

Serendipita indica: A Biostimulant Enhancing Low-Temperature Tolerance and Active Constituent Levels in Polygonum cuspidatum
{kind=link}
{kind=link}
{kind=link}
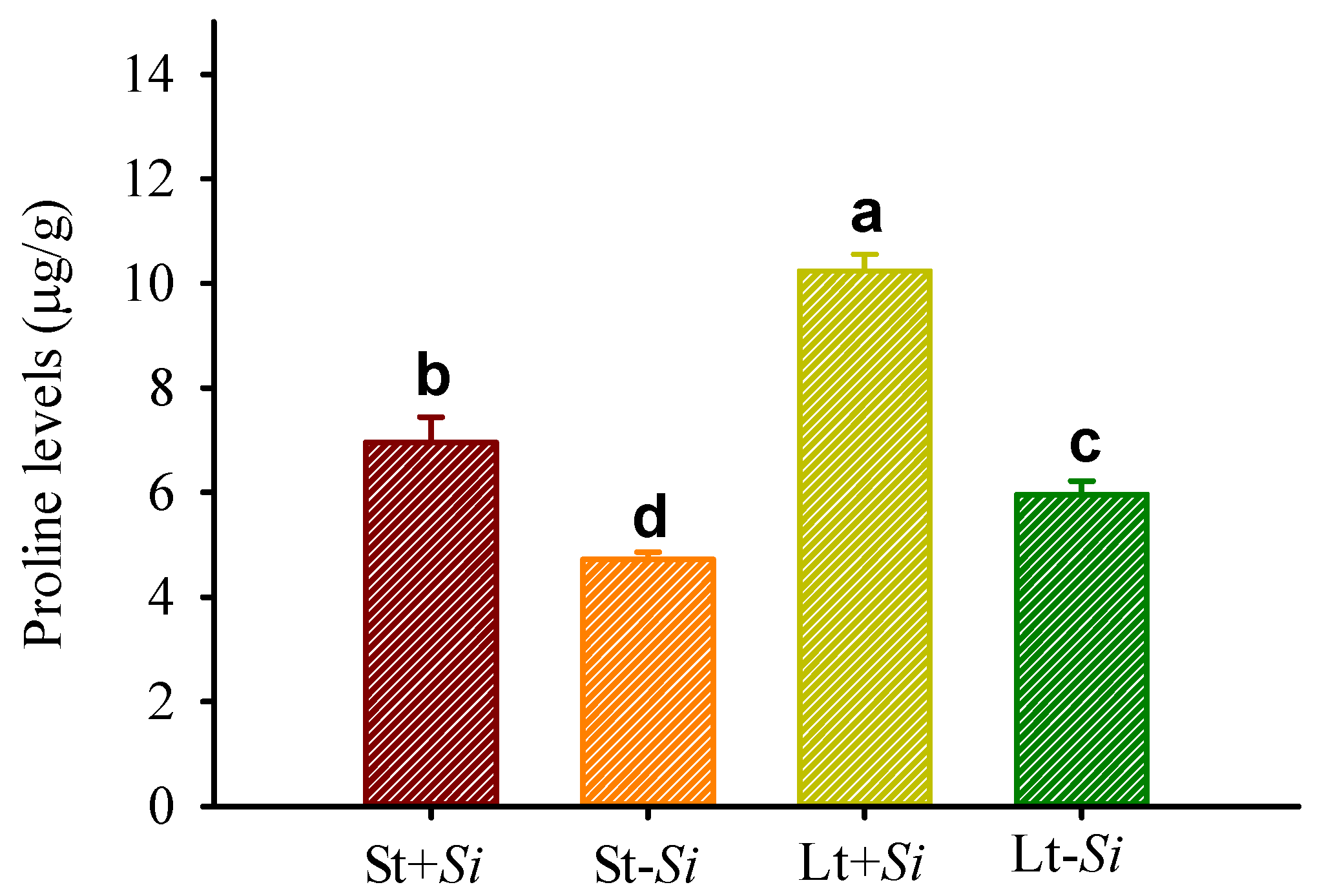{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of the Fungal Inoculum
2.2. Plant Culture, Fungal Inoculation, and Low-Temperature Treatment
2.3. Determination of Plant Biomass and Leaf Gas Exchange
2.4. Determination of Proline Levels in Leaves
2.5. Determination of the Activity of Three Antioxidant Enzymes in Leaves
2.6. Determination of the Levels of Four Medicinal Substances in Roots
2.7. Determinations of PcRS and PcRS11 Gene Expression in Roots
2.8. Data Analysis
3. Results
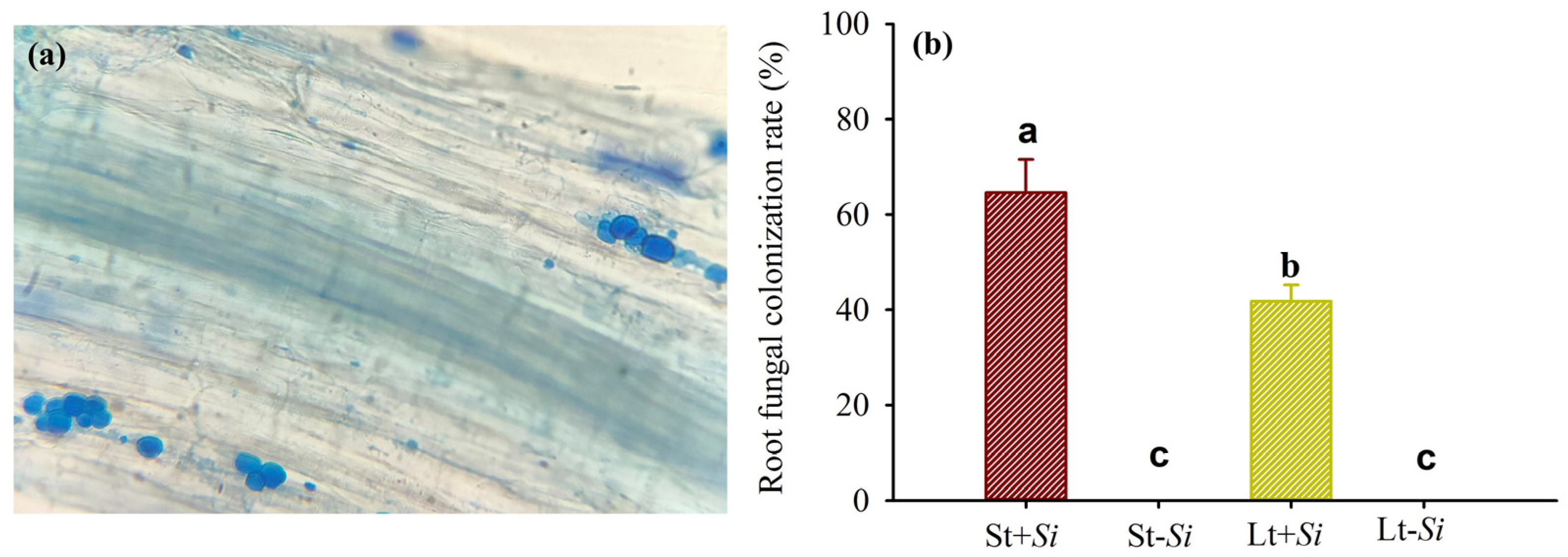
3.1. Effect of Low Temperatures on the Root Fungal Colonization Rate
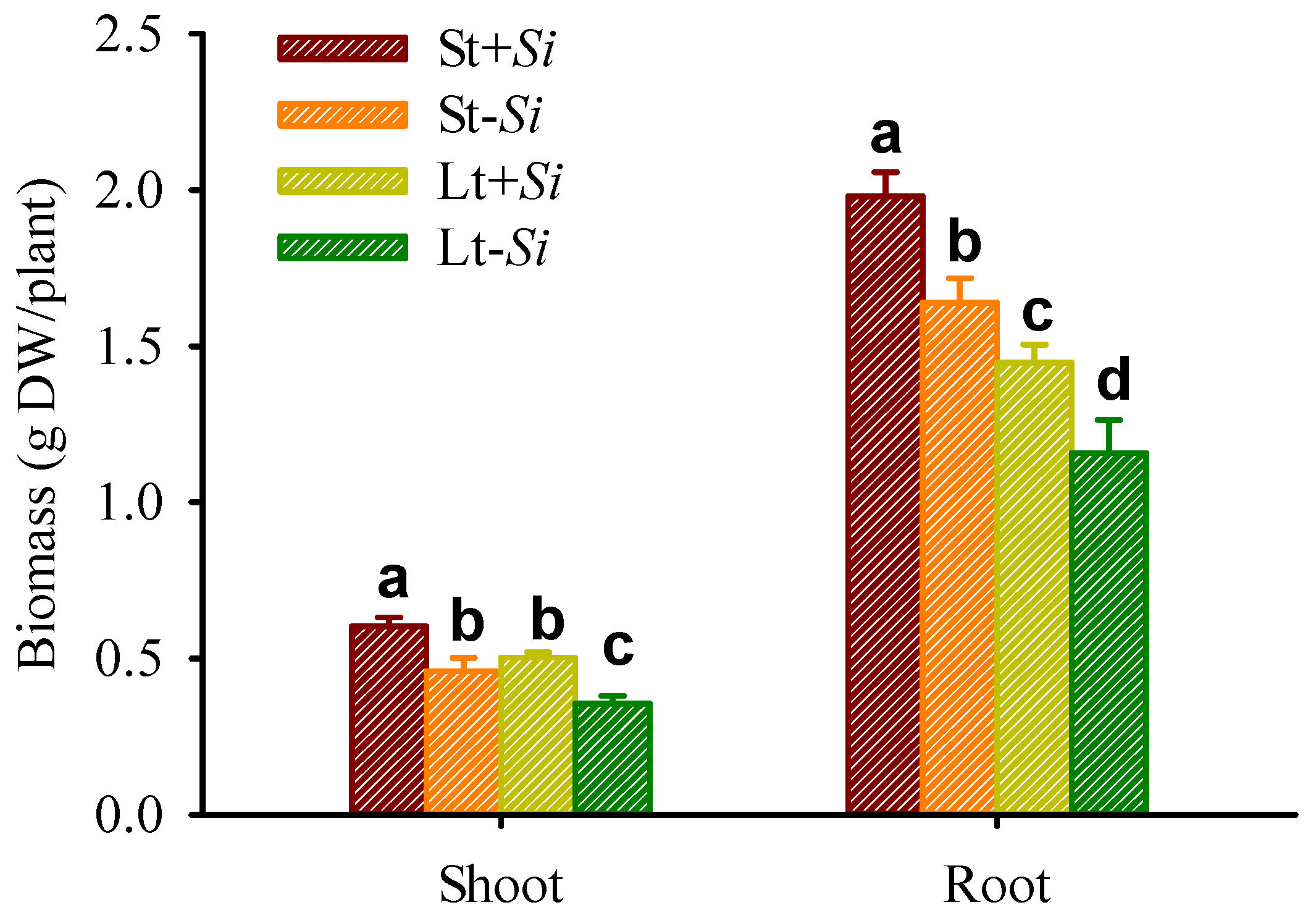
3.2. Effect of Low Temperatures and S. indica on Biomass Production
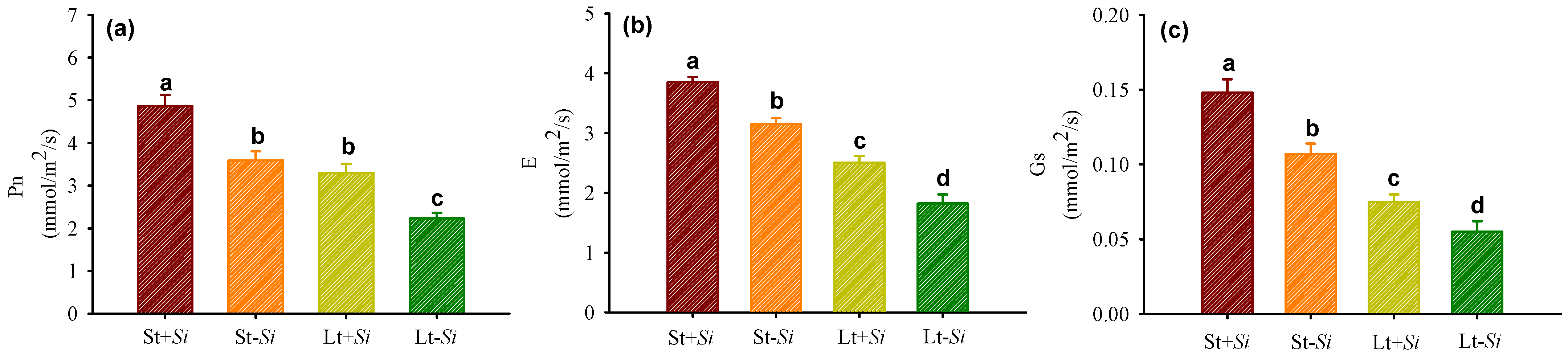
3.3. Effect of Low Temperatures and S. indica on Leaf Gas Exchange
3.4. Effect of Low Temperatures and S. indica on Proline Levels in Roots
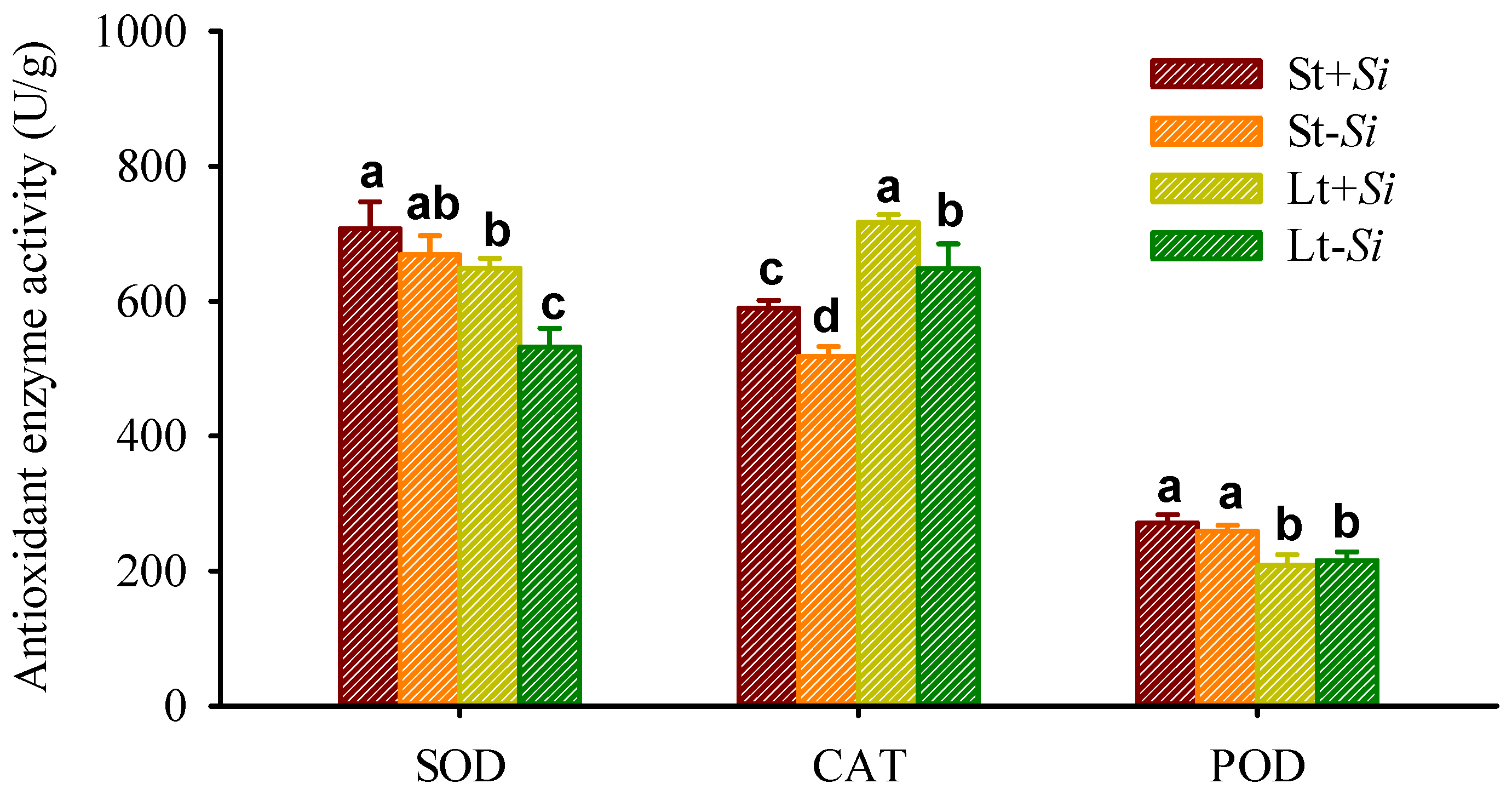
3.5. Effect of Low Temperatures and S. indica on Antioxidant Enzyme Activity in Roots
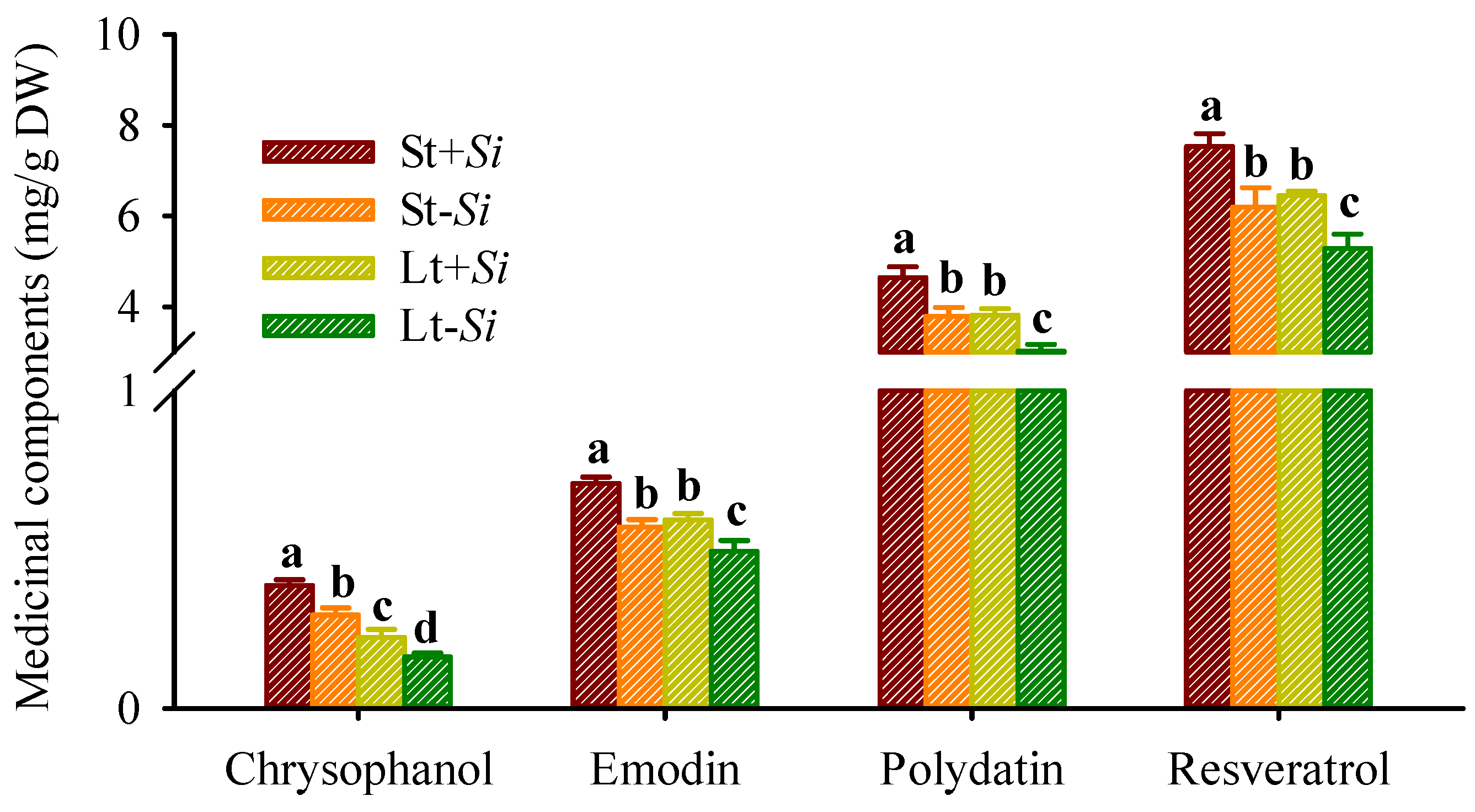
3.6. Changes in Medicinal Component Levels in Roots
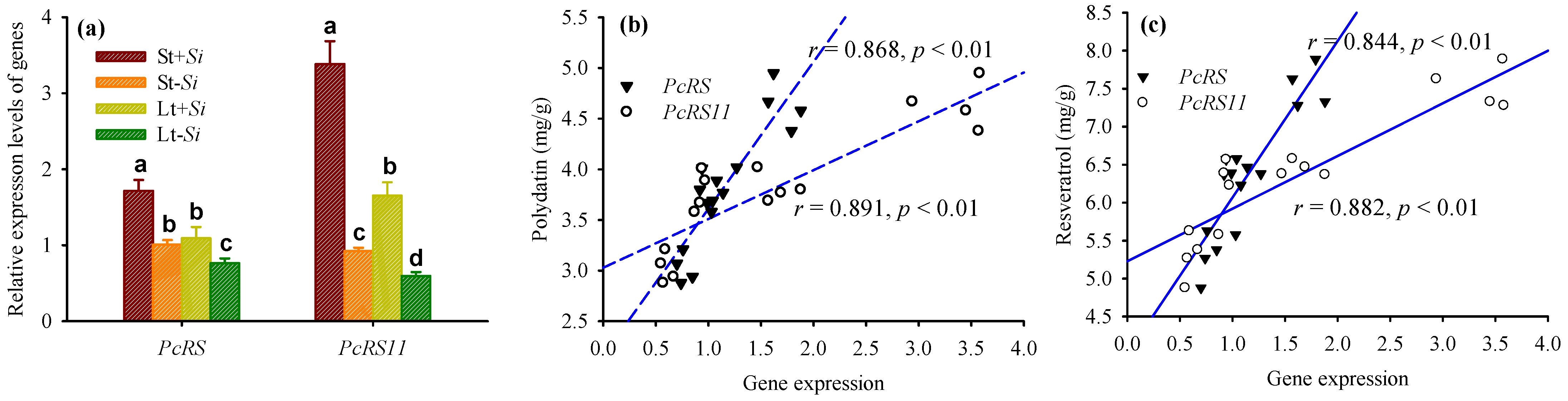
