

Rhizobia: A Promising Source of Plant Growth-Promoting Molecules and Their Non-Legume Interactions: Examining Applications and Mechanisms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Plant Growth-Promoting Rhizobia Diversity and Ecology
3. Characteristics of Rhizobia
4. Role of Rhizobia as PGPR and Mechanism of Action
4.1. Direct Mechanism
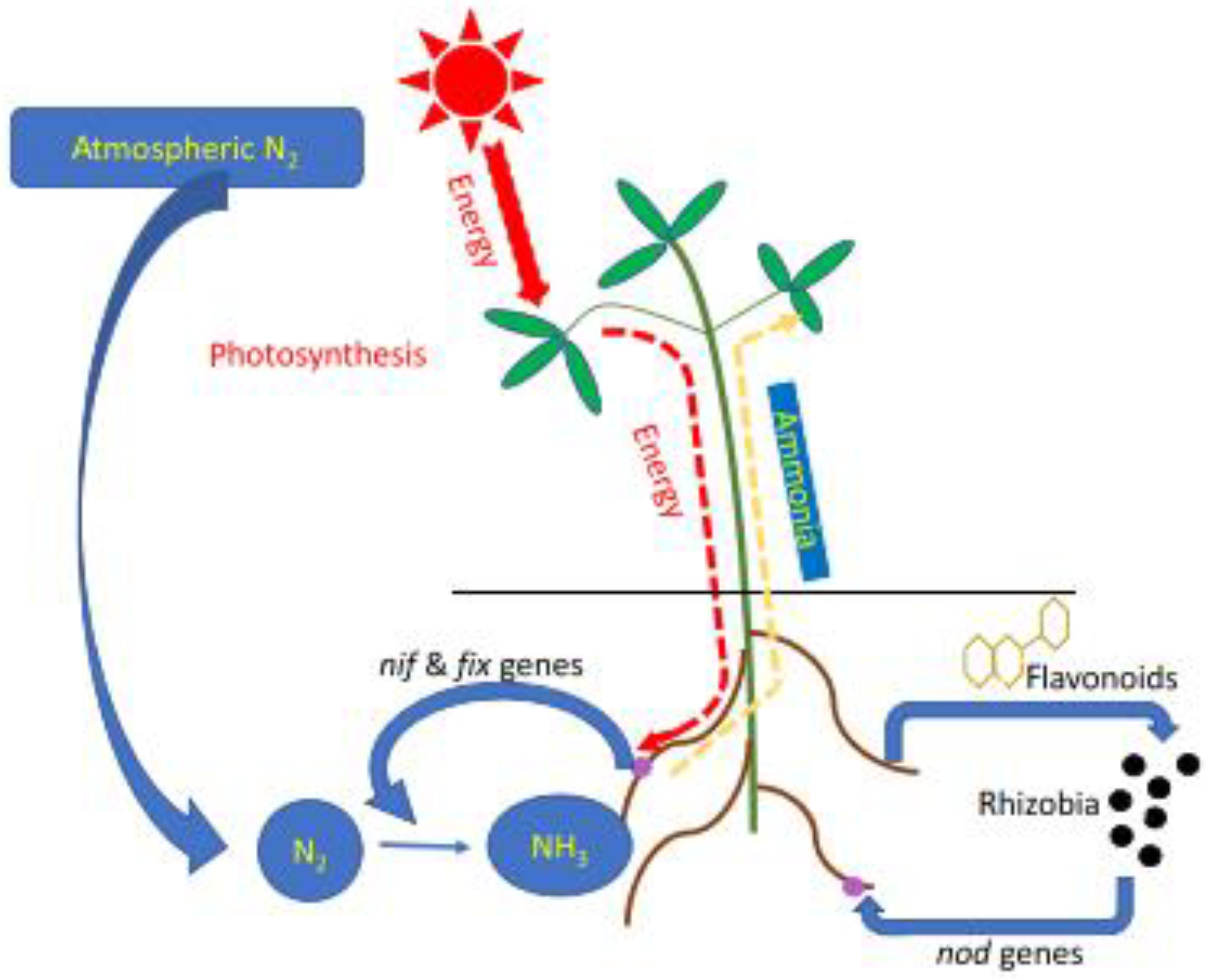
4.1.1. N2 Fixation
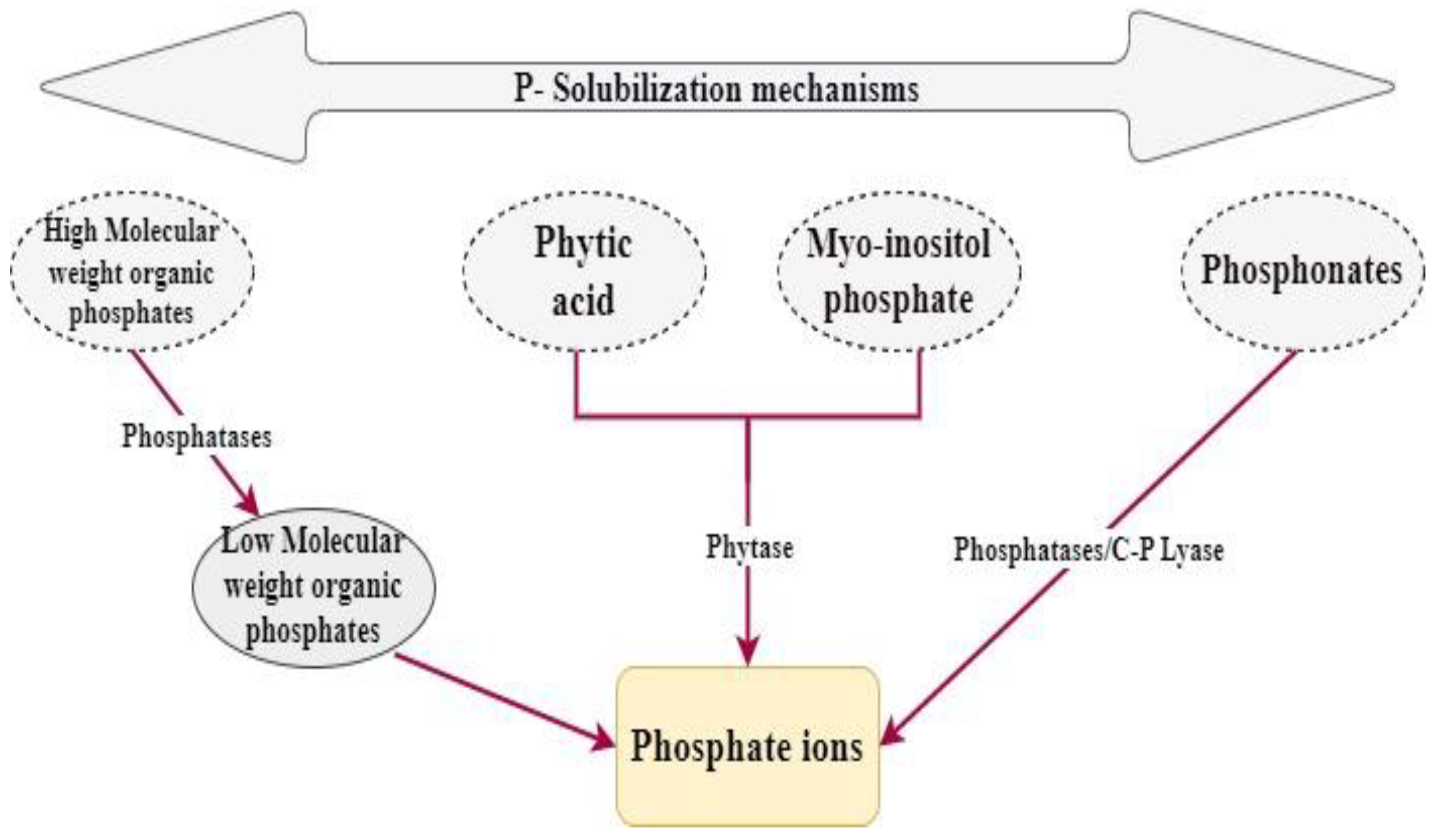
4.1.2. Phosphate Solubilization
- Inorganic P solubilization mechanisms
- Organic Acid ProductionThe production of organic acid is considered as the initial mechanism to be used in order to solubilize inorganic phosphorus [64]. This phenomenon depends mostly on the soil’s pH.Phosphate Solubilizing Microbes (PSM) produce, during their growth, some organic acids that have the potential to decrease the soil’s pH. This acidification enables the solubilization of rock phosphate [64].It has been established that the acids produced by PSM are mainly glycolic (monocarbocyclic hydroxy acids), 2-keto gluconic (monocarboxylic), acetic acids, malic (dicarboxylic hydroxy acids), oxalic acid, citric acid, and succinic acid (dicarboxylic acid). However, in the midst of all these acids, gluconic acid has been found to be the lead acid to be used in the P solubilization mechanism [44,64].
- Inorganic Acid ProductionInorganic acids do not appear to be as effective as organic acids for the solubilization of P. Nitrifying and sulfur-oxidizing bacteria generate inorganic acids during the oxidation of nitrogenous or inorganic sulfur compounds. These inorganic acids then interact with insoluble phosphate compounds, transforming them into soluble variants [64].
- ChelationFulvic, humic, and 2-keto gluconic acids are some known acids that play the role of chelators of substances, such as aluminum, calcium, and iron cations, which facilitate the inorganic phosphorus solubilization. These acids are liberated all along the processes of plant debris degradation by microorganisms [64].Besides the previously mentioned inorganic P solubilization mechanisms, there are other mechanisms, such as mineral P solubilization through proton (H+) extrusion. This process effectively lowers the pH of the environment without requiring the release of acids [63]. Furthermore, microorganisms that produce exopolysaccharides have the ability to form complexes with metals, resulting in the solubilization of metal phosphates [64].
- Organic P Solubilization Mechanisms
4.1.3. Phytohormones Production/Regulation (Plant Growth Regulators)
- Auxins
- Cytokinins
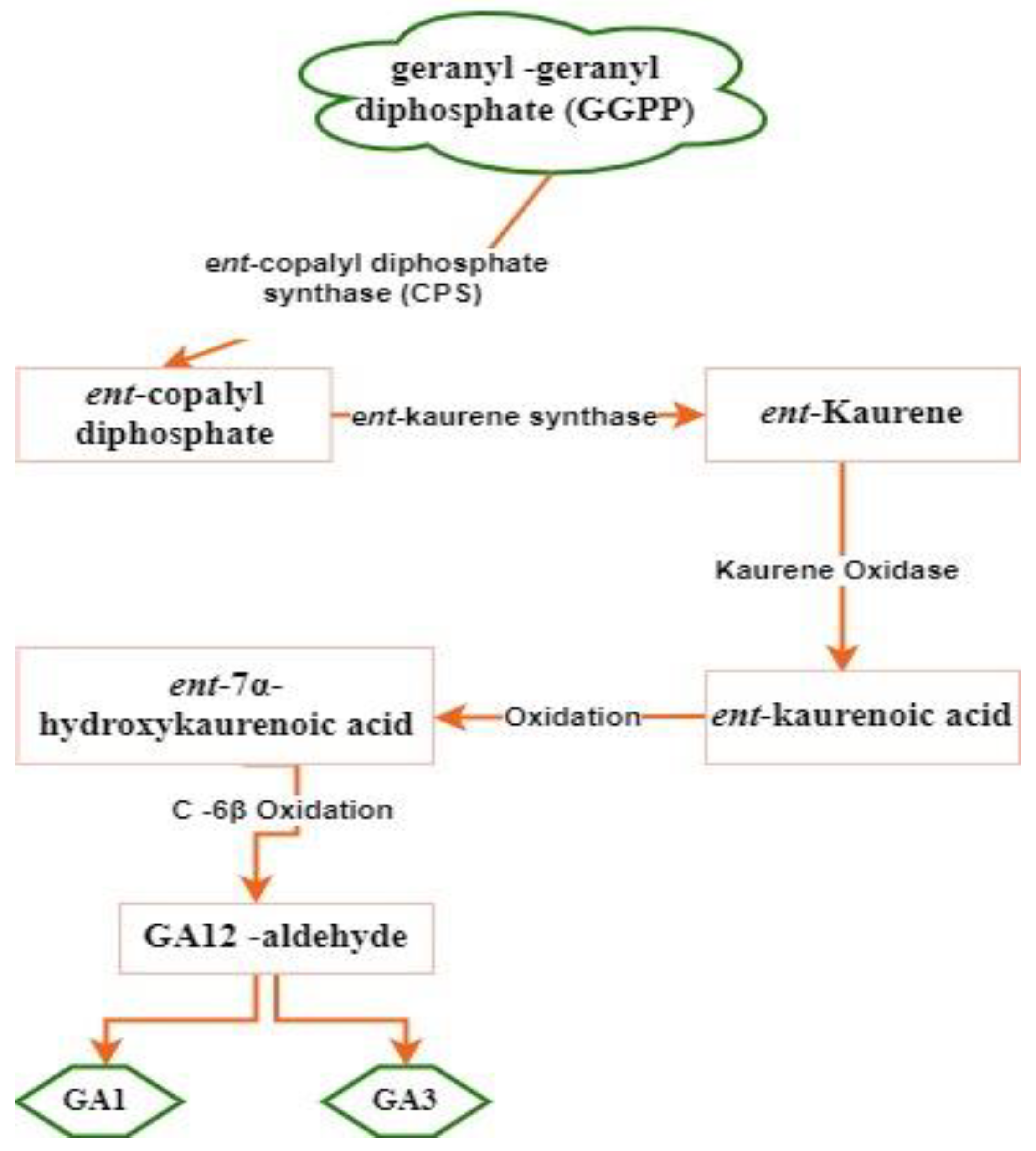
- Gibberellins

- Abscisic acid
- Ethylene
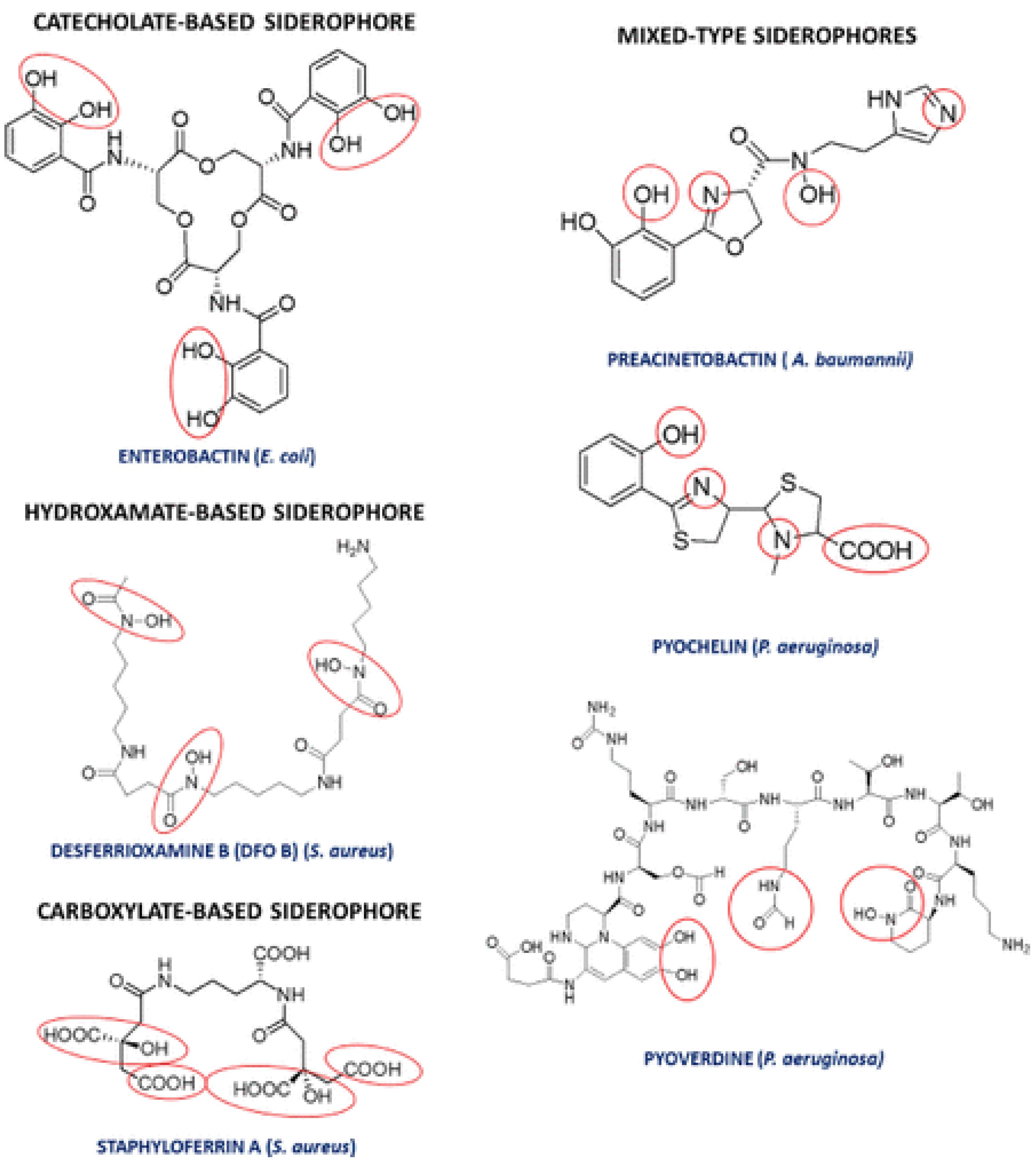
4.1.4. Siderophores Production
4.2. Indirect Mechanism
4.2.1. Antibiotics Synthesis
4.2.2. Induction of Systemic Resistance
- Systemic Acquired Resistance (SAR)
- Induced Systemic Resistance (ISR)
4.2.3. Production of Cell Wall-Degrading Enzymes
4.2.4. Production of Hydrogen Cyanide (HCN)
5. Examples of Successful Application of Rhizobia as Biofertilizer
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dent, D.; Cocking, E. Establishing symbiotic nitrogen fixation in cereals and other non-legume crops: The Greener Nitrogen Revolution. Agric. Food Secur. 2017, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Mehboob, I.; Naveed, M.; Zahir, Z.A. Rhizobial Association with Non-Legumes: Mechanisms and Applications. Crit. Rev. Plant Sci. 2009, 28, 432–456. [Google Scholar] [CrossRef]
