
QTL-seq Identifies Pokkali-Derived QTLs and Candidate Genes for Salt Tolerance at Seedling Stage in Rice (Oryza sativa L.)
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions and SKC1 Genotyping
2.2. Salinity Treatment and Phenotypic Evaluation
2.3. Sample Bulking, DNA Isolation and Whole-Genome Sequencing
2.4. QTL-seq Analysis
2.5. Candidate Gene Annotation and Prioritization
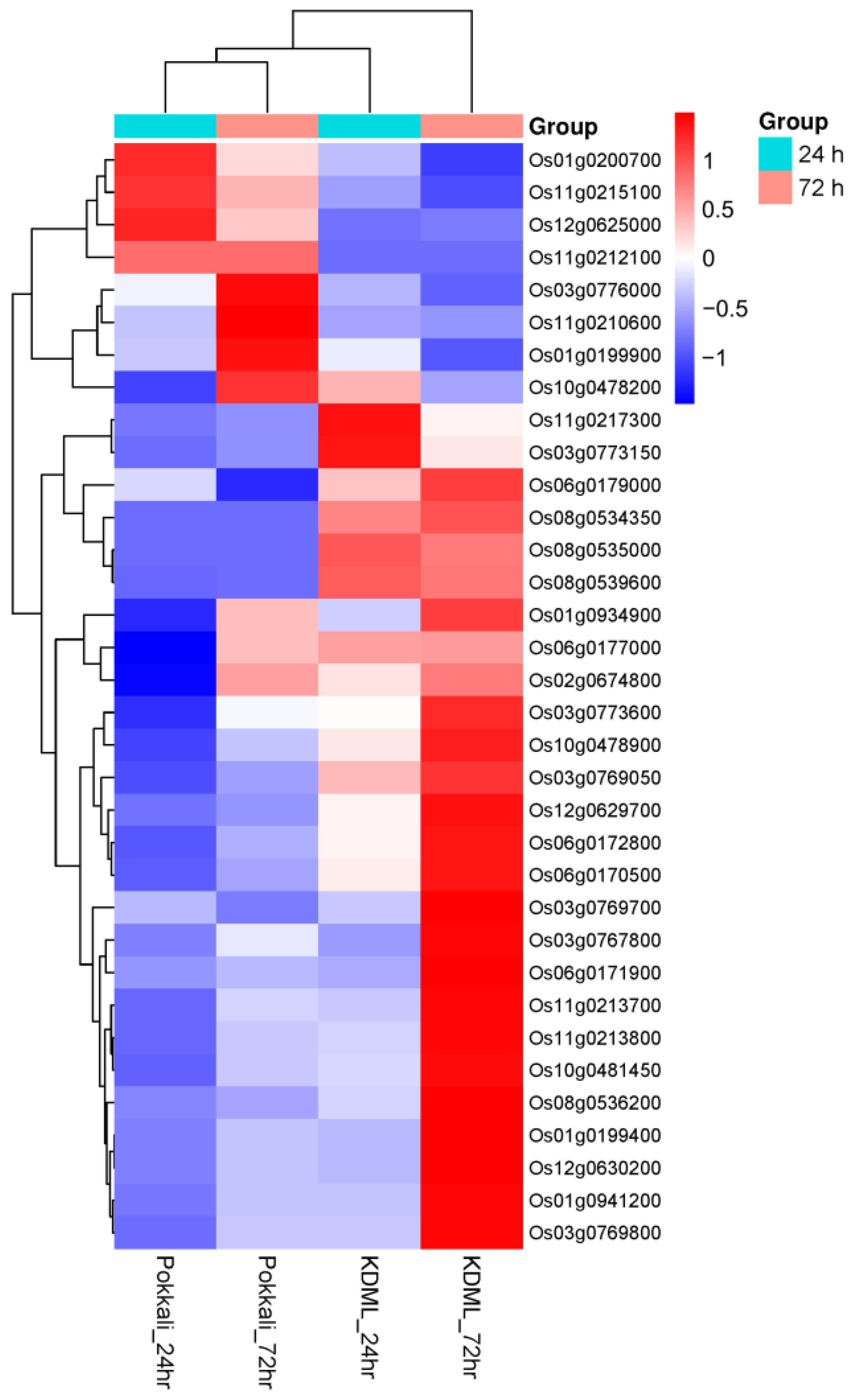
2.6. Differential Gene Expression Analysis of Candidate Genes
3. Results
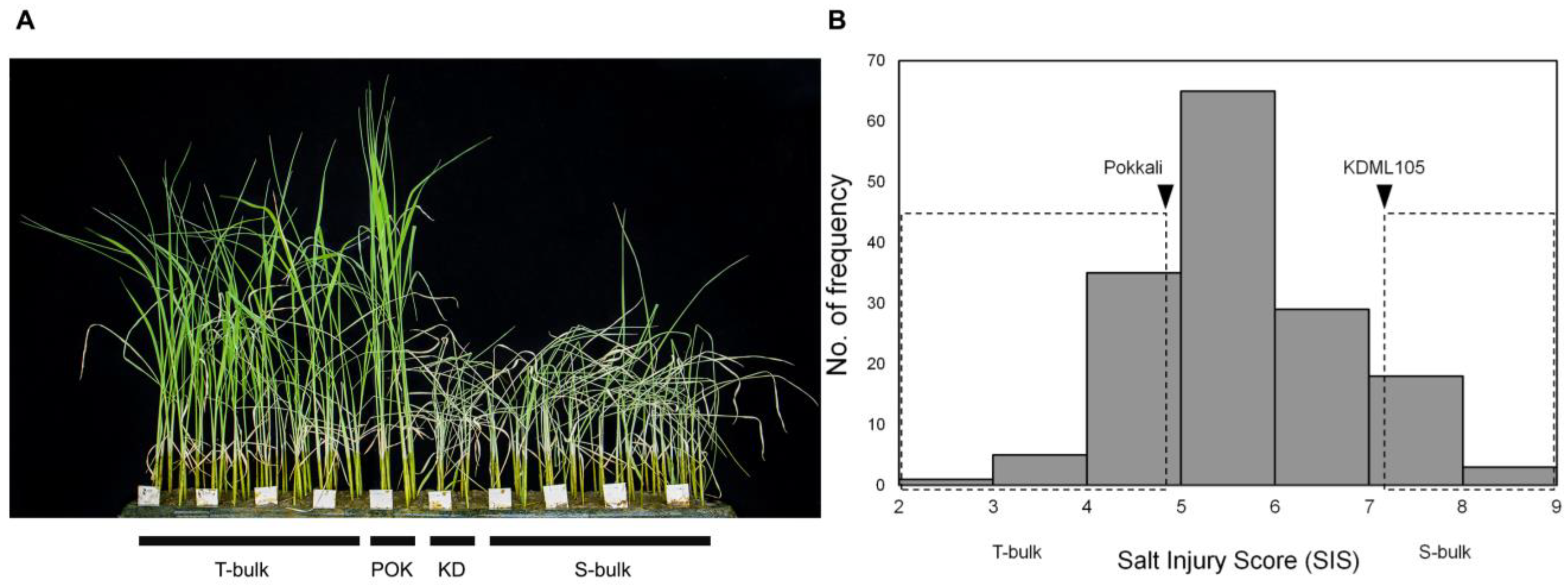
3.1. Phenotypic Screening of Salt Tolerance at Seedling Stage in RILs with Different SKC1 Alleles
3.2. Segregation of the Salinity Tolerance Phenotype in the RIL Population and Construction of Salt-Tolerance (T) and Salt Sensitive (S) Bulks
3.3. Whole-Genome Sequencing of Parents and the Two Bulks of RILs
3.4. Variant Detection and QTL-seq Analysis for Salt Tolerance
3.5. Annotation and Prioritization of Candidate Genes in QTL Regions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khush, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Al Hinai, M.S.; Hafeez, M.B.; Rehman, A.; Wahid, A.; Siddique, K.H.M.; Farooq, M. Regulation of photosynthesis under salt stress and associated tolerance mechanisms. Plant Physiol. Biochem. 2022, 178, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, R.; Sarkar, B.; Jat, H.S.; Sharma, P.C.; Bolan, N.S. Soil salinity under climate change: Challenges for sustainable agriculture and food security. J. Environ. Manag. 2021, 280, 111736. [Google Scholar] [CrossRef] [PubMed]
- Rad, H.E.; Aref, F.; Rezaei, M. Response of rice to different salinity levels during different growth stages. Res. J. Appl. Sci. Eng. Technol. 2012, 4, 3040–3047. [Google Scholar]
- Eynard, A.; Lal, R.; Wiebe, K. Crop Response in Salt-Affected Soils. J. Sustain. Agric. 2005, 27, 5–50. [Google Scholar] [CrossRef]
- Hoang, T.M.L.; Moghaddam, L.; Williams, B.; Khanna, H.; Dale, J.; Mundree, S.G. Development of salinity tolerance in rice by constitutive-overexpression of genes involved in the regulation of programmed cell death. Front. Plant Sci. 2015, 6, 175. [Google Scholar] [CrossRef]
- Gao, J.-P.; Chao, D.-Y.; Lin, H.-X. Understanding abiotic stress tolerance mechanisms: Recent studies on stress response in rice. J. Integr. Plant Biol. 2007, 49, 742–750. [Google Scholar] [CrossRef]
- Shi, Y.; Gao, L.; Wu, Z.; Zhang, X.; Wang, M.; Zhang, C.; Zhang, F.; Zhou, Y.; Li, Z. Genome-wide association study of salt tolerance at the seed germination stage in rice. BMC Plant Biol. 2017, 17, 92. [Google Scholar] [CrossRef] [Green Version]
- Measho, S.; Li, F.; Pellikka, P.; Tian, C.; Hirwa, H.; Xu, N.; Qiao, Y.; Khasanov, S.; Kulmatov, R.; Chen, G. Soil salinity variations and associated implications for agriculture and land resources development using remote sensing datasets in central asia. Remote Sens. 2022, 14, 2501. [Google Scholar] [CrossRef]
- Solis, C.A.; Yong, M.-T.; Zhou, M.; Venkataraman, G.; Shabala, L.; Holford, P.; Shabala, S.; Chen, Z.-H. Evolutionary significance of NHX family and NHX1 in salinity stress adaptation in the genus oryza. Int. J. Mol. Sci. 2022, 23, 2092. [Google Scholar] [CrossRef]
- Hakim, M.A.; Juraimi, A.S.; Hanafi, M.M.; Ismail, M.R.; Rafii, M.Y.; Islam, M.M.; Selamat, A. The effect of salinity on growth, ion accumulation and yield of rice varieties. JAPS J. Anim. Plant Sci. 2014, 24, 874–885. [Google Scholar]
