

Alleviation of Stripe Rust Disease in Wheat Seedlings Using Three Different Species of Trichoderma spp.

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design, Greenhouse Conditions, and Trichoderma Treatments
2.3. Preparation of Trichoderma spp.
2.4. Stripe Rust Inoculation and Evaluation
2.5. Scanning Electron Microscope (SEM)
2.6. Statistical Analysis of Stripe Rust and Slow Rust Components
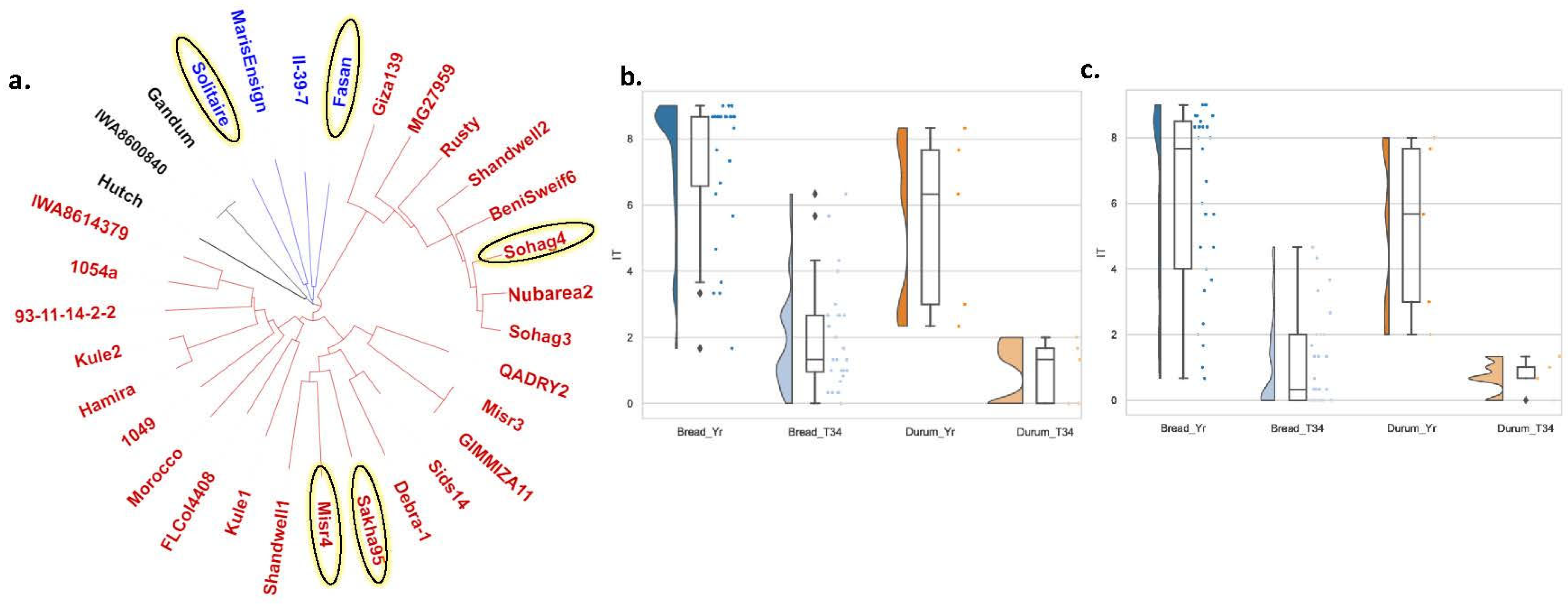
2.7. Genetic Distance between Each Pair of the Tested Genotypes
3. Results
3.1. Effect of the Different Temperatures on Stripe Rust Infection
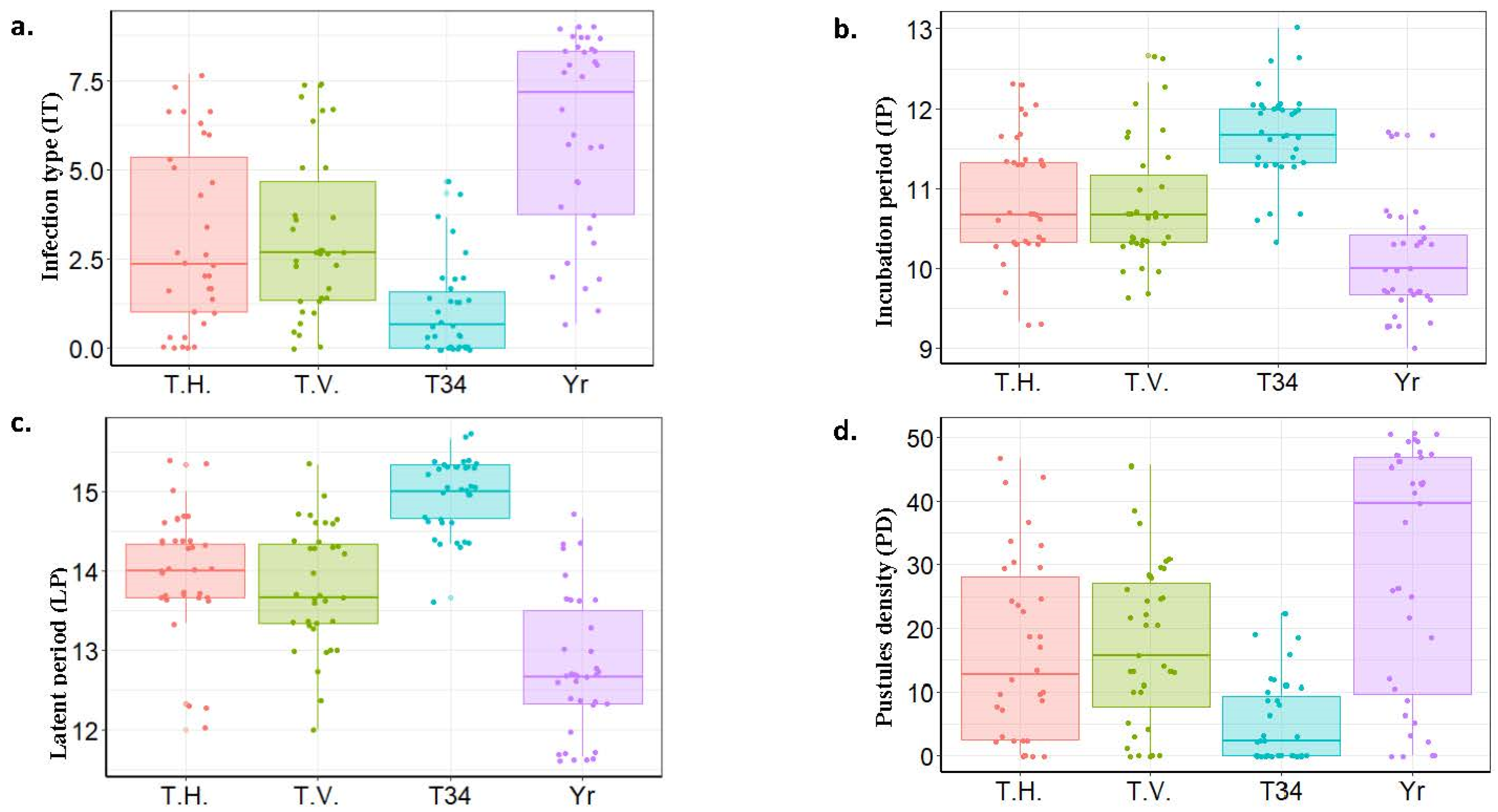
3.2. Effect of the Different Trichoderma spp. on Stripe Rust Symptoms
3.3. Insights into the Effect of T34 on Stripe Rust Disease
3.3.1. Effectiveness of T34 in Controlling Stripe Rust under Different Temperature Degrees
3.3.2. Effectiveness of T34 as a Bioagent Controlling Stripe Rust in Different Wheat Genetic Backgrounds
4. Discussion
4.1. Factor 1: Testing the Effectiveness of Biological Control in Different Environments
4.2. Factor 2: Testing the Effectiveness of Different Bioagents in the Biological Control of WSR
4.3. Factor 3: Testing the Effectiveness of T34 Bioagent in Controlling WSR in Different Wheat Germplasm
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Yuan, F.P.; Zeng, Q.D.; Wu, J.H.; Wang, Q.L.; Yang, Z.J.; Liang, B.P.; Kang, Z.S.; Chen, X.H.; Han, D.J. QTL mapping and validation of adult plant resistance to stripe rust in chinese wheat landrace humai 15. Front. Plant Sci. 2018, 9, 968. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Markell, S.G.; Milus, E.A. Emergence of a novel population of Puccinia striiformis f. sp. tritici in eastern United States. Phytopathology 2008, 98, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Elbasyoni, I.S.; El-Orabey, W.M.; Morsy, S.; Baenziger, P.S.; Al Ajlouni, Z.; Dowikat, I. Evaluation of a global spring wheat panel for stripe rust: Resistance loci validation and novel resources identification. PLoS ONE 2019, 14, e0222755. [Google Scholar] [CrossRef]
- Abou-Zeid, M.A.; Mourad, A.M.I. Genomic regions associated with stripe rust resistance against the Egyptian race revealed by genome-wide association study. BMC Plant Biol. 2021, 21, 42. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Abou-Zeid, M.A.; Eltaher, S.; Baenziger, P.S.; Börner, A. Identification of candidate genes and genomic regions associated with adult plant resistance to stripe rust in spring wheat. Agronomy 2021, 11, 2585. [Google Scholar] [CrossRef]