- Hassan, M.K.; McInroy, J.A.; Kloepper, J.W. The Interactions of Rhizodeposits with Plant Growth-Promoting Rhizobacteria in the Rhizosphere: A Review. Agriculture 2019, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- González-López, O.; Ruano-Rosa, D. Root exudates, a key factor in the plant-bacteria interaction mechanisms. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Elsevier: Amsterdam, The Netherlands, 2020; pp. 111–121. [Google Scholar] [CrossRef]
- Mehboob, I.; Naveed, M.; Zahir, Z.A.; Ashraf, M. Potential of Rhizobia for Sustainable Production of Non-legumes. In Crop Production for Agricultural Improvement; Ashraf, M., Öztürk, M., Ahmad, M.S.A., Aksoy, A., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 659–704. [Google Scholar] [CrossRef]
- Antoun, H.; Beauchamp, C.J.; Goussard, N.; Chabot, R.; Lalande, R. Potential of Rhizobium and Bradyrhizobium species as plant growth promoting rhizobacteria on non-legumes: Effect on radishes (Raphanus sativus L.). In Molecular Microbial Ecology of the Soil; Hardarson, G., Broughton, W.J., Eds.; Springer: Dordrecht, The Netherlands, 1998; pp. 57–67. [Google Scholar] [CrossRef]
- Mishra, R.P.N.; Singh, R.K.; Jaiswal, H.K.; Kumar, V.; Maurya, S. Rhizobium-Mediated Induction of Phenolics and Plant Growth Promotion in Rice (Oryza sativa L.). Curr. Microbiol. 2006, 52, 383–389. [Google Scholar] [CrossRef]
- Yanni, Y.G.; Rizk, R.Y.; El-Fattah, F.K.A.; Squartini, A.; Corich, V.; Giacomini, A.; de Bruijn, F.; Rademaker, J.; Maya-Flores, J.; Ostrom, P.; et al. The beneficial plant growth-promoting association of Rhizobium leguminosarum bv. trifolii with rice roots. Funct. Plant Biol. 2001, 28, 845–870. [Google Scholar] [CrossRef]
- Gutiérrez-Zamora, M. Natural endophytic association between Rhizobium etli and maize (Zea mays L.). J. Biotechnol. 2001, 91, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Chabot, R.; Antoun, H.; Cescas, M.P. Growth promotion of maize and lettuce by phosphate-solubilizing Rhizobium leguminosarum biovar. phaseoli. Plant Soil 1996, 184, 311–321. [Google Scholar] [CrossRef]
- Peix, A.; Rivas-Boyero, A.; Mateos, P.; Rodriguez-Barrueco, C.; Martínez-Molina, E.; Velazquez, E. Growth promotion of chickpea and barley by a phosphate solubilizing strain of Mesorhizobium mediterraneum under growth chamber conditions. Soil Biol. Biochem. 2001, 33, 103–110. [Google Scholar] [CrossRef]
- Alami, Y.; Achouak, W.; Marol, C.; Heulin, T. Rhizosphere Soil Aggregation and Plant Growth Promotion of Sunflowers by an Exopolysaccharide-Producing Rhizobium sp. Strain Isolated from Sunflower Roots. Appl. Environ. Microbiol. 2000, 66, 3393–3398. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.; Bisht, S.; Sood, A.; Aeron, A.; Sharma, G.D.; Maheshwari, D.K. Consortium of Plant-Growth-Promoting Bacteria: Future Perspective in Agriculture. In Bacteria in Agrobiology: Plant Probiotics; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 185–200. [Google Scholar] [CrossRef]
- Rochlani, A.; Dalwani, A.; Shaikh, N.; Shaikh, N.; Sharma, S.; Saraf, S.M. Plant Growth Promoting Rhizobacteria as Biofertilizers: Application in Agricultural Sustainability. Acta Sci. Microbiol. 2022, 5, 12–21. [Google Scholar] [CrossRef]
- Aloo, B.N.; Tripathi, V.; Makumba, B.A.; Mbega, E.R. Plant growth-promoting rhizobacterial biofertilizers for crop production: The past, present, and future. Front. Plant Sci. 2022, 13, 1002448. [Google Scholar] [CrossRef]
- Hayat, R.; Ahmed, I.; Sheirdil, R.A. An Overview of Plant Growth Promoting Rhizobacteria (PGPR) for Sustainable Agriculture. In Crop Production for Agricultural Improvement; Ashraf, M., Öztürk, M., Ahmad, M.S.A., Aksoy, A., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 557–579. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Valle, A.; López-Calleja, A.C.; Alvarez-Venegas, R. Enhancement of Pathogen Resistance in Common Bean Plants by Inoculation with Rhizobium etli. Front. Plant Sci. 2019, 10, 1317. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Viveros, O.; Jorquera, M.; Crowley, D.; Gajardo, G.; Mora, M. Mechanisms and Practical Considerations Involved In Plant Growth Promotion by Rhizobacteria. J. Soil Sci. Plant Nutr. 2010, 10, 293–319. [Google Scholar] [CrossRef] [Green Version]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2023, 255, 571–586. [Google Scholar] [CrossRef]
