1. Introduction
Garlic (
Allium sativum L.) is a bulbous perennial crop cultivated in different temperate and subtropical climates all over the world [
1,
2]. It belongs to the genus
Allium, which includes almost 1008 species distributed in 15 subgenera and more than 70 sections [
3,
4]. Within the Alliaceae family, garlic is the most widely consumed bulb after onion [
5]. The total world production for garlic reached 28,204,854.32 metric tons in 2021, with China being the largest producer, accounting for nearly 73% of total world production at 20,513,385.83 tons. In Greece, garlic is cultivated in approximately 780 ha with a total production of 8230 tons (
http://fao.org/faostat/, accessed on 28 April 2023).
A. sativum was domesticated long ago; with references to its use as a vegetable, condiment, and medicinal plant dating back to ancient Egypt and India 5000 years ago, garlic is one of the oldest known crops [
4]. Central Asia is considered as garlic’s primary center of origin, from where it spread to Southeastern Asia, Egypt, and the Mediterranean region [
6,
7]. Regarding garlic’s ancestry, the most recent theory is that
A. longicuspis Regel is either the closest wild relative or the wild progenitor of cultivated garlic, although there is some debate over it [
6,
8].
Throughout history, garlic has been recognized as a crop of great importance in many aspects, especially those related to its significant medicinal properties [
9,
10]. Garlic consumption confers many health benefits, as it is a good source of vitamins, metals, saponins, flavonoids, organic acids, and various organosulfur compounds originating from alliin that is metabolized to allicin by the enzyme alliinase. Allicin and other sulfoxides may encounter many transformations resulting in various organosulfur volatiles, responsible for garlic’s medicinal properties, as well as its flavor and aroma [
9,
11,
12]. Garlic has been proven beneficial for the prevention and treatment of cardiovascular and other metabolic diseases, and it stimulates the immune system, while it also possesses antifungal, antibacterial, antigenotoxic, antiviral, and antispasmodic properties [
10,
12,
13].
Garlic is a diploid (2n = 2x = 16) sterile species, which is primarily propagated asexually via its bulblets (e.g., cloves) due to obligate apomixis [
14,
15]. Despite the existence of a few fertile wild genotypes, most of the germplasm collections worldwide comprise nonflowering plants and thus clonal selection is the method of choice in garlic breeding [
4,
5].
Garlic exhibits wide biodiversity, environmental adaptation capacity, and phenotypic plasticity, resulting in the establishment of diverse ecotypes in various areas of cultivation [
16,
17,
18]. This variation in domesticated garlic is primarily due to spontaneous mutations arising throughout the history of large-scale cultivation [
19,
20,
21], but also due to variability inherited from its wild ancestor [
14]. Clonal propagation is thought to have caused a genetic bottleneck, accounting for the narrow genetic base observed in this crop. On the other hand, clonal lineages within this species exhibit high levels of diversity in several phenotypic characteristics and valuable agronomic traits, such as stress and drought tolerance [
14,
19,
22,
23].
A wealth of variability, primarily related to the crop’s morphological and genetic characteristics, chemical composition, and nutritional value, exists in germplasm accessions, particularly local landraces that are still cultivated by farmers either for trade or for their own consumption [
24,
25]. Since variation is essential for breeding, genetic research on germplasm accessions and landraces is necessary to understand the nature of variation and to ensure their effective use for breeding new varieties [
18,
23,
26,
27].
So far, variation assessment of garlic germplasm collections has revealed a significant amount of diversity in morphological, agronomic, and chemical traits, e.g., [
2,
26,
28,
29,
30,
31,
32]. Recently, garlic genotypes from different areas of Greece were evaluated for their chemical composition, quality, and bulb morphology [
33]. In a more comprehensive survey, Polyzos et al. [
34] examined the phenotypic diversity of thirty-four (34) Greek garlic accessions, using morphological descriptors and agronomic traits, while Avgeri et al. [
11] determined their levels of organosulfur compounds, pyruvate, total sugars, total phenolics, and antioxidant properties. All these studies revealed remarkable diversity, demonstrating the need for the valorization of Greek garlic genotypes and their importance for conservation and breeding.