- Bundó, M.; Martín-Cardoso, H.; Pesenti, M.; Gómez-Ariza, J.; Castillo, L.; Frouin, J.; Serrat, X.; Nogués, S.; Courtois, B.; Grenier, C.; et al. Integrative approach for precise genotyping and transcriptomics of salt tolerant introgression rice lines. Front. Plant Sci. 2022, 12, 797141. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Hussain, S.; Ali, B.; Ren, X.; Chen, X.; Li, Q.; Saqib, M.; Ahmad, N. Recent progress in understanding salinity tolerance in plants: Story of Na+/K+ balance and beyond. Plant Physiol. Biochem. 2021, 160, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Jahan, N.; Zhang, Y.; Lv, Y.; Song, M.; Zhao, C.; Hu, H.; Cui, Y.; Wang, Z.; Yang, S.; Zhang, A.; et al. QTL analysis for rice salinity tolerance and fine mapping of a candidate locus qSL7 for shoot length under salt stress. Plant Growth Regul. 2020, 90, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Koyama, M.L.; Flowers, S.A.; Sudhakar, C.; Singh, K.P.; Yeo, A.R. QTL: Their place in engineering tolerance of rice to salinity. J. Exp. Bot. 2000, 51, 99–106. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Jagendorf, A.; Zhu, J.-K. Understanding and Improving Salt Tolerance in Plants. Crop Sci. 2005, 45, 437. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Cheng, J.; Chen, Z.; Huang, J.; Bao, Y.; Wang, J.; Zhang, H. Identification of QTLs with main, epistatic and QTL × environment interaction effects for salt tolerance in rice seedlings under different salinity conditions. Theor. Appl. Genet. 2012, 125, 807–815. [Google Scholar] [CrossRef]
- Rahman, M.A.; Thomson, M.J.; De Ocampo, M.; Egdane, J.A.; Salam, M.A.; Shah-E-Alam, M.; Ismail, A.M. Assessing trait contribution and mapping novel QTL for salinity tolerance using the Bangladeshi rice landrace Capsule. Rice 2019, 12, 63. [Google Scholar] [CrossRef] [Green Version]
- Gregoria, G.B.; Senadhira, D.; Mendoza, R.D. Screening Rice for Salinity Tolerance; Interantional Rice Research Institute: Manila, Philippines, 1997. [Google Scholar]
- Koyama, M.L.; Levesley, A.; Koebner, R.M.; Flowers, T.J.; Yeo, A.R. Quantitative trait loci for component physiological traits determining salt tolerance in rice. Plant Physiol. 2001, 125, 406–422. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.X.; Zhu, M.Z.; Yano, M.; Gao, J.P.; Liang, Z.W.; Su, W.A.; Hu, X.H.; Ren, Z.H.; Chao, D.Y. QTLs for Na+ and K+ uptake of the shoots and roots controlling rice salt tolerance. Theor. Appl. Genet. 2004, 108, 253–260. [Google Scholar] [CrossRef]
- Lee, S.Y.; Ahn, J.H.; Cha, Y.S.; Yun, D.W.; Lee, M.C.; Ko, J.C.; Lee, K.S.; Eun, M.Y. Mapping QTLs related to salinity tolerance of rice at the young seedling stage. Plant Breed. 2007, 126, 43–46. [Google Scholar] [CrossRef]
- Ghomi, K.; Rabiei, B.; Sabouri, H.; Sabouri, A. Mapping QTLs for traits related to salinity tolerance at seedling stage of rice (Oryza sativa L.): An agrigenomics study of an Iranian rice population. OMICS 2013, 17, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.-R.; Fu, C.-Y.; Fan, Z.-L.; Chen, Y.; Chen, W.-F.; Zhang, J.; Jiang, L.-Q.; Lv, S.; Pan, D.-J.; Li, C. Genomic and transcriptomic analysis reveal molecular basis of salinity tolerance in a novel strong salt-tolerant rice landrace Changmaogu. Rice 2019, 12, 99. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J.; de Ocampo, M.; Egdane, J.; Rahman, M.A.; Sajise, A.G.; Adorada, D.L.; Tumimbang-Raiz, E.; Blumwald, E.; Seraj, Z.I.; Singh, R.K.; et al. Characterizing the saltol quantitative trait locus for salinity tolerance in rice. Rice 2010, 3, 148–160. [Google Scholar] [CrossRef] [Green Version]