- Kalyanasundaram, G.T.; Syed, N.; Subburamu, K. Recent developments in plant growth-promoting rhizobacteria (PGPR) for sustainable agriculture. In Recent Developments in Applied Microbiology and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2021; pp. 181–192. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ. Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Vargas, L.K.; Volpiano, C.G.; Lisboa, B.B.; Giongo, A.; Beneduzi, A.; Passaglia, L.M.P. Potential of Rhizobia as Plant Growth-Promoting Rhizobacteria. In Microbes for Legume Improvement; Zaidi, A., Khan, M.S., Musarrat, J., Eds.; Springer International Publishing: Cham, Germany, 2017; pp. 153–174. [Google Scholar] [CrossRef]
- Hassen, W.; Neifar, M.; Cherif, H.; Najjari, A.; Chouchane, H.; Driouich, R.C.; Salah, A.; Naili, F.; Mosbah, A.; Souissi, Y.; et al. Pseudomonas rhizophila S211, a New Plant Growth-Promoting Rhizobacterium with Potential in Pesticide-Bioremediation. Front. Microbiol. 2018, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Murray, E.W.; Greenberg, B.M.; Cryer, K.; Poltorak, B.; McKeown, J.; Spies, J.; Gerwing, P.D. Kinetics of phytoremediation of petroleum hydrocarbon contaminated soil. Int. J. Phytorem. 2019, 21, 27–33. [Google Scholar] [CrossRef]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant growth promoting rhizobacteria (PGPR): Current and future prospects for development of sustainable agriculture. J. Microb. Biochem. Technol. 2015, 7, 96–102. [Google Scholar] [CrossRef]
- Brannen, P.M.; Kenney, D.S. Kodiak®—A successful biological-control product for suppression of soil-borne plant pathogens of cotton. J. Ind. Microbiol. Biotechnol. 1997, 19, 169–171. [Google Scholar] [CrossRef]
- Reiss, A.; Jørgensen, L.N. Biological control of yellow rust of wheat (Puccinia striiformis) with Serenade ® ASO (Bacillus subtilis strain QST713). Crop. Prot. 2017, 93, 1–8. [Google Scholar] [CrossRef]
- Jeyanthi, V.; Kanimozhi, S. Plant Growth Promoting Rhizobacteria (PGPR)-Prospective and Mechanisms: A Review. J. Pure Appl. Microbiol. 2018, 12, 733–749. [Google Scholar] [CrossRef]
- Paudyal, S.P.; Gupta, V. Substitution of chemical fertilizer nitrogen through Rhizobium inoculation technology. Our Nat. 2018, 16, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Bharti, N.; Sharma, S.K.; Saini, S.; Verma, A.; Nimonkar, V.; Prakash, O. Microbial Plant Probiotics: Problems in Application and Formulation. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 317–335. [Google Scholar]
- Ledermann, R.; Schulte, C.C.M.; Poole, P.S. How Rhizobia Adapt to the Nodule Environment. J. Bacteriol. 2021, 203, e00539-20. [Google Scholar] [CrossRef]
- Helene, L.C.F.; Klepa, M.S.; Hungria, M. New Insights into the Taxonomy of Bacteria in the Genomic Era and a Case Study with Rhizobia. Int. J. Microbiol. 2022, 2022, 4623713. [Google Scholar] [CrossRef]
- De Lajudie, P.M.; Andrews, M.; Ardley, J.; Eardly, B.; Jumas-Bilak, E.; Kuzmanović, N.; Lassalle, F.; Lindström, K.; Mhamdi, R.; Martínez-Romero, E.; et al. Minimal standards for the description of new genera and species of rhizobia and agrobacteria. Int. J. Syst. Evol. Microbiol. 2019, 69, 1852–1863. [Google Scholar] [CrossRef]