Traditionally, morphological characters are used to assess garlic diversity. However, since morphological characters are affected by environmental conditions, they may not mirror the available genetic diversity [
26,
35]. Molecular markers are considered a dynamic tool for the identification, characterization, and assessment of genetic diversity, since they are less affected by age, the physiological condition of samples, and the environment [
1,
36]. So far, numerous types of markers have been used in garlic for germplasm identification, crop improvement, and genetic diversity evaluation, such as amplified fragment length polymorphisms (AFLPs) [
16,
37,
38,
39], random amplified polymorphic DNA (RAPD) [
14,
40,
41], sequence-related amplified polymorphisms (SRAPs) [
5,
42], insertions–deletions (InDels) [
39], inter-simple sequence repeats (ISSRs) [
1,
18,
22], and simple sequence repeats (SSRs) [
2,
15,
17,
35,
43,
44,
45]. Among these types of markers, SSRs became very popular since they are typically codominant, reproducible, highly polymorphic, and cross-species transferable [
17,
26], while ISSRs are broadly used in genetic studies as they are informative, reliable, and efficient in laboratory usage [
18,
28].
In this study, we evaluated the genetic diversity of a Greek garlic collection comprising 27 local varieties, also known as landraces, using SSR and ISSR markers. Moreover, in an attempt made to identify potential markers associated with variations in the chemical composition and the morphological characters of the bulb, we investigated possible significant associations between the SSR and the ISSR loci from this study and the characters recorded by Avgeri et al. [
11] and Polyzos et al. [
34] for the same cultivars. Our research, combined with the studies of Petropoulos et al. [
33], Polyzos et al. [
34], and Avgeri et al. [
11], provides for the first time a comprehensive overview on the genetic and phenotypic variation, the chemical composition, and the nutritional value of Greek garlic landraces. Our results will further contribute to the effective management and conservation of garlic genetic resources, also highlighting their value for potential utilization in breeding programs.
3. Results
3.1. Genetic Relationships Based on the SSR Markers
All the SSR markers tested were polymorphic in all the accessions studied. PCR amplification revealed four (Asa24, GB-AS-076, GB-AS-102) to nine (Asa08) alleles for each SSR locus, totaling 26 different alleles, with an average (Na) of 5.2 alleles per locus. Some of the alleles, e.g., Asa24-141, can be used as diagnostic for identification purposes. The average number of effective alleles (Ne) was 2.460, ranging from 1.157 (GB-AS-102) to 3.609 (Asa10), while the Shannon’s information index (I) ranged from 0.332 (GB-AS-102) to 1.548 (Asa08) with an average of 1.013. The estimated gene flow (Nm) ranged from 0.038 for GB-AS-102 to 0.936 for GB-AS-076 (mean 0.251), while the average PIC number was 0.471, ranging from 0.133 (GB-AS-102) to 0.683 (Asa10). All SSR markers except GB-AS-102 were informative (PIC ≥ 0.4).
Ranges for observed (Obs. He) and expected heterozygosity (Exp. He) were 0.036–0.893 and 0.136–0.726, respectively. Finally, for all the SSR markers, the Fis index was negative, while Fst ranged from 0.211 (GB-AS-076) to 0.868 (GB-AS-102) with a mean value of 0.5 (
Table 4).
Pairwise genetic distance [
61] values (D) among the garlic accessions ranged from 0 to 2.538 (
Table S1). Zero genetic distance values were estimated in several pairwise combinations, implying genetic identity among garlic accessions, at least for the loci studied. 17 out of 27 accessions had identical SSR genotype with at least one other accession. The genetically most distant accession was AS29 (
