1. Introduction
Xylella fastidiosa (
X.f.) is a globally distributed Gram-negative bacterium hosted by a wide range of plant species, including olive trees, grapes, almonds, fig, citrus, ornamentals (oleander, elm, periwinkle, and oak), and some wild plants [
1]. In olives, it causes olive quick decline syndrome (OQDS), which consists of leaf scorch and the desiccation of terminal branches that rapidly spread to the rest of the canopy, even leading to tree death [
2] as the xylem vessels are obstructed by the accumulation of bacterial biofilms [
3]. The global spread of this pathogen continues to increase via the transport of commodities and plant material (EPPO Global Database). In 2013,
X. fastidiosa subspecies
pauca strain ST53 was identified for the first time in the Apulia region (southern Italy) as an agent of the severe epidemic that caused widespread desiccation and tree mortality in the olive groves of Salento [
2,
4,
5]. The meadow spittlebug
Philaenus spumarius was identified as the vector of the bacterium and is responsible for the rapid spread of the pathogen from its original
foci area to the Apulia region [
6,
7,
8,
9].
The difficulty in determining the origin and actual impact of the damage caused by the disease slowed down sanitary efforts, resulting in the spread of the bacterium, which was also promoted by favorable climatic conditions [
10]. Therefore, the spread of the epidemic has led to the loss of millions of olive trees, causing severe damage to the Apulian economy and a dramatic change in the landscape [
11,
12]. So far, various efforts have been made to prevent the disease and to control the pathogen, but these have only led to a slowing down of the epidemic and not to the containment of the disease or the eradication of the bacterium [
1]. According to European and Italian legislation, the containment of
X.f. consists of monitoring the infection status of plants in the buffer zone and the life stage of the meadow spittlebug in order to plan agronomical and chemical control measures [
1,
13].
The rapid spread of
X.f. was favored by several co-factors related to biological, social, and climatic aspects. Indeed, olive tree cultivation plays an important social and symbolic role in the Apulia region, as ancient trees characterize the landscape of the region (e.g., the Valley of Millenary's olive trees in the area of Ostuni, Monopoli, and Fasano 40°43′ N; 17°34′ E), and the area is recognized as a World Heritage Site by UNESCO. Among the most commonly cultivated olive cultivars, Cellina di Nardò and Ogliarola Salentina proved to be the most susceptible, while Leccino and FS17 (also called Favolosa) are resistant to
X.f. [
14,
15].
Some studies have attempted to decipher the mechanisms underlying the different responses of Leccino and FS17 compared to susceptible cultivars [
16,
17,
18]. However, a comprehensive elucidation of the resistance mechanisms is still pending.
Current legislation restricts the conversion of infected areas with respect to Leccino and FS17 varieties only (Commission Implementing Decision (EU) 2015/789;
https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32015D0789, accessed on 28 July 2023). However, the selection of accessions that combine resistance or tolerance to the bacterium with other economically relevant traits could be a valuable source for the conversion of infected areas into resilient agroecosystems [
19]. In this scenario, the use of large olive germplasm collections, including modern and ancient genetic material, is a useful approach for identifying new sources of resistance [
20,
21,
22].
There are more than 900 olive cultivars in the Mediterranean basin [
23] and a large number of ecotypes, local genotypes, and wild trees (
Olea europaea subsp.
europaea var.
sylvestris) [
24]. However, the olive germplasm is characterized by high morphological and genetic variability and has many synonyms and homonyms due to misnaming by local farmers. To exploit olive biodiversity for the selection of agronomic traits and pathogen resistance, the accurate identification of available genotypes is required. Recently, the use of molecular markers and the development of next-generation sequencing (NGS) methods, which allow more reliable identification of cultivars, have replaced morphological characterization, which is strongly influenced by environmental factors [
25,
26,
27]. Among molecular markers, simple sequence repeats (SSRs) are the tools of choice in olives due to their codominant nature, high reproducibility, ease of use, and low cost [
28,
29,
30,
31].
The identification of new putatively resistant genotypes among the local olive germplasm would limit the loss of cultivars with interesting agronomic traits, contributing to maintaining a high genetic diversity and helping to preserve the Apulian olive growing tradition by allowing replanting in infected areas. Moreover, these genotypes could play an important role in the studies of the mechanisms involved in tolerance relative to X.f. The aim of this study was to search for new genotypes that are tolerant or resistant to X.f. in addition to the Leccino and FS17 cultivars. Therefore, a collection of 100 olive genotypes established in 2017 within the RedOXy regional project was molecularly characterized using a set of 10 SSR markers and evaluated for resistance to the pathogen.