- Chen, W.F.; Wang, E.T.; Ji, Z.J.; Zhang, J.J. Recent development and new insight of diversification and symbiosis specificity of legume rhizobia: Mechanism and application. J. Appl. Microbiol. 2021, 131, 553–563. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Mohammed, M.; Ibny, F.Y.I.; Dakora, F.D. Rhizobia as a Source of Plant Growth-Promoting Molecules: Potential Applications and Possible Operational Mechanisms. Front. Sustain. Food Syst. 2021, 4, 619676. [Google Scholar] [CrossRef]
- Wardell, G.E.; Hynes, M.F.; Young, P.J.; Harrison, E. Why are rhizobial symbiosis genes mobile? Philos. Trans. R. Soc. B Biol. Sci. 2022, 377, 20200471. [Google Scholar] [CrossRef]
- Bellabarba, A.; Fagorzi, C.; Dicenzo, G.C.; Pini, F.; Viti, C.; Checcucci, A. Deciphering the Symbiotic Plant Microbiome: Translating the Most Recent Discoveries on Rhizobia for the Improvement of Agricultural Practices in Metal-Contaminated and High Saline Lands. Agronomy 2019, 9, 529. [Google Scholar] [CrossRef] [Green Version]
- Soto, M.J.; Staehelin, C.; Gourion, B.; Cárdenas, L.; Vinardell, J.M. Editorial: Early signaling in the Rhizobium-legume symbiosis. Front. Plant Sci. 2022, 13, 1056830. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H. Root nodules of legumes: A suitable ecological niche for isolating non-rhizobial bacteria with biotechnological potential in agriculture. Curr. Res. Biotechnol. 2022, 4, 78–86. [Google Scholar] [CrossRef]
- Quilbé, J.; Montiel, J.; Arrighi, J.-F.; Stougaard, J. Molecular Mechanisms of Intercellular Rhizobial Infection: Novel Findings of an Ancient Process. Front. Plant Sci. 2022, 13, 922982. [Google Scholar] [CrossRef]
- Rae, A.E.; Rolland, V.; White, R.G.; Mathesius, U. New methods for confocal imaging of infection threads in crop and model legumes. Plant Methods 2021, 17, 24. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, J.; Zhu, H. Genetic and Molecular Mechanisms Underlying Symbiotic Specificity in Legume-Rhizobium Interactions. Front. Plant Sci. 2018, 9, 313. [Google Scholar] [CrossRef]
- Gage, D.J. Infection and Invasion of Roots by Symbiotic, Nitrogen-Fixing Rhizobia during Nodulation of Temperate Legumes. Microbiol. Mol. Biol. Rev. 2004, 68, 280–300. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, S.; Kaneko, T.; Sato, S.; Saeki, K. Hijacking of leguminous nodulation signaling by the rhizobial type III secretion system. Proc. Natl. Acad. Sci. USA 2013, 110, 17131–17136. [Google Scholar] [CrossRef] [Green Version]
- Alazard, D.; Duhoux, E. Development of Stem Nodules in a Tropical Forage Legume, Aeschynomene afraspera. J. Exp. Bot. 1990, 41, 1199–1206. [Google Scholar] [CrossRef] [Green Version]
- Tulumello, J.; Chabert, N.; Rodriguez, J.; Long, J.; Nalin, R.; Achouak, W.; Heulin, T. Rhizobium alamii improves water stress tolerance in a non-legume. Sci. Total. Environ. 2021, 797, 148895. [Google Scholar] [CrossRef]
- Sijilmassi, B.; Filali-Maltouf, A.; Fahde, S.; Ennahli, Y.; Boughribil, S.; Kumar, S.; Amri, A. In-Vitro Plant Growth Promotion of Rhizobium Strains Isolated from Lentil Root Nodules under Abiotic Stresses. Agronomy 2020, 10, 1006. [Google Scholar] [CrossRef]
- Akhtar, M.S. (Ed.) Salt Stress, Microbes, and Plant Interactions: Causes and Solution; Springer: Singapore, 2019; Volume 1. [Google Scholar] [CrossRef]
- Barquero, M.; Poveda, J.; Laureano-Marín, A.M.; Ortiz-Liébana, N.; Brañas, J.; González-Andrés, F. Mechanisms involved in drought stress tolerance triggered by rhizobia strains in wheat. Front. Plant Sci. 2022, 13, 1036973. [Google Scholar] [CrossRef]
- Sindhu, S.; Dahiya, A.; Gera, R.; Sindhu, S.S. Mitigation of Abiotic Stress in Legume-Nodulating Rhizobia for Sustainable Crop Production. Agric. Res. 2020, 9, 444–459. [Google Scholar] [CrossRef]