A. schoenoprasum) (Dmax = 2.538 from both AS04 and AS30 accessions); however, when we examined only
A. sativum accessions, the highest value was calculated between both AS04 and AS08 with AS06 (D = 2.079).
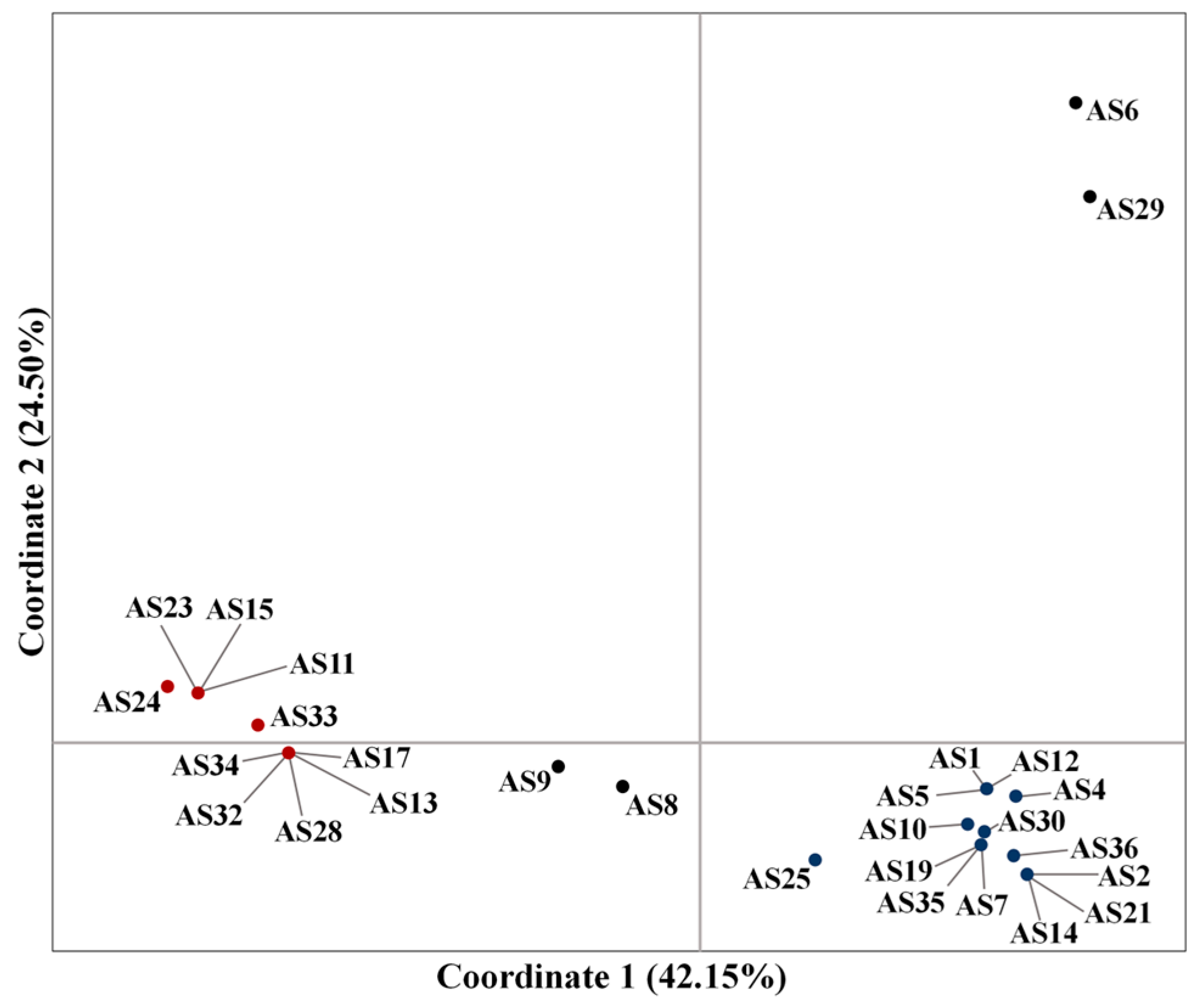
For the clarification of the genetic relationships among the 27 garlic genotypes studied, a principal coordinate analysis (PCoA) was performed (
Figure 1), including the
A. schoenoprasum accession (AS29). In the principal coordinate analysis (PCoA), the percentage of variation explained by the first three axes was 83.57%. The first axis explained 42.15%, while the second and third axes explained 24.50% and 16.92%, respectively. There was a clear separation of the AS6 and AS29 accessions from the rest. All other genotypes were assigned into two distinct groups. The first group comprised accessions AS1, AS2, AS4, AS5, AS7, AS10, AS12, AS14, AS19, AS21, AS25, AS30, AS35, and AS36, while the second comprised accessions AS11, AS13, AS15, AS17, AS23, AS24, AS28, AS32, AS33, and AS34. Finally, the accessions AS8 and AS9 were placed in between the two groups, with AS8 being closer to the first group of accessions and AS9 closer to the second group of accessions.