4. Discussion
The first outbreak of the quarantine pathogen
Xylella fastidiosa in the EU was detected on olive trees in Apulia in 2013 [
4]. Since then, the disease has spread widely and caused severe landscape and economic damage. So far, only the Leccino and FS17 varieties are considered resistant [
14]. However, in recent years, several regional and national projects have been carried out to identify new sources of tolerance/resistance to
X.f. in order to be used for replanting in infected areas by characterizing local accessions and studying their response to the pathogen [
19]. In 2017, an evaluation program was implemented for this purpose by the University of Bari Aldo Moro in the infected area, where one hundred cultivars/accessions were studied in a trial with randomized blocks of three replicates of four plants.
4.1. Genetic Diversity Assessment
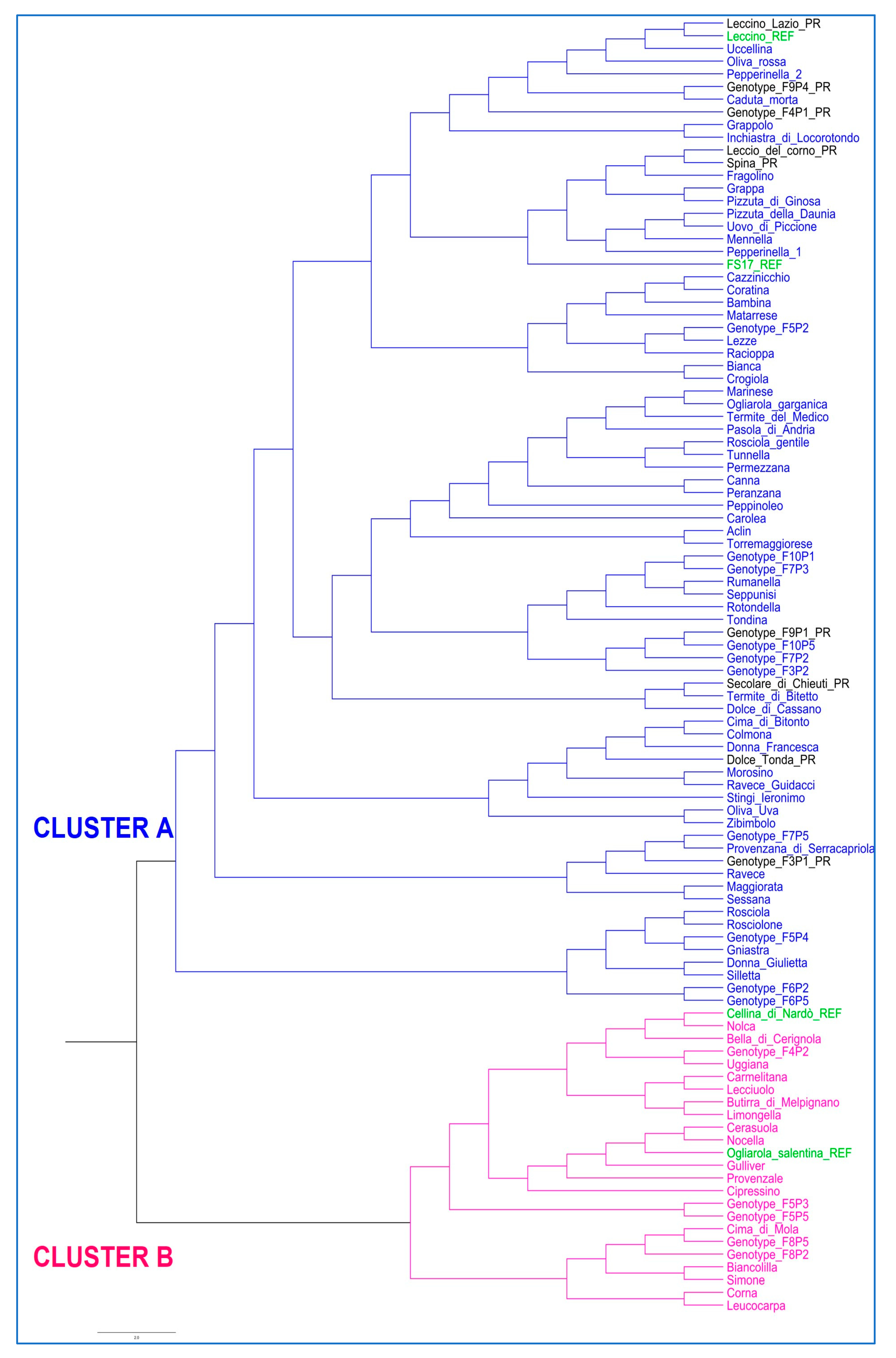
The 100 genotypes were genetically characterized with a set of 10 SSR molecular markers routinely used for olive genotyping, and the data were used to study the genetic relationships between them and the resistant and susceptible reference varieties. The only cases of synonymy revealed by the LRM analysis refer to four varieties in the province of Foggia (Northern Apulia): “Pepperinnella 1-Ravece Guidacci” and “Morosino-Pizzuta della Daunia”. It is likely that the misnaming is due to differences in morphology and use (
Table 1), which led local farmers to consider these varieties as different. In addition, the LRM analysis showed a clear differentiation of most varieties, although some pairs of varieties were strongly related, having pairwise values > 0.40, such as the pairs “Lezze-Racioppa”, “Grappa-Pizzuta di Ginosa”, and “Rosciola-Rotondella”. These results are in line with those of Miazzi et al. (2020) for Apulian varieties. It is likely that during the process of selection, which occurred within the Apulian agroecosystem, the local varieties were derived from crosses among selected trees or pollen coming from feral or wild olive trees [
45,
46], indicating the importance of the local role in the diversification process [
47]. This can be deduced by the results of the parental analysis for the nineteen F
1 genotypes derived from the open pollination of the Simone variety. Any F
1 genotypes appeared to be derived from Simone, thus they probably were derived from crosses with other local varieties. It is likely that the Simone variety used, although certified, was misidentified. This underlines the need to improve the protocols for the certification and marketing of olive varieties [
48]. Despite these results, these F
1 genotypes were retained in our analysis as carrying interesting agronomic traits.