3.7. Changes in PcRS Gene Expression in Roots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jiang, Q.C.; Du, J.; Wang, J.C.; Yu, Z.H.; Wang, Y.; Liu, Z.Y. Expression and function analysis of transcription factor PcMYB2 from Polygonum cuspidatum. Biotechnol. Bull. 2023, 39, 217–223. [Google Scholar]
- Sun, R.T.; Feng, X.C.; Zhang, Z.Z.; Zhou, N.; Feng, H.D.; Liu, Y.M.; Hashem, A.; Al-Arjani, A.B.F.; Abd_Allah, E.F.; Wu, Q.S. Root endophytic fungi regulate changes in sugar and medicinal compositions of Polygonum cuspidatum. Front. Plant Sci. 2022, 13, 818909. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.H.; Zha, J.P.; Zhan, W.H.; Zhang, D.Q. Contents comparison of resveratrol and polydatin in the wild Polygonum cuspidatum plant and its tissue cultures. China J. Chin. Mat. Med. 2006, 31, 637–641. [Google Scholar]
- Li, Y.H.; Cheng, C.C.; Yang, S.; Liu, C.B.; Yin, Y.F.; Xie, D.M.; Wang, H.B. Study on ecological suitability zoning of Polygonum cuspidatum Sieb. et Zucc. Chin. J. Informat. TCM 2022, 29, 1–4. [Google Scholar]
- Zhang, J.; Yu, J.; Jing, X.L.; Wu, C.J.; Zheng, J.; Gao, Y. Research on reproduction technology of Polygonum cuspidatum seeds. J. Anhui Agric. Sci. 2017, 45, 120–122. [Google Scholar]
- Zhao, W.H.; Bi, J.W.; Zhao, Y.J.; Qiu, S.Y.; Madalin, P.; Oliver, B.; Christina, R.; Ju, R.T.; Wu, J.H.; Liao, Z.Y.; et al. Latitudinal variation and influencing factors of two secondary metabolites in Reynoutria japonica leaves. J. Fudan Univ. (Nat. Sci.) 2022, 61, 249–259. [Google Scholar]
