographic distribution of Lippia javanica (Burm. f.) Spreng in Africa. Environ. Monit. Assess. 2024, 196, 62. [Google Scholar] [CrossRef]
- Egolf, D.R. Four new Lagerstroemia indica L. cultivars, ‘Catawba’, ‘Conestoga’, ‘Potomac’ and ‘Powhatan’. Baileya 1967, 15, 7–13. [Google Scholar]
- Wang, Y.S.; Ni, F.; Yin, D.H.; Chen, L.; Li, Y.H.; He, L.X.; Zhang, Y.L. Physiological response of Lagerstroemia indica (L.) Pers. seedlings to drought and rewatering. Trop. Plant Biol. 2021, 14, 360–370. [Google Scholar] [CrossRef]
- Wang, J. Shi Yi Ji; Zhonghua Book Company: Beijing, China, 1981. [Google Scholar]
- Zheng, G.; Wei, L.L.; Ma, L.; Wu, Z.Q.; Gu, C.H.; Chen, K. Comparative analyses of chloroplast genomes from 13 Lagerstroemia (Lythraceae) species: Identification of highly divergent regions and inference of phylogenetic relationships. Plant Mol. Biol. 2020, 102, 659–676. [Google Scholar] [CrossRef]
- Xiang, G.F.; Yan, L.H.; Qian, J.H.; Jiang, L.Y.; Tian, X.M.; Xiang, Y.W. Effects of the different site conditions on growth of Lagerstroemia excelsa. Hunan For. Sci. Technol. 2017, 44, 46–49. (In Chinese) [Google Scholar] [CrossRef]
- Zhou, J.W.; Yan, L.H.; Jiang, L.Y.; Rao, L.Y.; She, W.S. The application of Lagerstroemia excelsa in vegetation restoration of rocky desertification difficult afforestation land. Hunan For. Sci. Technol. 2018, 45, 56–59. (In Chinese) [Google Scholar] [CrossRef]
- Wu, C.H. Effect of softwood cutting propagation of Lagerstroemia excelsa. Hunan For. Sci. Technol. 2016, 97–100, 119. (In Chinese) [Google Scholar]
- Liu, J.; Wang, L.; Sun, C.; Xi, B.; Li, D.; Chen, Z.; He, Q.; Weng, X.; Jia, L. Global distribution of soapberries (Sapindus L.) habitats under current and future climate scenarios. Sci. Rep. 2021, 11, 19740. [Google Scholar] [CrossRef]
- Damaneh, J.M.; Ahmadi, J.; Rahmanian, S.; Sadeghi, S.M.M.; Nasiri, V.; Borz, S.A. Prediction of wild pistachio ecological niche using machine learning models. Ecol. Inform. 2022, 72, 101907. [Google Scholar] [CrossRef]
- Busby, J.R. Bioclim, a bioclimatic analysis and prediction system. In Nature Conservation: Cost Effective Biological Surveys and Data Analysis; Margules, C.R., Austin, M.P., Eds.; CSIRO: Canberra, Australia, 1991; pp. 64–68. [Google Scholar]
- Lanoiselet, V.; Cother, E.J.; Ash, G.J. CLIMEX and DYMEX simulations of the potential occurrence of rice blast disease in south-eastern Australia. Australas. Plant Pathol. 2002, 31, 1–7. [Google Scholar] [CrossRef]
- Stockwell, D.R.B. Genetic algorithms II—Species distribution modelling. In Machine Learning Methods for Ecological Applications; Fielding, A.H., Ed.; Springer Publishing: New York, NY, USA, 1999; pp. 123–144. [Google Scholar]
- Feng, L.; Tian, X.N.; EL-Kassaby, Y.A.; Qiu, J.; Feng, Z.; Sun, J.J.; Wang, G.B.; Wang, T.L. Predicting suitable habitats of Melia azedarach L. in China using data mining. Sci. Rep. 2022, 12, 12617. [Google Scholar] [CrossRef]
- Zhang, W.Q.; Zhang, D.L.; Zhao, Y.H.; Hao, S.W.; Jin, X.L. Impact of climate changes on natural and cultivated distribution of US magnolias. HortScience 2022, 57, S38. [Google Scholar]
- Zhao, Y.H.; Zhang, D.L.; Hao, S.W.; Zhang, W.Q.; Wen, Y.F. Shift of Phellodendron natural distribution under climate changes. HortScience 2022, 57, S59. [Google Scholar]
- Kumar, S.; Kler, T.K.; Singh, G.; Sekhon, G.; Sahni, T. Partial altitudinal movement of Himalayan bulbul Pycnonotus leucogenys in Himalayan siwalik range. Pharma Innov. J. 2022, 11, 1732–1738. [Google Scholar] [CrossRef]
- Almeida, A.M.; Martins, M.J.; Campagnolo, M.L.; Fernandez, P.; Albuquerque, T.; Gerassis, S.; Goncalves, J.C.; Ribeiro, M.M. Presiction scenarios of past, present, and future environmental suitability for the Mediterranean species Arbutus unedo L. Sci. Rep. 2022, 12, 84. [Google Scholar] [CrossRef] [PubMed]
- Sillero, N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Model. 2011, 222, 1343–1346. [Google Scholar] [CrossRef]
- Ye, P.; Zhang, G.; Zhao, X.; Chen, H.; Si, Q.; Wu, J. Potential geographical distribution and environmental explanations of rare and endangered plant species through combined modeling: A case study of Northwest Yunnan, China. Ecol. Evol. 2021, 11, 13052–13067. [Google Scholar] [CrossRef]
- Zhao, Y.; Wen, Y.; Zhang, W.; Wang, C.; Yan, Y.; Hao, S.; Zhang, D. Distribution pattern and change prediction of Phellodendron habitat in China under climate change. Ecol. Evol. 2023, 13, e10374. [Google Scholar] [CrossRef] [PubMed]