- Gregorio, G.B.; Islam, M.R.; Vergara, G.V.; Thirumeni, S. Recent advances in rice science to design salinity and other abiotic stress tolerant rice varieties. Sabrao J. Breed Genet. 2013, 45, 31–41. [Google Scholar]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef]
- Singh, V.K.; Khan, A.W.; Jaganathan, D.; Thudi, M.; Roorkiwal, M.; Takagi, H.; Garg, V.; Kumar, V.; Chitikineni, A.; Gaur, P.M.; et al. QTL-seq for rapid identification of candidate genes for 100-seed weight and root/total plant dry weight ratio under rainfed conditions in chickpea. Plant Biotechnol. J. 2016, 14, 2110–2119. [Google Scholar] [CrossRef] [Green Version]
- Pandey, M.K.; Khan, A.W.; Singh, V.K.; Vishwakarma, M.K.; Shasidhar, Y.; Kumar, V.; Garg, V.; Bhat, R.S.; Chitikineni, A.; Janila, P.; et al. QTL-seq approach identified genomic regions and diagnostic markers for rust and late leaf spot resistance in groundnut (Arachis hypogaea L.). Plant Biotechnol. J. 2017, 15, 927–941. [Google Scholar] [CrossRef] [Green Version]
- Clevenger, J.; Chu, Y.; Chavarro, C.; Botton, S.; Culbreath, A.; Isleib, T.G.; Holbrook, C.C.; Ozias-Akins, P. Mapping Late Leaf Spot Resistance in Peanut (Arachis hypogaea) Using QTL-seq Reveals Markers for Marker-Assisted Selection. Front. Plant Sci. 2018, 9, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisano, H.; Sakamoto, K.; Takagi, H.; Terauchi, R.; Sato, K. Exome QTL-seq maps monogenic locus and QTLs in barley. BMC Genom. 2017, 18, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, W.; Guo, N.; Zhang, Y.; Bu, Y.; Zhao, J.; Xing, H. Combining QTL-seq and linkage mapping to fine map a wild soybean allele characteristic of greater plant height. BMC Genom. 2018, 19, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Alam, M.K.; Xie, M.; Yang, L.; Liu, J.; Helal, M.M.U.; Huang, J.; Cheng, X.; Liu, Y.; Tong, C.; et al. Mapping of a major QTL controlling plant height using a high-density genetic map and QTL-seq methods based on whole-genome resequencing in Brassica napus. G3 2021, 11, jkab118. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Lin, T.; Klein, J.; Wang, S.; Qi, J.; Zhou, Q.; Sun, J.; Zhang, Z.; Weng, Y.; Huang, S. QTL-seq identifies an early flowering QTL located near Flowering Locus T in cucumber. Theor. Appl. Genet. 2014, 127, 1491–1499. [Google Scholar] [CrossRef]
- Cao, M.; Li, S.; Deng, Q.; Wang, H.; Yang, R. Identification of a major-effect QTL associated with pre-harvest sprouting in cucumber (Cucumis sativus L.) using the QTL-seq method. BMC Genom. 2021, 22, 249. [Google Scholar] [CrossRef]
- Sheng, C.; Song, S.; Zhou, R.; Li, D.; Gao, Y.; Cui, X.; Tang, X.; Zhang, Y.; Tu, J.; Zhang, X.; et al. QTL-Seq and Transcriptome Analysis Disclose Major QTL and Candidate Genes Controlling Leaf Size in Sesame (Sesamum indicum L.). Front. Plant Sci. 2021, 12, 580846. [Google Scholar] [CrossRef]
- Zhang, C.; Badri Anarjan, M.; Win, K.T.; Begum, S.; Lee, S. QTL-seq analysis of powdery mildew resistance in a Korean cucumber inbred line. Theor. Appl. Genet. 2021, 134, 435–451. [Google Scholar] [CrossRef]