- Verma, S.; Varma, A.; Rexer, K.-H.; Hassel, A.; Kost, G.; Sarbhoy, A.; Bisen, P.; Bütehorn, B.; Franken, P. Piriformospora indica, gen. et sp. nov., a new root-colonizing fungus. Mycologia 1998, 90, 896–903. [Google Scholar] [CrossRef]
- Zou, Y.N.; Xu, Y.J.; Liu, R.C.; Huang, G.M.; Kuča, K.; Srivastava, A.K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Two different strategies of Diversispora spurca-inoculated walnut seedlings to improve leaf P acquisition at low and moderate P levels. Front. Plant Sci. 2023, 14, 1140467. [Google Scholar] [CrossRef]
- Yang, L.; Zou, Y.N.; Tian, Z.H.; Wu, Q.S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Boorboori, M.R.; Zhang, H.-Y. The role of Serendipita indica (Piriformospora indica) in improving plant resistance to drought and salinity stresses. Biology 2022, 11, 952. [Google Scholar] [CrossRef]
- Li, L.; Feng, Y.; Qi, F.; Hao, R. Research progress of Piriformospora indica in improving plant growth and stress resistance to plant. J. Fungi 2023, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.M.; Zou, Y.N.; Shu, B.; Wu, Q.S. Arbuscular mycorrhizal fungi and endophytic fungi differentially modulate polyamines or proline of peach in response to soil flooding. Pedosphere 2023. [Google Scholar] [CrossRef]
- Wang, H.L. The Effects of Piriformospora indica on the Growth, Quality and Disease Resistance of Fruits and Vegetables and Related Mechanisms Involved. Master’s Dissertation, Zhejiang University, Hangzhou, China, 2015. [Google Scholar]