- Esmail, S.M.; Draz, I.S.; Ashmawy, M.A.; El-Orabey, W.M. Emergence of new aggressive races of Puccinia striiformis f. sp. tritici causing yellow rust epiphytotic in Egypt. Physiol. Mol. Plant Pathol. 2021, 114, 101612. [Google Scholar] [CrossRef]
- Draz, I.; Esmail, S.; Abou-zeid, M.; Hafez, Y. Changeability in stripe rust infection and grainy Yield of wheat associated to climatic conditions. Environ. Biodiv. Soil Secur. 2019, 2, 143–153. [Google Scholar] [CrossRef]
- Aoun, M.; Chen, X.; Somo, M.; Xu, S.S.; Li, X.; Elias, E.M. Novel stripe rust all-stage resistance loci identified in a worldwide collection of durum wheat using genome-wide association mapping. Plant Genome 2021, 14, e20136. [Google Scholar] [CrossRef]
- Mahmood, Z.; Ali, M.; Mirza, J.I.; Fayyaz, M.; Majeed, K.; Naeem, M.K.; Aziz, A.; Trethowan, R.; Ogbonnaya, F.C.; Poland, J.; et al. Genome-Wide Association and Genomic Prediction for Stripe Rust Resistance in Synthetic-Derived Wheats. Front. Plant Sci. 2022, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Bouvet, L.; Percival-Alwyn, L.; Berry, S.; Fenwick, P.; Mantello, C.C.; Sharma, R.; Holdgate, S.; Mackay, I.J.; Cockram, J. Wheat genetic loci conferring resistance to stripe rust in the face of genetically diverse races of the fungus Puccinia striiformis f. sp. tritici. Theor. Appl. Genet. 2022, 135, 301–319. [Google Scholar] [CrossRef]
- Jan, I.; Saripalli, G.; Kumar, K.; Kumar, A.; Singh, R.; Batra, R.; Sharma, P.K.; Balyan, H.S.; Gupta, P.K. Meta-QTLs and candidate genes for stripe rust resistance in wheat. Sci. Rep. 2021, 11, 22923. [Google Scholar] [CrossRef]
- Tene, M.; Adhikari, E.; Cobo, N.; Jordan, K.W.; Matny, O.; del Blanco, I.A.; Roter, J.; Ezrati, S.; Govta, L.; Manisterski, J.; et al. GWAS for Stripe Rust Resistance in Wild Emmer Wheat (Triticum dicoccoides) Population: Obstacles and Solutions. Crops 2022, 2, 42–61. [Google Scholar] [CrossRef]
- Maccaferri, M.; Zhang, J.; Bulli, P.; Abate, Z.; Chao, S.; Cantu, D.; Bossolini, E.; Chen, X.; Pumphrey, M.; Dubcovsky, J. A Genome-wide association study of resistance to stripe rust (Puccinia striiformis f. sp. tritici) in a worldwide collection of hexaploid spring wheat (Triticum aestivum L.). G3 Genes|Genomes|Genet. 2015, 5, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Alemui, S.K.; Huluka, A.B.; Tesfayei, K.; Haileselassie, T.; Uauy, C. Genome-wide association mapping identifies yellow rust resistance loci in Ethiopian durum wheat germplasm. PLoS ONE 2021, 16, e0243675. [Google Scholar] [CrossRef]