- Ramos, A.; Fu, Y.; Michael, V.; Meru, G. QTL-seq for identification of loci associated with resistance to Phytophthora crown rot in squash. Sci. Rep. 2020, 10, 5326. [Google Scholar] [CrossRef] [Green Version]
- Yaobin, Q.; Peng, C.; Yichen, C.; Yue, F.; Derun, H.; Tingxu, H.; Xianjun, S.; Jiezheng, Y. QTL-Seq Identified a Major QTL for Grain Length and Weight in Rice Using Near Isogenic F 2 Population. Rice Sci. 2018, 25, 121–131. [Google Scholar] [CrossRef]
- Kadambari, G.; Vemireddy, L.R.; Srividhya, A.; Nagireddy, R.; Jena, S.S.; Gandikota, M.; Patil, S.; Veeraghattapu, R.; Deborah, D.A.K.; Reddy, G.E.; et al. QTL-Seq-based genetic analysis identifies a major genomic region governing dwarfness in rice (Oryza sativa L.). Plant Cell Rep. 2018, 37, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Arikit, S.; Wanchana, S.; Khanthong, S.; Saensuk, C.; Thianthavon, T.; Vanavichit, A.; Toojinda, T. QTL-seq identifies cooked grain elongation QTLs near soluble starch synthase and starch branching enzymes in rice (Oryza sativa L.). Sci. Rep. 2019, 9, 8328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, L.; Zheng, H.; Bi, Y.; Yang, L.; Liu, H.; Wang, J.; Sun, J.; Zhao, H.; Li, X.; Li, J.; et al. Identification of a Major QTL and Candidate Gene Analysis of Salt Tolerance at the Bud Burst Stage in Rice (Oryza sativa L.) Using QTL-Seq and RNA-Seq. Rice 2020, 13, 55. [Google Scholar] [CrossRef]
- Nubankoh, P.; Wanchana, S.; Saensuk, C.; Ruanjaichon, V.; Cheabu, S.; Vanavichit, A.; Toojinda, T.; Malumpong, C.; Arikit, S. QTL-seq reveals genomic regions associated with spikelet fertility in response to a high temperature in rice (Oryza sativa L.). Plant Cell Rep. 2020, 39, 149–162. [Google Scholar] [CrossRef]
- Thianthavon, T.; Aesomnuk, W.; Pitaloka, M.K.; Sattayachiti, W.; Sonsom, Y.; Nubankoh, P.; Malichan, S.; Riangwong, K.; Ruanjaichon, V.; Toojinda, T.; et al. Identification and Validation of a QTL for Bacterial Leaf Streak Resistance in Rice (Oryza sativa L.) against Thai Xoc Strains. Genes 2021, 12, 1587. [Google Scholar] [CrossRef] [PubMed]
- Netpakdee, C.; Mathasiripakorn, S.; Sribunrueang, A.; Chankaew, S.; Monkham, T.; Arikit, S.; Sanitchon, J. QTL-Seq Approach Identified Pi63 Conferring Blast Resistance at the Seedling and Tillering Stages of Thai Indigenous Rice Variety “Phaladum”. Agriculture 2022, 12, 1166. [Google Scholar] [CrossRef]
- Pannak, S.; Wanchana, S.; Aesomnuk, W.; Pitaloka, M.K.; Jamboonsri, W.; Siangliw, M.; Meyers, B.C.; Toojinda, T.; Arikit, S. Functional Bph14 from Rathu Heenati promotes resistance to BPH at the early seedling stage of rice (Oryza sativa L.) as revealed by QTL-seq. Theor. Appl. Genet. 2023, 136, 25. [Google Scholar] [CrossRef]
- Riangwong, K.; Aesomnuk, W.; Sonsom, Y.; Siangliw, M.; Unartngam, J.; Toojinda, T.; Wanchana, S.; Arikit, S. QTL-seq Identifies Genomic Regions Associated with Resistance to Dirty Panicle Disease in Rice. Agronomy 2023, 13, 1905. [Google Scholar] [CrossRef]
- Kim, S.-H.; Bhat, P.R.; Cui, X.; Walia, H.; Xu, J.; Wanamaker, S.; Ismail, A.M.; Wilson, C.; Close, T.J. Detection and validation of single feature polymorphisms using RNA expression data from a rice genome array. BMC Plant Biol. 2009, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Chutimanukul, P.; Kositsup, B.; Plaimas, K.; Buaboocha, T.; Siangliw, M.; Toojinda, T.; Comai, L.; Chadchawan, S. Photosynthetic responses and identification of salt tolerance genes in a chromosome segment substitution line of ‘Khao dawk Mali 105’ rice. Environ. Exp. Bot. 2018, 155, 497–508. [Google Scholar] [CrossRef]