- Liang, D.D.; Huang, X.P.; Tang, Q.W.; Wei, X.X.; Li, B.Q.; Oelmüller, R.; Chen, F.X. Effect of Piriformospora indica on the cold resistance of passion fruit. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2022, 51, 748–753. [Google Scholar]
- Alizadeh, M.; Pirdashti, H.; Yaghoubian, Y.; Babaeizad, V. Effect of paclobutrazol and Priformospora indica inoculation on antioxidant enzymes activity and morphological characteristics of green beans (Phaseoluse vulgaris L.) in chilling stress. J. Plant Proc. Funct. 2016, 5, 133–146. [Google Scholar]
- Sun, R.-T.; Zhang, Z.-Z.; Feng, X.-C.; Zhou, N.; Feng, H.-D.; Liu, Y.-M.; Harsonowati, W.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.-S. Endophytic fungi accelerate leaf physiological activity and resveratrol accumulation in Polygonum cuspidatum by up-regulating expression of associated genes. Agronomy 2022, 12, 1220. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.A.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Mcgrath, B.M.; Carrell, R.W. Reactions involving superoxide and optimum and unstabl haemoglobins. Biochem. J. 1976, 3, 493–502. [Google Scholar] [CrossRef]
- Rong, Z.-Y.; Jiang, D.-J.; Cao, J.-L.; Hashem, A.; Abd_Allah, E.F.; Alsayed, M.F.; Harsonowati, W.; Wu, Q.-S. Endophytic fungus Serendipita indica accelerates ascorbate-glutathione cycle of white clover in response to water stress. Front. Microbiol. 2022, 13, 967851. [Google Scholar] [CrossRef]