- El Messoadi, K.; El Hanafi, S.; El Gataa, Z.; Kehel, Z.; Bouhouch, Y.; Tadesse, W. Genome wide association study for stripe rust resistance in spring bread wheat (Triticum aestivum L.). J. Plant Pathol. 2022, 104, 1049–1059. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, C.; Cheng, Y.; Yao, F.; Long, L.; Wu, Y.; Li, J.; Li, H.; Wang, J.; Jiang, Q.; et al. Genome-wide association mapping reveals potential novel loci controlling stripe rust resistance in a Chinese wheat landrace diversity panel from the southern autumn-sown spring wheat zone. BMC Genom. 2021, 22, 34. [Google Scholar] [CrossRef]
- Shahinnia, F.; Geyer, M.; Schürmann, F.; Rudolphi, S.; Holzapfel, J.; Kempf, H.; Stadlmeier, M.; Löschenberger, F.; Morales, L.; Buerstmayr, H.; et al. Genome-wide association study and genomic prediction of resistance to stripe rust in current Central and Northern European winter wheat germplasm. Theor. Appl. Genet. 2022, 135, 3583–3595. [Google Scholar] [CrossRef]
- Zhang, P.; Yan, X.; Weldu, T.; Zhou, Y.; Yang, E.; Xia, X.; He, Z.; Li, Z.; Liu, D. Genome-wide association mapping of leaf rust and stripe rust resistance in wheat accessions using the 90K SNP array. Theor. Appl. Genet. 2021, 134, 1233–1251. [Google Scholar] [CrossRef]
- Yao, F.; Guan, F.; Duan, L.; Long, L.; Tang, H.; Jiang, Y.; Li, H.; Jiang, Q.; Wang, J.; Qi, P.; et al. Genome-Wide Association Analysis of Stable Stripe Rust Resistance Loci in a Chinese Wheat Landrace Panel Using the 660K SNP Array. Front. Plant Sci. 2021, 12, 783830. [Google Scholar] [CrossRef]
- Bhavani, S.; Singh, P.K.; Qureshi, N.; He, X.; Biswal, A.K.; Juliana, P.; Dababat, A.; Mourad, A.M.I. Globally Important Wheat Diseases: Status, Challenges, Breeding and Genomic Tools to Enhance Resistance Durability. In Genomic Designing for Biotic Stress Resistant Cereal Crops; Kole, C., Ed.; Springer International Publishing: New Delhi, India, 2021; pp. 59–128. ISBN 9783030758790. [Google Scholar]
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. BioControl 2018, 63, 155–167. [Google Scholar] [CrossRef]
- Messing, R.; Brodeur, J. Current challenges to the implementation of classical biological control. BioControl 2018, 63, 1–9. [Google Scholar] [CrossRef]