- International Rice Research Institute. Standard Evaluation System for Rice; International Rice Research Institute: Los Banios, Philippines, 2013. [Google Scholar]
- Bonilla, P. RFLP and SSLP mapping of salinity tolerance genes in chromosome 1 of rice (Oryza sativa L.) using recombinant inbred lines. Philipp. Agric. Sci. 2002, 85, 68–76. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.; Irizarry, R.A.; Kingsford, C. Salmon provideds fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-H.; Kim, S.-M. Identification of Candidate Genes for Salt Tolerance at the Seedling Stage Using Integrated Genome-Wide Association Study and Transcriptome Analysis in Rice. Plants 2023, 12, 1401. [Google Scholar] [CrossRef]
- De Leon, T.B.; Linscombe, S.; Subudhi, P.K. Identification and validation of QTLs for seedling salinity tolerance in introgression lines of a salt tolerant rice landrace “Pokkali”. PLoS ONE 2017, 12, e0175361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Zhu, Y.; Chen, K.; Shen, C.; Zhao, X.; Shabala, S.; Shabala, L.; Meinke, H.; Venkataraman, G.; Chen, Z.; et al. Identification of new QTL for salt tolerance from rice variety Pokkali. J. Agro. Crop Sci. 2020, 206, 202–213. [Google Scholar] [CrossRef]
- Quan, R.; Wang, J.; Hui, J.; Bai, H.; Lyu, X.; Zhu, Y.; Zhang, H.; Zhang, Z.; Li, S.; Huang, R. Improvement of salt tolerance using wild rice genes. Front. Plant Sci. 2017, 8, 2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bañuelos, M.A.; Garciadeblas, B.; Cubero, B.; Rodríguez-Navarro, A. Inventory and functional characterization of the HAK potassium transporters of rice. Plant Physiol. 2002, 130, 784–795. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Ahn, J.H.; Cha, Y.S.; Yun, D.W.; Lee, M.C.; Ko, J.C.; Lee, K.S.; Eun, M.Y. Mapping of quantitative trait loci for salt tolerance at the seedling stage in rice. In Molecules & Cells; Springer Science & Business Media BV: Berlin/Heidelberg, Germany, 2006; Volume 21. [Google Scholar]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.-X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Dwiyanti, M.S.; Sakaguchi, S.; Koide, Y.; Le, D.V.; Watanabe, T.; Kishima, Y. Identification of a Saltol-Independent Salinity Tolerance Polymorphism in Rice Mekong Delta Landraces and Characterization of a Promising Line, Doc Phung. Rice 2022, 15, 65. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.-H.; Gao, J.-P.; Li, L.-G.; Cai, X.-L.; Huang, W.; Chao, D.-Y.; Zhu, M.-Z.; Wang, Z.-Y.; Luan, S.; Lin, H.-X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.I.; Yamaji, N.; Yamamoto, H.; Okubo, K.; Ueno, H.; Costa, A.; Tanoi, K.; Matsumura, H.; Fujii-Kashino, M.; Horiuchi, T.; et al. OsHKT1;5 mediates Na+ exclusion in the vasculature to protect leaf blades and reproductive tissues from salt toxicity in rice. Plant J. 2017, 91, 657–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, R.; Sazzadur Rahman, M.; Seraj, Z.I.; Thomson, M.J.; Ismail, A.M.; Tumimbang-Raiz, E.; Gregorio, G.B. Investigation of seedling-stage salinity tolerance QTLs using backcross lines derived from Oryza sativa L. Pokkali. Plant Breed. 2011, 130, 430–437. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, X.; Zhai, L.; Shao, K.; Jiang, K.; Shen, C.; Chen, K.; Wang, S.; Wang, Y.; Xu, J. Genetic Bases of the Stomata-Related Traits Revealed by a Genome-Wide Association Analysis in Rice (Oryza sativa L.). Front. Genet. 2020, 11, 611. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, J.; Liu, S.; Jiang, L.; Hu, L. Identification of the metallothionein gene family from cucumber and functional characterization of CsMT4 in Escherichia coli under salinity and osmotic stress. 3 Biotech 2019, 9, 394. [Google Scholar] [CrossRef]