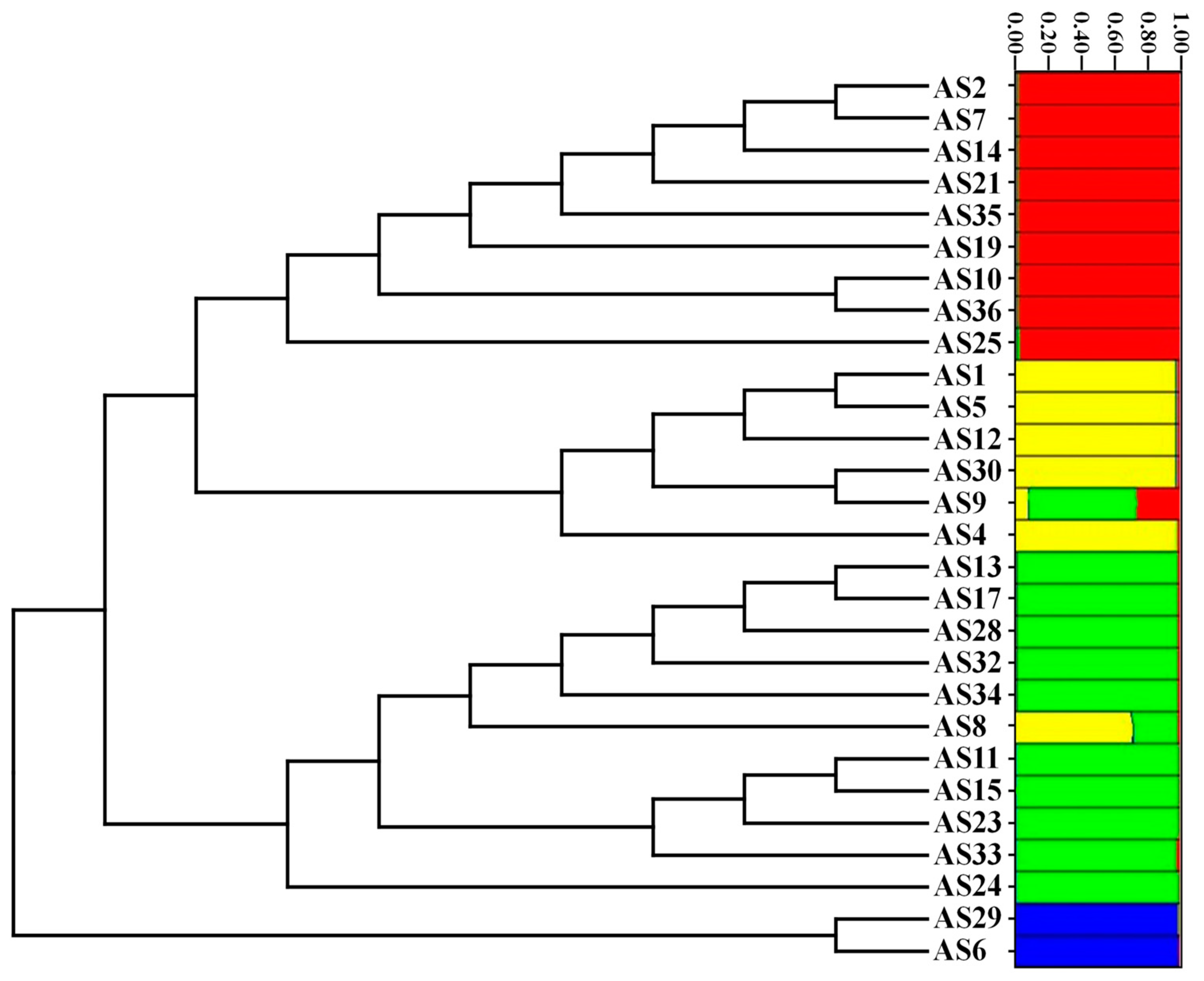
The UPGMA similarity dendrogram, based on Nei and Li/Dice similarity index, showed that the 27 accessions, and the outgroup (
A. schoenoprasum), were classified into three different clusters (
Figure 2), and for the most part, it is congruent with PCoA. The first cluster comprised accessions AS6 and AS29; the second cluster comprised accessions AS8, AS11, AS13, AS15, AS17, AS23, AS24, AS28, AS32, AS33, and AS34; while the third one comprised accessions AS1, AS2, AS4, AS5, AS7, AS9, AS10, AS12, AS14, AS19, AS21, AS25, AS30, AS35, and AS36.