- Obinger, C.; Maj, M.; Nicholls, P.; Loewen, P. Activity, peroxide compound formation, and heme d synthesis in Escherichia coli HPII catalase. Arch. Biochem. Biophys. 1997, 342, 58–67. [Google Scholar] [CrossRef]
- Liu, Z.Y. Regulation role of PcRS and PcPKS1 from Polygonum cuspidatum in the resveratrol biosynthesis. Ph.D. Dissertation, South China University of Technology, Guangzhou, China, 2012; pp. 1–110. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N. Beneficial roles of arbuscular mycorrhizas in citrus seedlings at temperature stress. Sci. Hortic. 2010, 125, 289–293. [Google Scholar] [CrossRef]
- Agurla, S.; Gahir, S.; Munemasa, S.; Murata, Y.; Raghavendra, A.S. Mechanism of stomatal closure in plants exposed to drought and cold stress. Adv. Exp. Med. Biol. 2018, 1081, 215–232. [Google Scholar] [PubMed]
- Tian, X.; Liu, X.Q.; Liu, X.R.; Li, Q.S.; Abd_Allah, E.F.; Wu, Q.S. Mycorrhizal cucumber with Diversispora versiformis has active heat stress tolerance by up-regulating expression of both CsHsp70s and CsPIPs genes. Sci. Hortic. 2023, 319, 112494. [Google Scholar] [CrossRef]
- Murphy, B.R.; Doohan, F.M.; Hodkinson, T.R. Yield increase induced by the fungal root endophyte Piriformospora indica in barley grown at low temperature is nutrient limited. Symbiosis 2014, 62, 29–39. [Google Scholar] [CrossRef]