- Romero, L.C.; Domínguez-Solís, J.R.; Gutiérrez-Alcalá, G.; Gotor, C. Salt regulation of O-acetylserine(thiol)lyase in Arabidopsis thaliana and increased tolerance in yeast. Plant Physiol. Biochem. 2001, 39, 643–647. [Google Scholar] [CrossRef]
- De Castro, O.; Innangi, M.; Menale, B.; Carfagna, S. O-acetylserine(thio)lyase (OAS-TL) molecular expression in Pancratium maritimum L. (Amaryllidaceae) under salt stress. Planta 2018, 247, 773–777. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Cleaned Reads (Million) | Cleaned Base (Gb) | % Alignment | Average Depth Coverage (x) |
|---|---|---|---|---|
| Pokkali | 37.44 | 3.74 | 94.21 | 10.38 |
| KDML105 | 41.69 | 4.16 | 94.03 | 11.44 |
| Tolerant bulk (17) | 340.00 | 34.00 | 90.61 | 84 |
| Sensitive bulk (16) | 320.00 | 32.00 | 90.06 | 80 |
| Chr | Length (bp) | No. of SNPs (Depth > 5) | No. of InDels (Depth > 5) | SNP (Depth > 15) | InDels (Depth > 15) |
|---|---|---|---|---|---|
| 1 | 43,270,923 | 338,100 | 74,011 | 26,989 | 8584 |
| 2 | 35,937,250 | 287,762 | 61,423 | 18,595 | 5970 |
| 3 | 36,413,819 | 270,715 | 58,493 | 20,410 | 5849 |
| 4 | 35,502,694 | 235,868 | 45,927 | 24,155 | 6498 |
| 5 | 29,958,434 | 225,149 | 45,596 | 22,070 | 5790 |
| 6 | 31,248,787 | 254,020 | 50,208 | 14,164 | 4220 |
| 7 | 29,697,621 | 238,007 | 45,750 | 11,104 | 3417 |
| 8 | 28,443,022 | 231,655 | 45,177 | 18,061 | 5523 |
| 9 | 23,012,720 | 187,085 | 36,248 | 10,646 | 2913 |
| 10 | 23,207,287 | 205,544 | 37,962 | 8752 | 2734 |
| 11 | 29,021,106 | 252,201 | 45,859 | 15,567 | 4434 |
| 12 | 27,531,856 | 220,737 | 41,052 | 23,409 | 6104 |
| Total | 373,245,519 | 2,946,843 | 587,706 | 213,922 | 62,036 |
| QTL | Chr. | QTL-Region (Mb) | p99 a | p95 b | SNP Index (T-Bulk) | SNP Index (S-Bulk) | Δ(SNP Index) | Reported QTLs/Genes |
|---|---|---|---|---|---|---|---|---|
| qST1.1 | 1 | 5.0–5.50 | 0.34 | 0.26 | 0.28 | 0.86 | 0.58 | |
| qST1.2 | 1 | 41.0–41.50 | 0.34 | 0.26 | 0.10 | 0.65 | 0.55 | qDTS1–2 [57]; qSIS1.41 [58]; qSIS1 [59]; qST1.1 [60]; OsHAK2, OsHAK5, OsHAK6 [61] |
| qST2 | 2 | 27.25–27.75 | 0.33 | 0.25 | 0.17 | 0.74 | 0.57 | qCHL2.20, qRTL2.26 [58] |
| qST3 | 3 | 31.70–32.25 | 0.35 | 0.27 | 0.25 | 0.89 | 0.64 | qCHL3, qSES3 [26]; qNaK3.32 [58] |
| qST6 | 6 | 3.5–4.0 | 0.34 | 0.26 | 0.21 | 0.79 | 0.58 | qSIS6.5, qSHL6.5, qDWT6.5 [58] |
| qST8 | 8 | 26.5–27.0 | 0.35 | 0.27 | 0.40 | 0.99 | 0.59 | qlogSIS8.24, qRTL8.27 [58]; qRSKC8 [59] |
| qST10 | 10 | 17.75–18.25 | 0.34 | 0.26 | 0.27 | 0.87 | 0.60 | |
| qST11 | 11 | 5.75–6.25 | 0.34 | 0.26 | 0.13 | 0.82 | 0.69 | |
| qST12 | 12 | 26.50–27.00 | 0.34 | 0.26 | 0.16 | 0.75 | 0.58 | qSHL12.25 [59] |
| QTL | Chr | Pos | SNP Index T-Bulk | SNP Index S-Bulk | Delta SNP Index | Pokkali | KDML105 | SNP Effect | Strand | Gene ID | Description |
|---|---|---|---|---|---|---|---|---|---|---|---|
| qST1.1 | 1 | 5,288,678 | 0.45 | 0.94 | 0.49 | G | A | GCA > GTA | - | Os01g0197900 | γ-clade RNA-dependent RNA polymerase 3 |