The maximum peak was calculated using STRUCTURE HARVESTER v0.6.94 at K = 4, with CLUMPAK (Tel Aviv University, Tel Aviv, Israel) supporting the peak of 10 out of 10 STRUCTURE runs. This suggests the existence of four different genetic clusters for the garlic accessions under study. The accessions AS6 and AS29 formed one cluster supporting their clear distinction from the rest. The second cluster contained AS2, AS7, AS10, AS14, AS19, AS21, AS25, AS35, and AS36, the third AS11, AS13, AS15, AS17, AS23, AS24, AS28, AS32, AS33, and AS34, while the accessions AS1, AS4, AS5, AS12, and AS30 formed the fourth cluster. Finally, the AS8 and AS9 accessions were distributed in more than one cluster.
Analysis of molecular variance (AMOVA) revealed that 34% of the overall molecular diversity was attributed to variations among accessions, while the rest (66%) was within them (
Table 5).
3.2. Genetic Relationships Based on ISSR Analysis
Six ISSR primers were used to screen the 27 garlic accessions and the A. schoenoprasum and to estimate the genetic diversity among them. A total of 84 clear and stable polymorphic bands were generated with an average of 14 bands for each primer; all bands were found to be polymorphic among the garlic landraces studied, whereas some of those bands can be used as diagnostic to discriminate among the accessions studied. Among the primers utilized, UBC-811 produced the most abundant number of bands (19), while UBC-880 exhibited the lowest band count (7).
The PIC values for the six ISSR primers ranged from 0.370 to 0.500 with a mean value of 0.465. The binary matrix generated from the ISSR banding profiles corresponded to eighty-four different loci. All calculated Nei’s genetic distances (GDs) were used to perform a PCoA. From the PcoA, the first three axes explained 67.87% of the total variation (the first, second and third axes explained 31.92%, 21.52%, and 14.44%, respectively). These values were due to the low levels of polymorphism exhibited by the specific ISSR markers. Consequently, the garlic accessions were scattered throughout the plot, not forming any visible groups, as expected from the coordinate values (
Figure S1), reducing the resolution power of those markers.
The UPGMA similarity dendrogram, based on Nei and Li/Dice similarity index, classified the 27 accessions, and the
A. schoenoprasum accession (outgroup), into six clusters (
Figure S2). The first cluster comprised the
A. schoenoprasum accession (AS29), the second cluster comprised the accession AS6, and the third one comprised the accession AS36. The fourth cluster comprised the accession AS11 and the fifth one comprised the accessions AS2, AS9, AS28, AS32, AS33, and AS34.
Lastly, the sixth cluster was further divided into two subclusters. The first subcluster comprised accessions AS19, AS21, AS23, AS24, AS25, and AS35, while the second, accessions AS1, AS4, AS5, AS7, AS8, AS10, AS12, AS13, AS14, AS15, AS17, and AS30. Although the UPGMA dendrogram had some similarities with the PCoA, they were not totally in agreement.
The AMOVA revealed 79% of the total molecular diversity within and 21% among the Greek garlic landraces (data not presented).
Based on the Mantel test, a relatively low correlation was observed between the SSR and the ISSR data (r = 0.474, p < 0.01, 999 permutations).
3.3. Association of SSR Loci with Chemical Compounds and Bulb Morphological Traits
Fisher’s exact tests revealed many SSR and ISSR loci being significantly associated with the chemical compounds [
11] as well as the morphological traits of the bulb [
35] for the same accessions (
Tables S2 and S3).
The statistically significant associations were further evaluated using logistic regression analysis. The statistically significant correlations (
p < 0.05), which were identified using Fisher’s exact test and multinomial logistic regression analysis, between the SSR and ISSR loci, the chemical compounds, and the morphological traits of the bulb are presented in
Table 6.
4. Discussion
Although most garlic varieties are asexually propagated, significant morphological differences exist within and among them, mainly due to varying environmental conditions, thus creating obstacles in their identification and conservation [
17,
35]. Assessing the levels of genetic variation existing within garlic accessions is beneficial for phenotypic identification and core germplasm construction. Moreover, the evaluation of genetic diversity and the estimation of the degree of relatedness between accessions are critical steps for garlic selection and for breeding purposes [
62].