- Rong, Z.Y.; Zhang, Z.Z.; Alqahtani, M.D.; Wu, Q.S.; Gao, X.B. Serendipita indica is a biostimulant that improves tea growth at low P levels by modulating P acquisition and hormone levels. Rhizosphere 2023, 28, 100796. [Google Scholar] [CrossRef]
- Song, Y.; Feng, L.; Alyafei, M.A.M.; Jaleen, A.; Ren, M. Function of chloroplasts in plant stress responses. Int. J. Mol. Sci. 2021, 22, 13464. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, J.L.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Serendipita indica mitigates drought-triggered oxidative burst in trifoliate orange by stimulating antioxidant defense systems. Front. Plant Sci. 2023, 14, 1247342. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.Z.; Chow, W.S.; Sun, L.L.; Chen, J.W.; Chen, Y.J.; Peng, C.L. The influence of low temperature on photosynthesis and antioxidant enzymes in sensitive banana and tolerant plantain (Musa sp.) cultivars. Photosynthetica 2011, 49, 201–208. [Google Scholar] [CrossRef]
- Ramazan, S.; Qazi, H.A.; Dar, Z.A.; John, R. Low temperature elicits differential biochemical and antioxidant responses in maize (Zea mays) genotypes with different susceptibility to low temperature stress. Physiol. Mol. Biol. Plants 2021, 27, 1395–1412. [Google Scholar] [CrossRef]
- Cao, J.L.; He, W.X.; Zou, Y.N.; Wu, Q.S. An endophytic fungus, Piriformospora indica, enhances drought tolerance of trifoliate orange by modulating the antioxidant defense system and composition of fatty acids. Tree Physiol. 2023, 43, 452–466. [Google Scholar] [CrossRef]