- Brodeur, J.; Abram, P.K.; Heimpel, G.E.; Messing, R.H. Trends in biological control: Public interest, international networking and research direction. BioControl 2018, 63, 11–26. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Promputtha, I. Fungi vs. Fungi in Biocontrol: An Overview of Fungal Antagonists Applied Against Fungal Plant Pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, T.; Rajan, S.; Manoharan, M.; Gopal, R. Biological Management of Banana Fusarium Wilt Caused by Fusarium oxysporum f. sp. cubense Tropical Race 4 Using Antagonistic Fungal Isolate CSR-T-3 (Trichoderma reesei). Front. Microbiol. 2020, 11, 595845. [Google Scholar] [CrossRef]
- Pocurull, M.; Fullana, A.M.; Ferro, M.; Valero, P.; Escudero, N.; Saus, E.; Gabaldón, T.; Sorribas, F.J. Commercial Formulates of Trichoderma Induce Systemic Plant Resistance to Meloidogyne incognita in Tomato and the Effect Is Additive to That of the Mi-1.2 Resistance Gene. Front. Microbiol. 2020, 10, 3042. [Google Scholar] [CrossRef]
- La Spada, F.; Stracquadanio, C.; Riolo, M.; Pane, A.; Cacciola, S.O. Trichoderma Counteracts the Challenge of Phytophthora nicotianae Infections on Tomato by Modulating Plant Defense Mechanisms and the Expression of Crinkler, Necrosis-Inducing Phytophthora Protein 1, and Cellulose-Binding Elicitor Lectin Pathogenic Effecto. Front. Plant Sci. 2020, 11, 583539. [Google Scholar] [CrossRef]
- De Medeiros, H.A.; Vieira, J.; Filho, D.A.; Freitas, L.G. De Tomato progeny inherit resistance to the nematode Meloidogyne javanica linked to plant growth induced by the biocontrol fungus Trichoderma atroviride. Nat. Publ. Gr. 2017, 7, 40216. [Google Scholar] [CrossRef]
- Sallam, N.M.A.; Eraky, A.M.I.; Sallam, A. Effect of Trichoderma spp. on Fusarium wilt disease of tomato. Mol. Biol. Rep. 2019, 46, 4463–4470. [Google Scholar] [CrossRef] [PubMed]
- Morán-Diez, M.E.; Martínez de Alba, Á.E.; Rubio, M.B.; Hermosa, R.; Monte, E. Trichoderma and the plant heritable priming responses. J. Fungi 2021, 7, 318. [Google Scholar] [CrossRef] [PubMed]
- Esmail, S.M.; Omar, G.E.; Mourad, A. In-depth understanding of the genetic control of stripe rust resistance (Puccinia striiformis f. sp. tritici) induced in wheat (Triticum aestivum L.) by Trichoderma asperellum T34. Plant Dis. 2023, 107, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Kiani, T.; Mehboob, F.; Hyder, M.Z.; Zainy, Z.; Xu, L.; Huang, L.; Farrakh, S. Control of stripe rust of wheat using indigenous endophytic bacteria at seedling and adult plant stage. Sci. Rep. 2021, 11, 14473. [Google Scholar] [CrossRef] [PubMed]
- Omara, R.I.; Essa, T.A.; Khalil, A.A.; Elsharkawy, M.M. A case study of non-traditional treatments for the control of wheat stem rust disease. Egypt. J. Biol. Pest Control 2020, 30, 83. [Google Scholar] [CrossRef]