4.2. Evaluation of the Response to X.f. Infection
Infection was monitored using quantitative real-time polymerase chain reaction (qPCR), a diagnostic tool that can detect the pathogen in the early stages of the disease even if the infected plant does not yet show symptoms [
49]. The first assay was performed in 2021, three years after infection with the pathogen. No symptoms were observed during the visual assessment in the first two years (2018–2020). In the third year of assessment, almost all samples showed no symptoms, with the exception of Gulliver and Rosciola Gentile. This was expected as OQDS has a slow progression [
50]. The evaluation performed on the second date, five years after inoculation, confirmed the resistance of the cultivar Leccino and the susceptibility of the cultivar Cellina di Nardò [
15,
51,
52]. Based on the Cq value and the symptomatology, nine accessions could be classified as putatively resistant (PR) in the second assessment. These were Secolare di Chieuti, Spina, Leccio del Corno, Dolce Tonda, and Leccino Lazio and genotypes F
4P
1, F
9P
4, F
3P
1, and F
9P
1. Among them are the accessions Dolce Tonda, F
4P
1, and F
9P
1, which have Cq values of 30.72, 28.04, and 31.67, respectively, and they do not show symptoms that deserve more attention. The PR accessions will need to be further characterized as they represent a valuable resource for studying the mechanisms involved in the response to the
X.f. pathogen.
The susceptible reference cultivar Cellina di Nardò did not seem to be the most susceptible cultivar among the studied genotypes. In fact, the lowest Cq value and the highest CFU/mL value were observed in the cultivar Lezze, although Cellina di Nardò had a symptom score of 2 and Lezze had a symptom score of 1. A discrepancy between the Cq value and symptomatology has been noted before. Studies conducted on different plant species, such as plum, coffee, citrus, and grapevine, have shown that sometimes the symptomatology does not reflect pathogen concentrations [
53,
54,
55,
56]. The intensification of leaf scorch symptoms during
X.f. pathogenesis has been shown to be due to several factors related to the physiological status of the plant. For example, some growth regulators, such as ethylene, can stimulate and accelerate leaf senescence, which exacerbates the symptomatology associated with OQDS [
57,
58]. This could partly explain the leaf scorch symptoms in trees with low concentrations of pathogens and the behavior of PR genotypes Spina, F
9P
4, and F
3P
1, which show a Cq > 27 despite the symptom score of 2. These results show that it is necessary to understand in detail the impact of the developmental status of plants on the manifestation of the symptomatology due to
X.f. infection.