In this study, using SSR and ISSR markers, we aimed to molecularly characterize and assess the genetic diversity and structure of a garlic collection comprising 27 accessions collected from different regions of Greece. These accessions have been previously investigated in terms of phenotypic diversity as well as chemical content by Polyzos et al. [
34] and Avgeri et al. [
11], respectively.
SSRs are considered ideal DNA markers due to their stability and reproducibility [
26]; thus, they have been widely used to assess genetic diversity in local garlic collections [
2,
15,
17,
26,
35,
43,
44,
45]. The SSR markers used in this study revealed a mean PIC value of 0.471, which is lower than the PIC values reported by Cuhna et al. [
43,
44], which were 0.518 and 0.500, respectively. The PIC value revealed in this study is also lower than the mean PICs of 0.72 and 0.60 reported by Chen et al. [
26] and Ipek et al. [
63], but higher than those reported by Li et al. [
17] (0.37) and Barboza et al. [
31] (0.38) in Korean and Argentinian accessions, respectively.
Compared to SSRs, ISSR markers revealed a similar mean PIC value (0.47), which was higher than the PIC value (0.19) reported by Kıraç et al. [
22] in Turkish accessions, but significantly lower than those reported by Chen et al. [
27] (0.72) and Rakesh Sharma et al. [
27] (0.65) for Chinese and Indian collections, respectively.
Differences in the mean PIC values observed among various genetic diversity studies are attributed to several factors, such as the number and type of SSR markers, e.g., EST-SSRs or genomic, their distribution and abundance in the genome, the number and geographic location of the accessions, the population size, and the genetic variation existing in the germplasm resources analyzed [
17,
31].
Other estimated parameters, such as observed (Obs. He) and expected heterozygosity (Exp. He), Shannon’s Information index (I), number of alleles per locus (Na), and effective number of alleles (Ne) (
Table 4), reflected comparative levels of genetic variation between ours and other previously characterized garlic germplasm collections, e.g., [
15,
31,
44]. It should also be noted that the values of Obs. Ho > Exp. He, as observed for the SSR loci studies, except for Asa10 and GB-AS-102, and the negative values of fixation index (Fis) indicate an excess of heterozygotes and the absence of null alleles in the garlic collection, as expected in vegetatively propagated species [
44].
DNA markers can be effectively used to resolve issues in the nomenclature of accessions and reveal possible redundancies in a germplasm collection. Identification of potential duplicates is not only of paramount importance to identify genuine variability for breeding purposes, but also helps to reduce the maintenance costs and to secure the authenticity of the accessions [
17,
64]. In our study, zero genetic distance values were observed in various pairwise comparisons, indicating genetically identical accessions for the loci analyzed. Furthermore, 63% of the accessions (17 out of 27) shared the same SSR genotype with at least one other accession and only one genotype (AS6) had a unique genotype revealing some private alleles. This fact may reflect a high level of redundancy and existence of duplicates in the garlic collection studied, although the same observation appeared less frequently in the ISSR screening. High levels of redundancy have also been observed in other garlic collections with potential duplicates reaching up to 50% of the collection [
31,
35,
44,
63].
The genetic similarity among accessions revealed in our study does not correlate with the levels of phenotypic and chemical variation observed for the same collection by Polyzos et al. [
34] and Avgeri et al. [
11]. Most of the accessions analyzed in this study are phenotypically distinct with respect to morphological, compositional (e.g., considering the content of bulb phytochemicals), and agronomic traits. In garlic, clonal propagation accounts for the narrow genetic base observed in this crop. As a result, meiotic recombination is missing, and gene exchange and genetic introgression are at low frequency [
19,
22,
23]. Unlike other seed crops, the bulbs, seedlings, and stems of garlic are targeted harvesting organs, harvested before flowering and seeding, leading also to the narrow genetic base. On the other hand, garlic exhibits wide phenotypic diversity in different ecological areas of cultivation due to naturally occurring mutations and also soil type, environmental parameters, and cultural techniques, which eventually lead to a great number of cultivars [
21]. This discrepancy between phenotypic and genetic data may arise because either the farmers tend to exchange their garlic varieties and thus generate confusion in garlic nomenclature through naming the same local variety/landrace with different names or vice versa [
17,
39], or it could be due to epigenetic phenomena which also cause significant phenotypic variation in garlic lines cultivated in the same region for an extended period [
15,
65,
66].