- Lindström, K.; Mousavi, S.A. Effectiveness of nitrogen fixation in rhizobia. Microb. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.K.; Gowda, C.L.; Krishnamurthy, L. Plant growth promoting rhizobia: Challenges and opportunities. 3 Biotech 2015, 5, 355–377. [Google Scholar] [CrossRef] [Green Version]
- Laslo, É.; Mara, G. Is PGPR an Alternative for NPK Fertilizers in Sustainable Agriculture? In Microbial Interventions in Agriculture and Environment; Singh, D.P., Gupta, V.K., Prabha, R., Eds.; Springer: Singapore, 2019; pp. 51–62. [Google Scholar] [CrossRef]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total. Environ. 2020, 743, 140682. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Ruiz-Lozano, J. Induction of Plant Tolerance to Semi-arid Environments by Beneficial Soil Microorganisms—A Review. In Climate Change, Intercropping, Pest Control and Beneficial Microorganisms; Lichtfouse, E., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 121–135. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Al-Mallah, M.K.; Davey, M.R.; Cocking, E.C. Nodulation of Oilseed Rape (Brassica napus) by Rhizobia. J. Exp. Bot. 1990, 41, 1567–1572. [Google Scholar] [CrossRef]
- Tahir, M.; Sarwar, M.A. Plant Growth Promoting Rhizobacteria (PGPR): A Budding Complement of Synthetic Fertilizers for Improving Crop Production. Group 2013, 19, 79–87. [Google Scholar]
- Sharma, D.; Gahtyari, N.C.; Chhabra, R.; Kumar, D. Role of Microbes in Improving Plant Growth and Soil Health for Sustainable Agriculture. In Advances in Plant Microbiome and Sustainable Agriculture; Yadav, A.N., Rastegari, A.A., Yadav, N., Kour, D., Eds.; Microorganisms for Sustainability; Springer: Singapore, 2020; Volume 19, pp. 207–256. [Google Scholar] [CrossRef]
- Tailor, A.J.; Joshi, B.H. Harnessing Plant Growth Promoting Rhizobacteria Beyond Nature: A Review. J. Plant Nutr. 2014, 37, 1534–1571. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of Plant Growth Promoting Rhizobacteria in Grain Legumes: Growth Promotion and Crop Production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef]
- Prabhu, N.; Borkar, S.; Garg, S. Phosphate solubilization by microorganisms. In Advances in Biological Science Research; Elsevier: Amsterdam, The Netherlands, 2019; pp. 161–176. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Romano, S.; Pocsfalvi, G.; Fiume, I.; Esposito, R.; Angelini, C.; Bianco, C. Bacterial IAA-Delivery into Medicago Root Nodules Triggers a Balanced Stimulation of C and N Metabolism Leading to a Biomass Increase. Microorganisms 2019, 7, 403. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Ghosh, P.; Maiti, T.K. Production and Metabolism of Indole Acetic Acid (IAA) by Root Nodule Bacteria (Rhizobium): A Review. Appl. Microbiol. 2011, 5, 523–540. [Google Scholar]
- Silini, A.R.; Parolini, O.; Huppertz, B.; Lang, I. Soluble Factors of Amnion-Derived Cells in Treatment of Inflammatory and Fibrotic Pathologies. Curr. Stem Cell Res. Ther. 2013, 8, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, J.A. (Ed.) Handbook of Research on Microbial Remediation and Microbial Biotechnology for Sustainable Soil. In Advances in Environmental Engineering and Green Technologies; IGI Global: Pennsylvania, PA, USA, 2021. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [Green Version]
- Gray, E.J.; Smith, D.L. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant–bacterium signaling processes. Soil. Biol. Biochem. 2005, 37, 395–412. [Google Scholar] [CrossRef]
- Pahari, A.; Pradhan, A.; Nayak, S.; Mishra, B. Plant Growth Promoting Rhizobacteria (Pgpr): Prospects and Application. In Frontiers in Soil and Environmental Microbiology, 1st ed.; Nayak, S.K., Mishra, B.B., Eds.; CRC Press: Boca Raton, FL, USA, 2020; Volume 56, pp. 47–56. [Google Scholar] [CrossRef]