Symptom scores consistent with the Cq values were found for the cultivars Torremaggiorese, Bianca, and Leucocarpa, which had low Cq values and a high incidence of symptoms due to
X.f. (symptoms scores between 4 and 5). For this reason, these accessions can be considered highly susceptible to the bacterium. At the same time, varieties Secolare di Chieuti, Leccio del Corno, Dolce Tonda, and Leccino Lazio and PR genotypes F
4P
1 and F
9P
1 showed no or only minor symptoms (score of 1). The symptomatology of the cultivar Leccino Lazio, albeit to a lesser extent (symptoms score of 1), is in line with the results of an earlier study [
59], confirming that the resistant Leccino cultivar can also show disease symptoms.
According to [
60], plants that consistently show positive qPCR results at 6 and 12 months after inoculation can be considered systemically infected. In the study, Cellina di Nardò, Leccino, and FS17 cultivars all had Cq values of less than 22.05 but a completely different symptomatology. The authors concluded that, despite a similar bacterial load, the different host responses to bacterial infections were due to variations in the physiological state of the plant rather than the direct influence of the pathogen's abundance [
60].
In most of the genotypes studied, the Cq value decreases over the years, indicating an increasing bacterial load over time. However, eleven genotypes showed a constant Cq value, and nine genotypes showed an increasing trend. We hypothesized that the increase in Cq values from 2021 to 2023 may depend on the sampling of the plant material, which may influence the qPCR assay's result; indeed, the different concentrations and spatial variability of the pathogen in the plant could be due to the irregular distribution of xylem vessels [
61]. Thus, future studies need to be performed by sampling larger portions of the canopy in order to increase the reliability of the detection method [
56].
4.3. Comparison between Genetic Data and the Response to X.f.
A comparison between genetic information from the analyzed accessions and their response to the pathogen can provide information on the role of the genetic background with respect to susceptibility/resistance to X.f.
LRM analysis highlighted the genetic similarity of five pairs of accessions (
Table 2). In these samples, such as the Rosciola and Rotondella varieties, we found a similar response to the bacterium (Cq of 21.25 and 20.93 and a symptomatic score of 3) (
Table 4), indicating a possible correlation between the genetic background and the response to the infection.
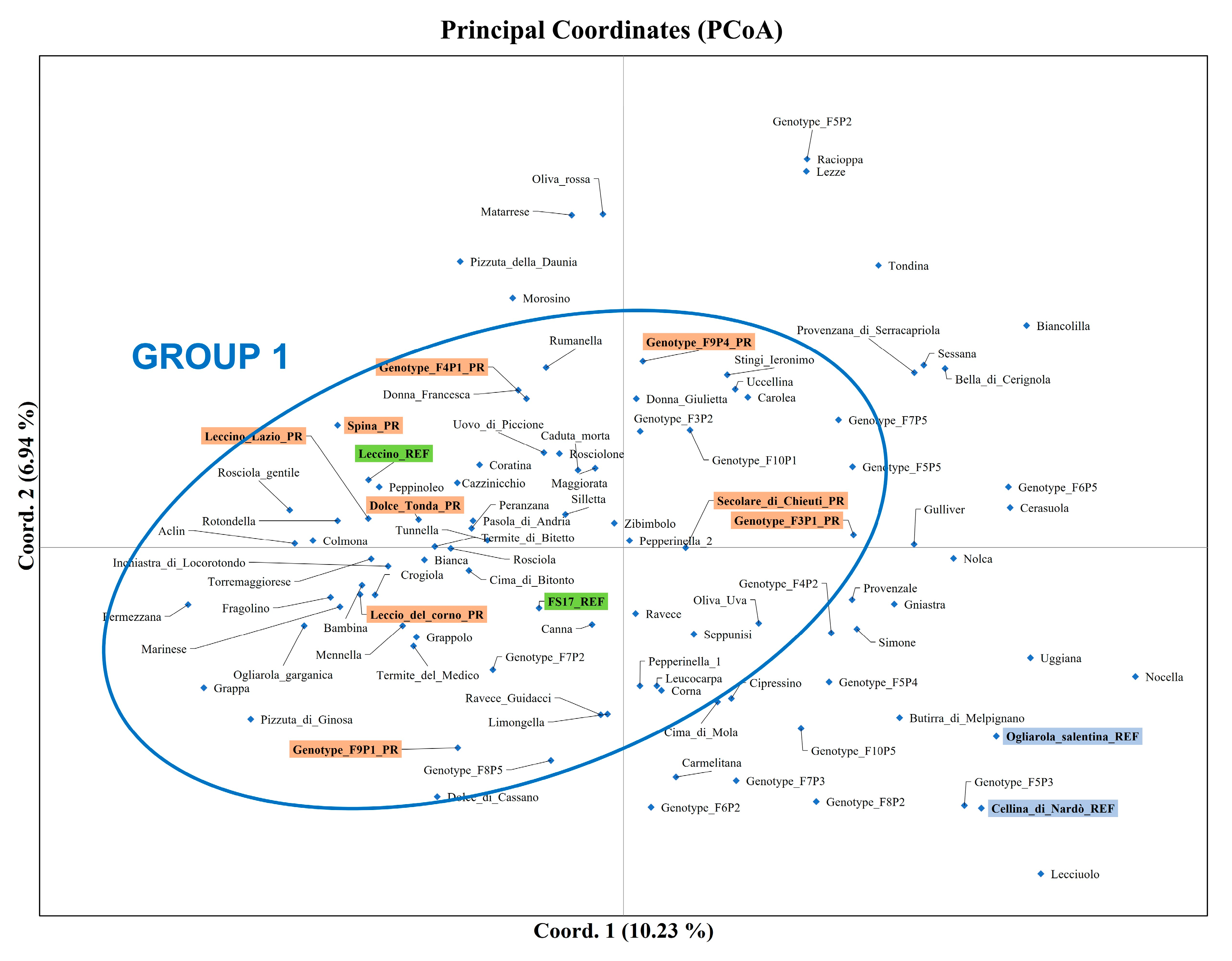
The genetic relationships between the analyzed genotypes, investigated using phylogenetic analysis and PCoA, showed similar results, separating the two resistant references Leccino and FS17 and five PR accessions from the two susceptible varieties Cellina di Nardò and Ogliarola Salentina. Among PR accessions, F