| 1 | 5,313,606 | 0.39 | 0.94 | 0.55 | C | T | GGG > GAG | - | Os01g0198000 | γ-clade RNA-dependent RNA polymerase 4 | |
| qST1.2 | 1 | 41,055,751 | 0.12 | 0.71 | 0.59 | T | C | TAC > CAT | + | Os01g0935300 | Similar to cullin-1 |
| qST6 | 6 | 3,594,804 | 0.24 | 0.85 | 0.62 | T | A | ATT > TTT | - | Os06g0171600 | Membrane insertion protein, OxaA/YidC domain containing protein |
| 6 | 3,687,651 | 0.23 | 0.84 | 0.62 | T | C | ATT > ACT | + | Os06g0173000 | Armadillo-type fold domain containing protein | |
| qST10 | 10 | 18,151,130 | 0.36 | 0.93 | 0.57 | C | T | AGG > AAG | - | Os10g0481400 | Similar to Zinc finger, C3HC4 type family protein |
| qST11 | 11 | 5,909,429 | 0.10 | 0.86 | 0.76 | T | C | AAG > GAG | - | Os11g0213700 | Leucine-rich repeat, typical subtype containing protein |
| 11 | 5,909,645 | 0.15 | 0.90 | 0.76 | G | C | CAG > GAG | - | Os11g0213700 | Leucine-rich repeat, typical subtype containing protein | |
| 11 | 6,024,033 | 0.00 | 0.33 | 0.33 | C | T | GTC > ATC | - | Os11g0215400 | Peptidase aspartic, catalytic domain containing protein. | |
| 11 | 6,063,875 | 0.16 | 0.93 | 0.77 | C | T | GCG > GTG | + | Os11g0216000 | Pyruvate kinase | |
| 11 | 6,147,820 | 0.17 | 0.89 | 0.72 | C | A | GTG > TTG | - | Os11g0218100 | Similar to RNApol24 | |
| qST12 | 12 | 26,580,410 | 0.15 | 0.86 | 0.71 | G | A | GAA > AAA | + | Os12g0622500 | Homologue of the archaeal topoisomerase VIA |
| 12 | 26,602,781 | 0.14 | 0.87 | 0.74 | C | T | GCA > GTA | + | Os12g0622900 | Mov34/MPN/PAD-1 family protein |
| QTL | Chr | Locus ID | 24 h after Salt Stress | 72 h after Salt Stress | Description | ||||
|---|---|---|---|---|---|---|---|---|---|
| PK (TPM) | KD (TPM) | p-Value | PK (TPM) | KD (TPM) | p-Value | ||||
| qST1.1 | 1 | Os01g0199400 | 17.33 | 24.67 | ns * | 26.33 | 67.33 | 0.01 | Alpha/beta hydrolase family protein |
| 1 | Os01g0199900 | 15.33 | 16.33 | ns | 23.00 | 12.33 | 0.00 | Phosphoribosylaminoimidazole carboxylase catalytic subunit | |
| 1 | Os01g0200700 | 4529.00 | 2433.33 | ns | 3203.33 | 1442.00 | 0.01 | Metallothionein-like protein (Tolerance to salinity and heavy metal stresses) | |
| qST1.2 | 1 | Os01g0934900 | 3.67 | 19.33 | 0.01 | 30.00 | 42.33 | ns | Esterase PIR7A |
| 1 | Os01g0941200 | 3.00 | 11.33 | ns | 11.33 | 43.33 | 0.00 | Glucan endo-1,3-beta-glucosidase GII precursor | |
| qST2.1 | 2 | Os02g0674800 | 13.00 | 34.67 | 0.00 | 40.00 | 42.67 | ns | Homeodomain leucine zipper class IV transcriptional factor |
| qST3 | 3 | Os03g0767800 | 128.33 | 138.00 | ns | 171.33 | 292.00 | 0.00 | Cold acclimation protein WCOR413-like protein |
| 3 | Os03g0769050 | 2.00 | 10.00 | 0.01 | 4.67 | 14.33 | 0.01 | Similar to EMB2756 | |
| 3 | Os03g0769700 | 2.33 | 2.67 | ns | 1.33 | 8.00 | 0.01 | Uncharacterised conserved protein | |
| 3 | Os03g0769800 | 16.33 | 21.67 | ns | 21.67 | 39.33 | 0.01 | Homeodomain-leucine zipper protein interfascicular fiberless 1 (Revoluta) | |
| 3 | Os03g0773150 | 99.00 | 226.00 | 0.01 | 110.00 | 156.33 | ns | 40S ribosomal protein S29 (salt tolerance (TO:0006001)) | |
| 3 | Os03g0773600 | 32.00 | 72.33 | 0.00 | 70.33 | 113.00 | 0.00 | Kinesin, motor region domain containing protein | |
| 3 | Os03g0776000 | 56.00 | 51.33 | 0.01 | 74.00 | 45.67 | 0.00 | Glucose-6-phosphate isomerase, cytosolic A | |