- El-Sharkawy, H.H.A.; Rashad, Y.M.; Ibrahim, S.A. Biocontrol of stem rust disease of wheat using arbuscular mycorrhizal fungi and Trichoderma spp. Physiol. Mol. Plant Pathol. 2018, 103, 84–91. [Google Scholar] [CrossRef]
- Hafez, Y.; Abdelfatah, A.; El-Nashar, F.; Badr, M.; Elkady, S. Management of barley net blotch using Trichoderma asperellum (T34), eugenol, non-traditional compounds and fungicides. Egypt. J. Biol. Pest Control 2019, 29, 88. [Google Scholar] [CrossRef]
- Degani, O.; Gordani, A. New Antifungal Compound, 6-Pentyl-α-Pyrone, against the Maize Late Wilt Pathogen, Magnaporthiopsis maydis. Agronomy 2022, 12, 2339. [Google Scholar] [CrossRef]
- Abdel-Fattah, G.M.; Shabana, Y.M.; Ismail, A.E.; Rashad, Y.M. Trichoderma harzianum: A biocontrol agent against Bipolaris oryzae. Mycopathologia 2007, 164, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Maheshwari, V. Biological seed treatment for the control of loose smut of wheat. Indian Phytopathol. 2001, 54, 457–460. [Google Scholar]
- Bayoumi, Y.; Taha, N.; Shalaby, T.; Alshaal, T.; El-Ramady, H. Sulfur promotes biocontrol of purple blotch disease via Trichoderma spp. and enhances the growth, yield and quality of onion. Appl. Soil Ecol. 2019, 134, 15–24. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Marchi, A. Control of storage rots on various pear cultivars with a saprophytic strain of Pseudomonas syringae. Plant Dis. 1992, 76, 555–560. [Google Scholar] [CrossRef]
- Stakman, E.C.; Stewart, D.M.; Loegering, W.Q. Identification of Physiologic Races of Puccinia Graminis var. tritici; USDA_ARS: Washington, DC, USA, 1962. [Google Scholar]
- McNeal, F.H.; Konzak, C.F.; Smith, E.P.; Tate, W.S.; Russell, T.S. A Uniform System for Recording and Processing Cereal Research Data; U.S. Department of Agriculture, Agricultural Research Service, ARS: Washington, DC, USA, 1971. [Google Scholar]
- Katsuya, K.; Green, G.J. Reproductive potentials of races 15B and 56 of wheat stem rust. Can. J. Bot. 1967, 45, 1077–1091. [Google Scholar] [CrossRef]
- Parlevliet, J.E.; Ommeren, A.V. Partial resistance of barley to leaf rust, Puccinia hordei. II. Relationship between field trials, micro plot tests and latent period. Euphytica 1975, 24, 293–303. [Google Scholar] [CrossRef]
- Harley, M.; Fergusen, L. Thee of SEM in pollen morphology and plant systemic. In Scanning Electron Microscopy Studies in Taxonomy and Functional Morphology; Clarendon Press: Oxford, UK, 1990; pp. 45–68. [Google Scholar]