- Kass, J.M.; Muscarella, R.; Galante, P.J.; Bohl, C.L.; Pinilla-Buitrago, G.E.; Boria, R.A.; Soley-Guardia, M.; Anderson, R.P. ENMeval 2.0: Redesigned for customizable and reproducible modeling of species’ niches and distributions. Methods Ecol. Evol. 2021, 12, 1602–1608. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, J. Predicting the Potential Global Geographical Distribution of Two Icerya Species under Climate Change. Forests 2020, 11, 684. [Google Scholar] [CrossRef]
- Alizadeh, B.; Hitchmough, J. A review of urban landscape adaptation to the challenge of climate change. Int. J. Clim. Chang. Strateg. Manag. 2019, 11, 178–194. [Google Scholar] [CrossRef]
- Tiwari, S.; Patel, A.; Singh, M.; Prasad, S.M. Regulation of temperature stress in plants. In Plant Life Under Changing Environment; Academic Press: Cambridge, MA, USA, 2020; pp. 25–45. [Google Scholar] [CrossRef]
- Thakur, D.; Chawla, A. Functional diversity along elevational gradients in the high-altitude vegetation of the western Himalaya. Biodivers. Conserv. 2019, 28, 1977–1996. [Google Scholar] [CrossRef]
- Lambers, H.; Oliveira, R.S. Plant Water Relations. In Plant Physiological Ecology; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Mattera, M.G.; Pastorino, M.J.; Lantschner, M.V.; Marchelli, P.; Soliani, C. Genetic diversity and population structure in Nothofagus pumilio, a foundation species of Patagonian forests: Defining priority conservation areas and management. Sci. Rep. 2020, 10, 19231. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attributes | Summary |
|---|---|
| bio1 | Annual mean temperature |
| bio2 | Mean diurnal range |
| bio3 | Isothermality |
| bio4 | Temperature seasonality |
| bio5 | Max temperature of warmest month |
| bio6 | Min temperature of coldest month |
| bio7 | Temperature annual range |
| bio8 | Mean temperature of wettest quarter |
| bio9 | Mean temperature of driest quarter |
| bio10 | Mean temperature of warmest quarter |
| bio11 | Mean temperature of coldest quarter |
| bio12 | Annual precipitation |
| bio13 | Precipitation of wettest month |
| bio14 | Precipitation of driest month |
| bio15 | Precipitation seasonality coefficient of variation |
| bio16 | Precipitation of wettest quarter |
| bio17 | Precipitation of driest quarter |
| bio18 | Precipitation of warmest quarter |
| bio19 | Precipitation of coldest quarter |
| Ele | Elevation |
| Attributes | Summary | Percent Contribution | Permutation Importance |
|---|---|---|---|
| bio1 | Annual mean temperature | 4.9 | 30.1 |
| bio2 | Mean diurnal range | 13.5 | 6.7 |
| bio4 | Temperature seasonality | 8.6 | 3.9 |
| bio12 | Annual precipitation | 38.3 | 47 |
| bio13 | Precipitation of wettest month | 0.4 | 0.4 |
| bio18 | Precipitation of warmest quarter | 0.1 | 1.5 |
| bio19 | Precipitation of coldest quarter | 28.7 | 0.1 |
| Ele | Elevation | 4.9 | 30.1 |
| High-Suitability Area (km²) | Proportion of High-Suitable Area (%) | Medium-Suitability Area (km²) | Low-Suitability Area (km²) | Unsuitable Area (km²) | Suitability Area (km²) | ||
|---|---|---|---|---|---|---|---|
| Mid-Holocene (MH) | 3.23 × 105 | 24.84 | 3.79 × 105 | 5.99 × 105 | 8.29 × 106 | 1.30 × 106 | |
| Current | 3.35 × 105 | 23.83 | 4.34 × 105 | 6.36 × 105 | 8.19 × 106 | 1.41 × 106 | |
| 2050 | RCP2.6 | 3.47 × 105 | 23.55 | 3.74 × 105 | 7.54 × 105 | 8.12 × 106 | 1.48 × 106 |
| RCP4.5 | 3.43 × 105 | 22.89 | 4.54 × 105 | 7.01 × 105 | 8.09 × 106 | 1.50 × 106 | |
| RCP8.5 | 3.47 × 105 | 23.12 | 4.29 × 105 | 7.24 × 105 | 8.09 × 106 | 1.50 × 106 | |
| 2070 | RCP2.6 | 3.29 × 105 | 22.34 | 4.44 × 105 | 6.99 × 105 | 8.12 × 106 | 1.47 × 106 |
| RCP4.5 | 3.48 × 105 | 23.88 | 4.31 × 105 | 6.78 × 105 | 8.14 × 106 | 1.46 × 106 | |
| RCP8.5 | 3.26 × 105 | 23.99 | 3.46 × 105 | 6.86 × 105 | 8.23 × 106 | 1.36 × 106 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, S.; Zhang, D.; Wen, Y. Potential Geographical Distribution of Lagerstroemia excelsa under Climate Change. Agriculture 2024, 14, 191. https://doi.org/10.3390/agriculture14020191
Hao S, Zhang D, Wen Y. Potential Geographical Distribution of Lagerstroemia excelsa under Climate Change. Agriculture. 2024; 14(2):191. https://doi.org/10.3390/agriculture14020191
Chicago/Turabian StyleHao, Siwen, Donglin Zhang, and Yafeng Wen. 2024. "Potential Geographical Distribution of Lagerstroemia excelsa under Climate Change" Agriculture 14, no. 2: 191. https://doi.org/10.3390/agriculture14020191