- Liang, S.M.; Hashem, A.; Abd-Allah, E.F.; Wu, Q.S. Root-associated symbiotic fungi enhance waterlogging tolerance of peach seedlings by increasing flavonoids and activities and gene expression of antioxidant enzymes. Chem. Biol. Technol. Agric. 2023, 10, 124. [Google Scholar] [CrossRef]
- Yang, L.; Cao, J.L.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Piriformospora indica: A root endophytic fungus and its roles in plants. Not. Bot. Horti Agrobot. 2020, 48, 1–13. [Google Scholar] [CrossRef]
- Liu, J.L.; Liu, C.; Dong, Q.; Yao, W.; Wu, C. Effects of Piriformospora indica inoculation on gas exchange, biomass accumulation and contents of secondary metabolites of Ocimum gratissimum. Acta Agric. Jiangxi 2021, 33, 19–24. [Google Scholar]
- Arora, M.; Saxena, P.; Choudhary, D.K.; Abdin, M.Z.; Varma, A. Dual symbiosis between Piriformospora indica and Azotobacter chroococcum enhances the artemisinin content in Artemisia annua L. World J. Microbiol. Biotechnol. 2016, 32, 19. [Google Scholar] [CrossRef] [PubMed]
- Li, S.X.; Liu, J.L.; Zhang, N.; Cui, Z. Effects of treatment methods on the amounts of resveratrol and polydatin in different parts of Polygonum cuspidatum. Chin. Tradit. Herb. Drug 2009, 40, 469–471. [Google Scholar]