1 genotypes F
4P
1 and F
9P
1 were highly resistant to
X.f. infection. Interestingly, a putative parent of F
4P
1 was identified as Leccino, which is presumed to be responsible for its tolerance, while the putative parent of F
9P
1 was the susceptible Caduta Morta variety (Cq value of 23.48 and a symptom score of 1 at the second time point of evaluation). The great variability in the response to bacterial infection shown by F
1 genotypes could be the result of the genetic recombinations in the progenies. Likewise, the promising PR accession Dolce Tonda had the highly susceptible Coratina variety as a putative parent. This is not surprising because the resistant FS17 cultivar also has the partially susceptible Frantoio as a parent [
62,
63]. Both F
4P
1 and F
9P
1 will require further investigation in order to identify both parents and to elucidate the mechanisms involved in response to OQDS.
For the remaining PR genotypes (Leccino Lazio, Secolare di Chieuti, and Spina), the putative parents were identified in the autochthonous Apulian germplasm, highlighting the importance of minor neglected accessions in the identification of new sources of tolerance.
Both phylogenetic analysis and PCoA grouped the PR genotypes with the two resistant references (Leccino and FS17). In particular, five of them (Leccino Lazio, Leccio del Corno, Spina, F
9P
4, and F
4P
1) were in the same subcluster of the two references. Similarly, five highly susceptible F
1 genotypes (F
4P
2, F
5P
3, F
5P
5, F
8P
2, and F
8P
5) and cultivars Bianca and Leucocarpa exhibited high genetic similarity with respect to the two susceptible cultivars Ogliarola Salentina and Cellina di Nardò (
Figure 2). These results suggest a possible share of genetic background and indicate the need to further characterize the mechanisms of responses to the pathogen.
5. Conclusions
In the last decade, several multidisciplinary approaches have been adopted to limit the spread of X.f. in Apulia. However, to date, no complete understanding of the pathogenicity of the bacterium, the infection process, and the defense mechanisms of host plants has been achieved. In olive trees, which are of great importance for Apulia’s economy and tradition, only the Leccino and FS17 varieties exhibit high tolerance to the bacterium. In our work, we molecularly characterized and evaluated a collection of 100 local olive genotypes after infection with X. fastidiosa for bacterial load and symptomatology. We identified nine putatively resistant genotypes, of which genotypes Dolce Tonda, F4P1, and F9P1 proved to be of particular interest due to their low bacterial load and the absence of symptoms. The further characterization of these genotypes will allow the identification of new sources of tolerance among the local autochthonous Apulian germplasm and the dissection of the mechanisms involved in plant responses to X.f. infection.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}