- Medeot, D.B.; Paulucci, N.S.; Albornoz, A.I.; Fumero, M.V.; Bueno, M.A.; Garcia, M.B.; Woelke, M.R.; Okon, Y.; Dardanelli, M.S. Plant Growth Promoting Rhizobacteria Improving the Legume–Rhizobia Symbiosis. In Microbes for Legume Improvement; Khan, M.S., Musarrat, J., Zaidi, A., Eds.; Springer: Vienna, Austria, 2010; pp. 473–494. [Google Scholar] [CrossRef]
- Nieto, K.F.; Frankenberger, W.T. Influence of adenine, isopentyl alcohol and Azotobacter chroococcum on the vegetative growth of Zea mays. Plant Soil 1991, 135, 213–221. [Google Scholar] [CrossRef]
- Kisiala, A.; Laffont, C.; Emery, R.J.N.; Frugier, F. Bioactive Cytokinins Are Selectively Secreted by Sinorhizobium meliloti Nodulating and Nonnodulating Strains. Mol. Plant-Microbe Interact. 2013, 26, 1225–1231. [Google Scholar] [CrossRef] [Green Version]
- Jan, B.; Sajad, S.; Reshi, Z.A.; Mohiddin, F.A. Plant Growth Promoting Rhizobacteria (PGPR): Eco-Friendly Approach for Sustainable Agriculture. In Plant-Microbe Dynamics: Recent Advances for Sustainable Agriculture, 1st ed.; Pirzadah, T.B., Malik, B., Hakeem, K.R., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 185–200. [Google Scholar] [CrossRef]
- Tatsukami, Y.; Ueda, M. Rhizobial gibberellin negatively regulates host nodule number. Sci. Rep. 2016, 6, 27998. [Google Scholar] [CrossRef] [Green Version]
- Lucas, T.M.R.; Carlos, H.C.; Fabio, L.C.M.; Gustavo, V.M. Azospirillum spp. potential for maize growth and yield. Afr. J. Biotechnol. 2018, 17, 574–585. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-M.; Khan, A.L.; Waqas, M.; Asaf, S.; Lee, K.-E.; Park, Y.-G.; Kim, A.-Y.; Khan, M.A.; You, Y.-H.; Lee, I.-J. Integrated phytohormone production by the plant growth-promoting rhizobacterium Bacillus tequilensis SSB07 induced thermotolerance in soybean. J. Plant Interact. 2019, 14, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Dobbelaere, S.; Vanderleyden, J.; Okon, Y. Plant Growth-Promoting Effects of Diazotrophs in the Rhizosphere. Crit. Rev. Plant Sci. 2003, 22, 107–149. [Google Scholar] [CrossRef]
- Boiero, L.; Perrig, D.; Masciarelli, O.; Penna, C.; Cassán, F.; Luna, V. Phytohormone production by three strains of Bradyrhizobium japonicum and possible physiological and technological implications. Appl. Microbiol. Biotechnol. 2007, 74, 874–880. [Google Scholar] [CrossRef]
- Contesto, C.; Desbrosses, G.; Lefoulon, C.; Bena, G.; Borel, F.; Galland, M.; Gamet, L.; Varoquaux, F.; Touraine, B. Effects of rhizobacterial ACC deaminase activity on Arabidopsis indicate that ethylene mediates local root responses to plant growth-promoting rhizobacteria. Plant Sci. 2008, 175, 178–189. [Google Scholar] [CrossRef]
- Wani, S.P.; Gopalakrishnan, S. Plant Growth-Promoting Microbes for Sustainable Agriculture. In Plant Growth Promoting Rhizobacteria (PGPR): Prospects for Sustainable Agriculture; Sayyed, R.Z., Reddy, M.S., Antonius, S., Eds.; Springer: Singapore, 2019; pp. 19–45. [Google Scholar] [CrossRef] [Green Version]
- Kircheva, N.; Dudev, T. Gallium as an Antibacterial Agent: A DFT/SMD Study of the Ga3+/Fe3+ Competition for Binding Bacterial Siderophores. Inorg. Chem. 2020, 59, 6242–6254. [Google Scholar] [CrossRef]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.A.; Reddy, M.S.; El Enshasy, H.; Gafur, A.; et al. Bacterial Plant Biostimulants: A Sustainable Way towards Improving Growth, Productivity, and Health of Crops. Sustainability 2021, 13, 2856. [Google Scholar] [CrossRef]
- Omar, S.A.; Abd-Alla, M.H. Biocontrol of fungal root rot diseases of crop plants by the use of rhizobia and bradyrhizobia. Folia Microbiol. 1998, 43, 431–437. [Google Scholar] [CrossRef]
- Albelda-Berenguer, M.; Monachon, M.; Joseph, E. Siderophores: From Natural Roles to Potential Applications. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 193–225. [Google Scholar]