| qST6 | 6 | Os06g0170500 | 38.33 | 111.00 | 0.00 | 66.00 | 197.67 | 0.00 | RNA-binding protein-like |
| 6 | Os06g0171900 | 50.67 | 54.67 | ns | 59.00 | 136.00 | 0.00 | WD40 subfamily protein, Salt stress | |
| 6 | Os06g0172800 | 3.00 | 9.67 | ns | 6.33 | 18.00 | 0.01 | Alkaline alpha galactosidase 2 | |
| 6 | Os06g0177000 | 49.33 | 120.33 | 0.00 | 114.33 | 121.00 | ns | Cytochrome b-c1 complex subunit 8 | |
| 6 | Os06g0179000 | 36.00 | 42.33 | ns | 24.00 | 51.33 | 0.00 | Glycoside hydrolase family 79 | |
| qST8 | 8 | Os08g0534350 | 0.00 | 13.33 | 0.00 | 0.00 | 16.00 | 0.00 | Cation cation antiporter |
| 8 | Os08g0535000 | 0.00 | 5.67 | 0.00 | 0.00 | 5.00 | 0.00 | Cation cation antiporter | |
| 8 | Os08g0536200 | 0.00 | 2.33 | ns | 1.00 | 11.67 | 0.01 | trans-membrane plant subgroup domain containing protein | |
| 8 | Os08g0539600 | 0.00 | 74.33 | 0.00 | 0.67 | 68.67 | 0.00 | Ankyrin repeat domain-containing protein 2 | |
| qST10 | 10 | Os10g0478200 | 469.00 | 770.67 | ns | 917.00 | 581.67 | 0.00 | NAD-dependent cytosolic malate dehydrogenase (CMDH) |
| 10 | Os10g0478900 | 21.67 | 44.33 | ns | 35.33 | 65.67 | 0.01 | Multi-organelle localized protein, Control of leaf senescence, Disease resistance, Salt tolerance | |
| 10 | Os10g0481450 | 15.67 | 23.33 | ns | 22.33 | 42.00 | 0.01 | Zinc finger, C3HC4 type family protein | |
| qST11 | 11 | Os11g0210600 | 5.67 | 5.00 | ns | 13.00 | 4.67 | 0.01 | Alcohol dehydrogenase |
| 11 | Os11g0212100 | 2.33 | 0.00 | 0.01 | 2.33 | 0.00 | 0.01 | Similar to NBS-LRR disease resistance protein family-4 | |
| 11 | Os11g0213700 | 2.33 | 6.00 | ns | 6.33 | 17.33 | 0.01 | Leucine-rich repeat, typical subtype containing protein | |
| 11 | Os11g0213800 | 3.67 | 8.67 | ns | 8.33 | 22.67 | 0.00 | NBS-LRR disease resistance protein | |
| 11 | Os11g0215100 | 28.33 | 11.33 | 0.00 | 21.00 | 6.67 | 0.00 | Plant disease resistance response protein family protein | |
| 11 | Os11g0217300 | 8.33 | 22.67 | 0.01 | 9.33 | 14.00 | ns | Mevalonate and galactokinase family protein | |
| qST12 | 12 | Os12g0625000 | 326.67 | 156.00 | 0.00 | 249.67 | 161.00 | ns | O-acetylserine(thiol)lyase, (stress trait (TO:0000164)) |
| 12 | Os12g0629700 | 0.67 | 9.00 | 0.00 | 2.67 | 21.00 | 0.00 | Thaumatin-like protein precursor | |
| 12 | Os12g0630200 | 2.67 | 13.00 | 0.01 | 15.00 | 72.67 | 0.00 | Thaumatin, pathogenesis-related family protein | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Songtoasesakul, D.; Aesomnuk, W.; Pannak, S.; Siangliw, J.L.; Siangliw, M.; Toojinda, T.; Wanchana, S.; Arikit, S. QTL-seq Identifies Pokkali-Derived QTLs and Candidate Genes for Salt Tolerance at Seedling Stage in Rice (Oryza sativa L.). Agriculture 2023, 13, 1596. https://doi.org/10.3390/agriculture13081596
Songtoasesakul D, Aesomnuk W, Pannak S, Siangliw JL, Siangliw M, Toojinda T, Wanchana S, Arikit S. QTL-seq Identifies Pokkali-Derived QTLs and Candidate Genes for Salt Tolerance at Seedling Stage in Rice (Oryza sativa L.). Agriculture. 2023; 13(8):1596. https://doi.org/10.3390/agriculture13081596
Chicago/Turabian StyleSongtoasesakul, Decha, Wanchana Aesomnuk, Sarinthip Pannak, Jonaliza Lanceras Siangliw, Meechai Siangliw, Theerayut Toojinda, Samart Wanchana, and Siwaret Arikit. 2023. "QTL-seq Identifies Pokkali-Derived QTLs and Candidate Genes for Salt Tolerance at Seedling Stage in Rice (Oryza sativa L.)" Agriculture 13, no. 8: 1596. https://doi.org/10.3390/agriculture13081596