- Utz, H. PLABSTAT: A Computer Program for Statistical Analysis of Plant Breeding Experiments; Institute of Plant Breeding, Seed Science and Population Genetics: Hohenheim, Germany, 1997. [Google Scholar]
- Aleksandrov, V.; Kartseva, T.; Alqudah, A.M.; Kocheva, K.; Tasheva, K.; Börner, A.; Misheva, S. Genetic diversity, linkage disequilibrium and population structure of bulgarian bread wheat assessed by genome-wide distributed SNP markers: From old germplasm to semi-dwarf cultivars. Plants 2021, 10, 1116. [Google Scholar] [CrossRef] [PubMed]
- Amro, A.; Harb, S.; Youssef, K.; Ali, M.M.F.; Mohammed, A.G.; Mourad, A.M.I.; Afifi, M.; Börner, A.; Sallam, A. Growth responses and genetic variation among highly ecologically diverse spring wheat genotypes grown under seawater stress. Front. Plant Sci. 2022, 13, 996538. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Eltaher, S.; Börner, A.; Sallam, A. Unlocking the genetic control of spring wheat kernel traits under normal and heavy metals stress conditions. Plant Soil 2023, 484, 257–278. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.B. The ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Cordo, C.A.; Monaco, C.I.; Segarra, C.I.; Simon, M.R.; Mansilla, A.Y.; Perelló, A.E.; Kripelz, N.I.; Bayo, D.; Conde, R.D. Trichoderma spp. as elicitors of wheat plant defense responses against Septoria tritici. Biocontrol Sci. Technol. 2007, 17, 687–698. [Google Scholar] [CrossRef]
- Boamah, S.; Zhang, S.; Xu, B.; Li, T.; Calderón-Urrea, A. Trichoderma longibrachiatum (TG1) Enhances Wheat Seedlings Tolerance to Salt Stress and Resistance to Fusarium pseudograminearum. Front. Plant Sci. 2021, 12, 741231. [Google Scholar] [CrossRef] [PubMed]
- Vitti, A.; Bevilacqua, V.; Logozzo, G.; Bochicchio, R.; Amato, M.; Nuzzaci, M. Seed Coating with Trichoderma harzianum T-22 of Italian Durum Wheat Increases Protection against Fusarium culmorum-Induced Crown Rot. Agriculture 2022, 12, 714. [Google Scholar] [CrossRef]
- Segarra, G.; Van Der Ent, S.; Trillas, I.; Pieterse, C.M.J. MYB72, a node of convergence in induced systemic resistance triggered by a fungal and a bacterial beneficial microbe. Plant Biol. 2009, 11, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Hafez, Y.M.; El-Nagar, A.S.; Elzaawely, A.A.; Kamel, S.; Maswada, H.F. Biological control of podosphaera xanthii the causal agent of squash powdery mildew disease by upregulation of defense-related enzymes. Egypt. J. Biol. Pest Control 2018, 28, 57. [Google Scholar] [CrossRef]
- Ali, S.; Khan, M.I.; Shahbaz, M.; Nabi, G.; Zeeshan, M.; Aleem, S.; Hussain, M.; Saadia. Environmental Factors and Yellow Rust Epidemic on Wheat Varieties in Punjab, Pakistan. J. Agric. Food 2022, 3, 86–96. [Google Scholar]