- Roy, N.; Chakrabartty, P.K. Effect of Aluminum on the Production of Siderophore by Rhizobium sp. (Cicer arietinum). Curr. Microbiol. 2000, 41, 5–10. [Google Scholar] [CrossRef]
- Rogers, N.J.; Carson, K.C.; Glenn, A.R.; Dilworth, M.J.; Poole, R.K. Alleviation of aluminum toxicity to Rhizobium leguminosarum bv. viciae by the hydroxamate siderophore vicibactin. Biometals 2001, 14, 59–66. [Google Scholar] [CrossRef]
- O'Hara, G.W.; Goss, T.J.; Dilworth, M.J.; Glenn, A.R. Maintenance of Intracellular pH and Acid Tolerance in Rhizobium meliloti. Appl. Environ. Microbiol. 1989, 55, 1870–1876. [Google Scholar] [CrossRef] [Green Version]
- Karthika, S.; Varghese, S.; Jisha, M.S. Exploring the efficacy of antagonistic rhizobacteria as native biocontrol agents against tomato plant diseases. 3 Biotech 2020, 10, 320. [Google Scholar] [CrossRef]
- Kenawy, A.; Dailin, D.J.; Abo-Zaid, G.A.; Malek, R.A.; Ambehabati, K.K.; Zakaria, K.H.N.; Sayyed, R.Z.; El Enshasy, H.A. Biosynthesis of Antibiotics by PGPR and Their Roles in Biocontrol of Plant Diseases. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Sayyed, R.Z., Ed.; Microorganisms for Sustainability; Springer: Singapore, 2019; Volume 13, pp. 1–35. [Google Scholar] [CrossRef]
- Breil, B.; Borneman, J.; Triplett, E.W. A newly discovered gene, tfuA, involved in the production of the ribosomally synthesized peptide antibiotic trifolitoxin. J. Bacteriol. 1996, 178, 4150–4156. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, U.; Purkayastha, R.P. Role of rhizobitoxine in protecting soybean roots from Macrophomina phaseolina infection. Can. J. Microbiol. 1984, 30, 285–289. [Google Scholar] [CrossRef]
- Elbadry, M.; Taha, R.M.; Eldougdoug, K.A.; Gamal-Eldin, H. Induction of systemic resistance in faba bean (Vicia faba L.) to bean yellow mosaic potyvirus (BYMV) via seed bacterization with plant growth promoting rhizobacteria. J. Plant Dis. Prot. 2006, 113, 247–251. [Google Scholar] [CrossRef]
- Dutta, S.; Mishra, A.; Kumar, B.D. Induction of systemic resistance against fusarial wilt in pigeon pea through interaction of plant growth promoting rhizobacteria and rhizobia. Soil Biol. Biochem. 2008, 40, 452–461. [Google Scholar] [CrossRef]
- Kumar, H.; Bajpai, V.K.; Dubey, R.; Maheshwari, D.; Kang, S.C. Wilt disease management and enhancement of growth and yield of Cajanus cajan (L) var. Manak by bacterial combinations amended with chemical fertilizer. Crop. Prot. 2010, 29, 591–598. [Google Scholar] [CrossRef]
- Deshwal, V.K.; Dubey, R.C.; Maheshwari, D.K. Isolation of plant growth-promoting strains of Bradyrhizobium (Arachis) sp. with biocontrol potential against Macrophomina phaseolina causing charcoal rot of peanut. Curr. Sci. 2023, 84, 3. [Google Scholar]
- Abd El-Rahman, A.F.; Shaheen, H.A.; Abd El-Aziz, R.M.; Ibrahim, D.S.S. Influence of hydrogen cyanide-producing rhizobacteria in controlling the crown gall and root-knot nematode, Meloidogyne incognita. Egypt. J. Biol. Pest Control 2019, 29, 41. [Google Scholar] [CrossRef]
- Mishra, J.; Arora, N.K. Secondary metabolites of fluorescent pseudomonads in biocontrol of phytopathogens for sustainable agriculture. Appl. Soil Ecol. 2018, 125, 35–45. [Google Scholar] [CrossRef]
- Riaz, U.; Murtaza, G.; Anum, W.; Samreen, T.; Sarfraz, M.; Nazir, M.Z. Plant Growth-Promoting Rhizobacteria (PGPR) as Biofertilizers and Biopesticides. In Microbiota and Biofertilizers; Hakeem, K.R., Dar, G.H., Mehmood, M.A., Bhat, R.A., Eds.; Springer International Publishing: Cham, Germany, 2021. [Google Scholar] [CrossRef]
- Soumare, A.; Diédhiou, A.G.; Arora, N.K.; Al-Ani, L.K.T.; Ngom, M.; Fall, S.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L.; Sy, M.O. Potential Role and Utilization of Plant Growth Promoting Microbes in Plant Tissue Culture. Front. Microbiol. 2021, 12, 649878. [Google Scholar] [CrossRef] [PubMed]