- Liu, H.J.; Yi, Y.J.; Yang, J.K.; Zhong, Y.L.; Chen, X. Isolation and identification of endophytic fungi capable of transforming polydatin from Polygonum cuspidatum. Food Sci. 2012, 33, 172–176. [Google Scholar]
- Sun, R.T.; Zhang, Z.Z.; Liu, M.Y.; Feng, X.C.; Zhou, N.; Feng, H.D.; Hashem, A.; Abd_Allah, E.F.; Harsonowati, W.; Wu, Q.S. Arbuscular mycorrhizal fungi and phosphorus supply accelerate main medicinal component production of Polygonum cuspidatum. Front. Microbiol. 2022, 13, 1006140. [Google Scholar] [CrossRef]
- Li, X.X.; Liu, C.; Xu, J.X.; Wang, Y.Y.; Wu, X.; Qin, J.B.; Liu, Z.Y. Construction of eukaryotic expression vector of Polygonum cuspidatum gene PcMYB1 and its genetic transforamtion. J. Yangtze Univ. (Nat. Sci. Ed.) 2018, 15, 39–42. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, J.; Chen, Y. Serendipita indica: A Biostimulant Enhancing Low-Temperature Tolerance and Active Constituent Levels in Polygonum cuspidatum. Agriculture 2024, 14, 7. https://doi.org/10.3390/agriculture14010007
Shen J, Chen Y. Serendipita indica: A Biostimulant Enhancing Low-Temperature Tolerance and Active Constituent Levels in Polygonum cuspidatum. Agriculture. 2024; 14(1):7. https://doi.org/10.3390/agriculture14010007
Chicago/Turabian StyleShen, Junhao, and Yongqin Chen. 2024. "Serendipita indica: A Biostimulant Enhancing Low-Temperature Tolerance and Active Constituent Levels in Polygonum cuspidatum" Agriculture 14, no. 1: 7. https://doi.org/10.3390/agriculture14010007