- Grabow, B.S.; Shah, D.A.; DeWolf, E.D. Environmental conditions associated with stripe rust in kansas winter wheat, Kansas State University. Plant Dis. 2016, 100, 2306–2312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Liao, Z.; Wu, S.; Nobis, M.P.; Wang, J.; Wu, N. Impact of climate change on wheat security through an alternate host of stripe rust. Food Energy Secur. 2022, 11, e356. [Google Scholar] [CrossRef]
- Knott, D. The Wheat Rusts—Breeding for Resistance; Frnakel, R., Grossma, M., Mgliga, P., Eds.; Springer: Berlin/Heidelberg, Germany, 1989; ISBN 9781425803780. [Google Scholar]
- Hovmøller, M.S.; Walter, S.; Bayles, R.A.; Hubbard, A.; Flath, K.; Sommerfeldt, N.; Leconte, M.; Czembor, P.; Rodriguez-Algaba, J.; Thach, T.; et al. Replacement of the European wheat yellow rust population by new races from the centre of diversity in the near-Himalayan region. Plant Pathol. 2016, 65, 402–411. [Google Scholar] [CrossRef]
- Hubbard, A.; Lewis, C.M.; Yoshida, K.; Ramirez-Gonzalez, R.H.; de Vallavieille-Pope, C.; Thomas, J.; Kamoun, S.; Bayles, R.; Uauy, C.; Saunders, D.G.O. Field pathogenomics reveals the emergence of a diverse wheat yellow rust population. Genome Biol. 2015, 16, 23. [Google Scholar] [CrossRef] [PubMed]
- Milus, E.A.; Kristensen, K.; Hovmøller, M.S. Evidence for increased aggressiveness in a recent widespread strain of Puccinia striiformis f. sp. tritici causing stripe rust of wheat. Phytopathology 2009, 99, 89–94. [Google Scholar] [CrossRef] [PubMed]
- El-Kazzaz, M.K.; Ghoniem, K.E.; Ashmawy, M.A.; Omar, G.E.; Hafez, Y.M. Suppression of wheat strip rust disease caused by Puccinia striiformis f. sp. Tritici by eco-friendly bio-control agents correlated with yield improvement. Fresenius Environ. Bull. 2020, 29, 8385–8393. [Google Scholar]
- Mahmoud, A.F. Genetic variation and biological control of Fusarium graminearum isolated from wheat in Assiut-Egypt. Plant Pathol. J. 2016, 32, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Omara, R.I.; El-Kot, G.A.; Fadel, F.M.; Abdelaal, K.A.A.; Saleh, E.M. Efficacy of certain bioagents on patho-physiological characters of wheat plants under wheat leaf rust stress. Physiol. Mol. Plant Pathol. 2019, 106, 102–108. [Google Scholar] [CrossRef]
- Martanto, E.A.; Tanati, A.E.; Baan, S.; Tata, H.R.; Murdjoko, A. Effectiveness of biological control of trichoderma harzianum on soybean leaf rust disease and the production in west papua lowland, indonesia. Biodiversitas 2020, 21, 1935–1939. [Google Scholar] [CrossRef]