- Mitter, E.K.; Tosi, M.; Obregón, D.; Dunfield, K.E.; Germida, J.J. Rethinking Crop Nutrition in Times of Modern Microbiology: Innovative Biofertilizer Technologies. Front. Sustain. Food Syst. 2021, 5, 606815. [Google Scholar] [CrossRef]
- Hiltner, L. Inoculation of the Soil for Cultivating Leguminous Plants. Available online: https://patentimages.storage.googleapis.com/49/98/c3/5cb3d700750937/US570813.pdf (accessed on 14 April 2023).
- Savala, C.E.N.; Wiredu, A.N.; Chikoye, D.; Kyei-Boahen, S. Prospects and Potential of Bradyrhizobium diazoefficiens Based Bio-Inoculants on Soybean Production in Different Agro-Ecologies of Mozambique. Front. Sustain. Food Syst. 2022, 6, 908231. [Google Scholar] [CrossRef]
- Ismail, S.; Dhamak, A.L.; Mohanty, S.R. Technical Bulletin—Rhizobium Biofertilizer Technology for Legumes of Maharashtra; AINP SBB Technical Bulletin VNMKV: Parbhani, India, 2021. [Google Scholar] [CrossRef]
- Dashadi, M.; Khosravi, H.; Moezzi, A.; Nadian, H.; Heidari, M.; Radjabi, R. Co-Inoculation of Rhizobium and Azotobacter on Growth of Faba bean. Environ. Sci. 2011, 1, 314–319. [Google Scholar]
- Gan, Y.; Warkentin, T.; Bing, D.; Stevenson, F.; McDonald, C. Chickpea water use efficiency in relation to cropping system, cultivar, soil nitrogen and Rhizobial inoculation in semiarid environments. Agric. Water Manag. 2010, 97, 1375–1381. [Google Scholar] [CrossRef]
- Chaintreuil, C.; Giraud, E.; Prin, Y.; Lorquin, J.; Bâ, A.; Gillis, M.; de Lajudie, P.; Dreyfus, B. Photosynthetic Bradyrhizobia Are Natural Endophytes of the African Wild Rice Oryza breviligulata. Appl. Environ. Microbiol. 2000, 66, 5437–5447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husssain, M.B.; Mehboob, I.; Zahir, Z.A.; Naveed, M.; Asghar, H.N. Potential of Rhizobium spp. for improving growth and yield of rice (Oryza sativa L.). Soil Environ. 2009, 15, 49–55. [Google Scholar]
- Egamberdiyeva, D.; Juraeva, D.; Poberejskaya, S.; Myachina, O.; Teryuhova, P.; Seydalieva, L.; Aliev, A. Improvement of wheat and cotton growth and nutrient uptake by phosphate solubilizing bacteria. In Proceedings of the 26th annual conservation tillage conference for sustainable agriculture, Auburn, Australia, 8 June 2004; pp. 58–65. [Google Scholar]
- Naveed, M.; Hussain, M.B.; Mehboob, I.; Zahir, Z.A. Rhizobial Amelioration of Drought Stress in Legumes. In Microbes for Legume Improvement; Zaidi, A., Khan, M.S., Musarrat, J., Eds.; Springer International Publishing: Cham, Germany, 2017; pp. 341–365. [Google Scholar] [CrossRef]
- Figueiredo, M.V.B.; Martinez, C.R.; Burity, H.A.; Chanway, C.P. Plant growth-promoting rhizobacteria for improving nodulation and nitrogen fixation in the common bean (Phaseolus vulgaris L.). World J. Microbiol. Biotechnol. 2008, 24, 1187–1193. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahde, S.; Boughribil, S.; Sijilmassi, B.; Amri, A. Rhizobia: A Promising Source of Plant Growth-Promoting Molecules and Their Non-Legume Interactions: Examining Applications and Mechanisms. Agriculture 2023, 13, 1279. https://doi.org/10.3390/agriculture13071279
Fahde S, Boughribil S, Sijilmassi B, Amri A. Rhizobia: A Promising Source of Plant Growth-Promoting Molecules and Their Non-Legume Interactions: Examining Applications and Mechanisms. Agriculture. 2023; 13(7):1279. https://doi.org/10.3390/agriculture13071279
Chicago/Turabian StyleFahde, Sara, Said Boughribil, Badreddine Sijilmassi, and Ahmed Amri. 2023. "Rhizobia: A Promising Source of Plant Growth-Promoting Molecules and Their Non-Legume Interactions: Examining Applications and Mechanisms" Agriculture 13, no. 7: 1279. https://doi.org/10.3390/agriculture13071279