The distribution of genetic variation in the garlic accessions based on the SSR markers, as examined via AMOVA analysis, indicated that 34% of the total molecular diversity was attributable to differences among accessions, while the rest (66%), within them. A similar trend was also observed for the ISSR data. These findings agree with the results of Zhao et al. [
67], Cuhna et al. [
44], Kumar et al. [
45], Poljuha et al. [
15], and Li et al. [
17], who also detected higher levels of variation within the accessions rather than among them. Nevertheless, this pattern contradicts the hypothesis of genetic erosion and possibly indicates the presence of genetic structures due to ancestral sexual reproduction, followed by independent domestication history [
15,
17]. The presence of higher levels of molecular diversity within garlic accessions rather than among them is a result not only found in our study, but also in many others, and is usually attributed to the lack of sexual propagation but also to human-mediated gene flow, which leads to a substantial amount of variation that can be attributed to differences within the accessions rather than among them [
65]. Generally, landraces typically exhibit varying degrees of variability, which can be assessed through measuring different traits. Within a landrace, some traits can express high variability and therefore cannot be used as discrimination characters. On the other hand, several traits within a landrace are uniform and can be used for identification [
68]. Moreover, since an Fst value greater than 0.15 is considered significant in differentiating populations [
69], the value observed in our study (0.51) implies a considerable degree of differentiation among the accessions studied, especially when combined with low levels of gene flow.
Assessment of the genetic diversity and population structure of germplasm collections facilitates crop breeding, allowing the more efficient use of genetic resources and management of genetic variation. In our study, in the SSR data analysis, both STRUCTURE results (optimal K = 4) including
A. schoenoprasum coincided with the UPGMA similarity dendrogram, based on the Nei and Li/Dice similarity index, indicating convergent results to the garlic accessions’ genetic relatedness. Moreover, the PCoA was for the most part concordant to both STRUCTURE and clustering analyses for the SSR data. The number of subpopulations (K) revealed using STRUCTURE was also in agreement with that found by other studies of garlic germplasm collections [
31,
39].
In the present study, we did not find any association between genetic clustering and the geographical origin/collection site of the garlic accessions. The same has also been reported by Garcia-Lampasona et al. [
37,
70], Volk et al. [
16], Morales et al. [
71], and Barboza et al. [
31], whereas other studies reported a moderate level of correlation between genetic clustering based on SSR markers and geographical origin [
35,
45,
67,
72]. This lack of correlation may be due to various factors, such as the exchange of genetic materials without keeping track of their geographical origin information, the introduction of cultivars with the same origin to various regions using different names, the accumulation of different somatic mutations due to different selective pressures when the same clone is cultivated for a long time in different environments, etc. [
31,
39,
44].
In an attempt made to unpack the degree of association and reveal potential molecular markers related to important traits, we examined if any association exists between the molecular data and the chemical composition [
11], as well as between the molecular data and the morphological traits of the bulb [
34], for the same accessions. For this reason, Fisher’s exact tests were initially employed and then we conducted for the genetic loci exhibiting significant associations (
p < 0.05), a multinomial logistic regression analysis to further explore their relationships with the specific traits studied.
The multinomial logistic regression is suitable for MAS breeding programs and GWAS studies [
73,
74]; thus, the markers which present significant correlation with important traits, either chemical or morphological, are potentially good candidates to be utilized for the selection of promising progeny in breeding programs. We found significant associations between specific loci and certain chemical compounds (i.e., allyl-prop-1-enyl disulfide, methyl allyl disulfide, and pyruvate), as well as the morphological characters of the bulb (i.e., BDC, BPCTB, BPRD, SBB, SMB). For example, a positive association was revealed between Asa10-Locus 4 and pyruvate. Pyruvate has been utilized to indirectly assess the presence of allicin in fresh raw garlic, since it is produced as a by-product during the conversion of alliin into allicin [
75]. Additionally, previous studies have shown a positive correlation between pyruvate levels and both flavor intensity (pungency) [
76] and antiplatelet activity [
77]. Avgeri et al. [
11] have observed significant variability in pyruvate levels, with a nearly 15-fold difference between the genotypes possessing the lowest and highest concentrations. This suggests that molecular markers could foster the identification of mild and pungent garlic genotypes, as well as genotypes with high functional properties, to facilitate their selection in future breeding programs.

 ,
, 


{kind=link}
{kind=link}