- Afzal, S.; Haroon, A.; Hussain, M.A.; Chaudary, A.-U.-R.; Bashir, M.A.; Atta, S.; Bashir, S.; Bodlah, M.A. Potential of Trichoderma Isolates to Control Plant Pathogen, Leaf Rust on Different Commercial Wheat Varieties/Genotypes. In Wheat-Recent Advances; Ansari, M.-R., Ed.; IntechOpen: London, UK, 2022; pp. 197–212. ISBN 978-1-80355-523-2. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.O.V. | d.f. | M.S. |
|---|---|---|
| Experiment (E) | 1 | 45.49 ** |
| Replications (R) | 2 | 0.115 |
| Genotypes (G) | 33 | 29.44 ** |
| GE | 32 | 9.13 ** |
| GRE | 132 | 0.45 |
| Total | 200 | -- |
| Heritability | 0.69 | |
| Trait | IT | IP | LP | PD | ||||
|---|---|---|---|---|---|---|---|---|
| S.O.V. | d.f. | M.S. | d.f. | M.S. | d.f. | M.S. | d.f. | M.S. |
| Treatment (T) | 3 | 470.95 ** | 3 | 36.57 ** | 3 | 62.03 ** | 3 | 9270.70 ** |
| Replications (R) | 2 | 0.53 | 2 | 0.72 | 2 | 10.43 ** | 2 | 41.62 * |
| Genotypes (G) | 32 | 42.30 ** | 32 | 9.12 ** | 32 | 7.67 ** | 32 | 1308.01 ** |
| GT | 95 | 7.30 ** | 95 | 1.07 ** | 95 | 1.85 ** | 95 | 228.95 ** |
| GRT | 260 | 0.98 | 260 | 0.35 | 260 | 0.39 | 260 | 13.4477 |
| Total | 392 | - | 392 | - | 392 | |||
| Trait | IT | IP | LP | PD | ||||
|---|---|---|---|---|---|---|---|---|
| S.O.V. | d.f. | M.S. | d.f. | M.S. | d.f. | M.S. | d.f. | M.S. |
| Treatment (T) | 3 | 425.61 ** | 3 | 44.74 ** | 3 | 79.94 ** | 3 | 10879.96 ** |
| Replications (R) | 2 | 0.18 | 2 | 1.23 * | 2 | 2.72 ** | 2 | 31.63 ** |
| Genotypes (G) | 33 | 43.55 ** | 33 | 4.27 ** | 33 | 4.28 ** | 33 | 1533.03 ** |
| GT | 98 | 6.65 ** | 98 | 0.72 ** | 98 | 0.79 ** | 98 | 250.17 ** |
| GRT | 267 | 0.42 | 268 | 0.28 | 268 | 0.31 | 268 | 10.4896 |
| Total | 403 | - | 404 | 404 | 404 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mourad, A.M.I.; Börner, A.; Esmail, S.M. Alleviation of Stripe Rust Disease in Wheat Seedlings Using Three Different Species of Trichoderma spp. Agriculture 2023, 13, 1755. https://doi.org/10.3390/agriculture13091755
Mourad AMI, Börner A, Esmail SM. Alleviation of Stripe Rust Disease in Wheat Seedlings Using Three Different Species of Trichoderma spp. Agriculture. 2023; 13(9):1755. https://doi.org/10.3390/agriculture13091755
Chicago/Turabian StyleMourad, Amira M. I., Andreas Börner, and Samar M. Esmail. 2023. "Alleviation of Stripe Rust Disease in Wheat Seedlings Using Three Different Species of Trichoderma spp." Agriculture 13, no. 9: 1755. https://doi.org/10.3390/agriculture13091755