
Response of New Yellow Lupin Varieties to Inoculation with Bradyrhizobium sp. Lupinus under Central European Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Growing Conditions
2.2. Inoculant Treatments
2.3. Agrotechnics
2.4. Plant Biometric Assessment
2.5. Chemical Analyses
2.6. Statistical Analysis
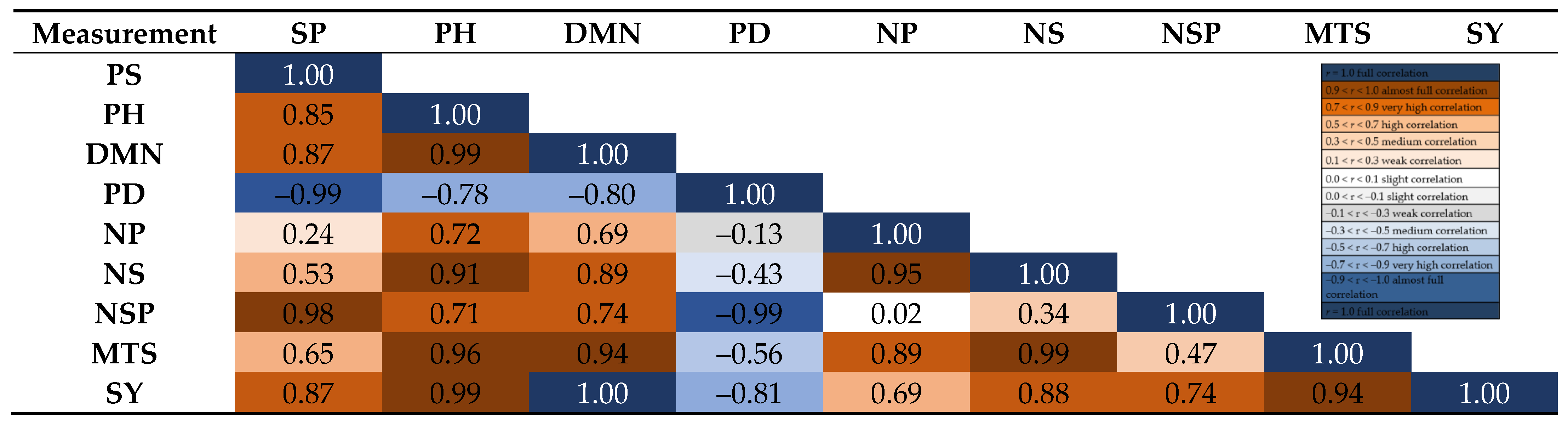
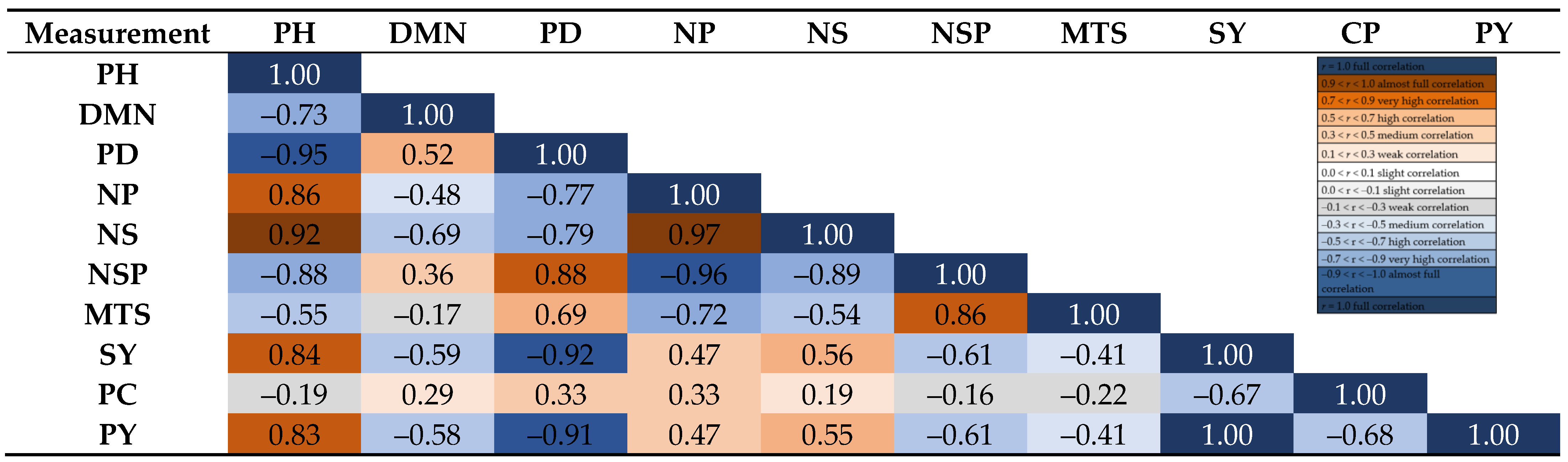
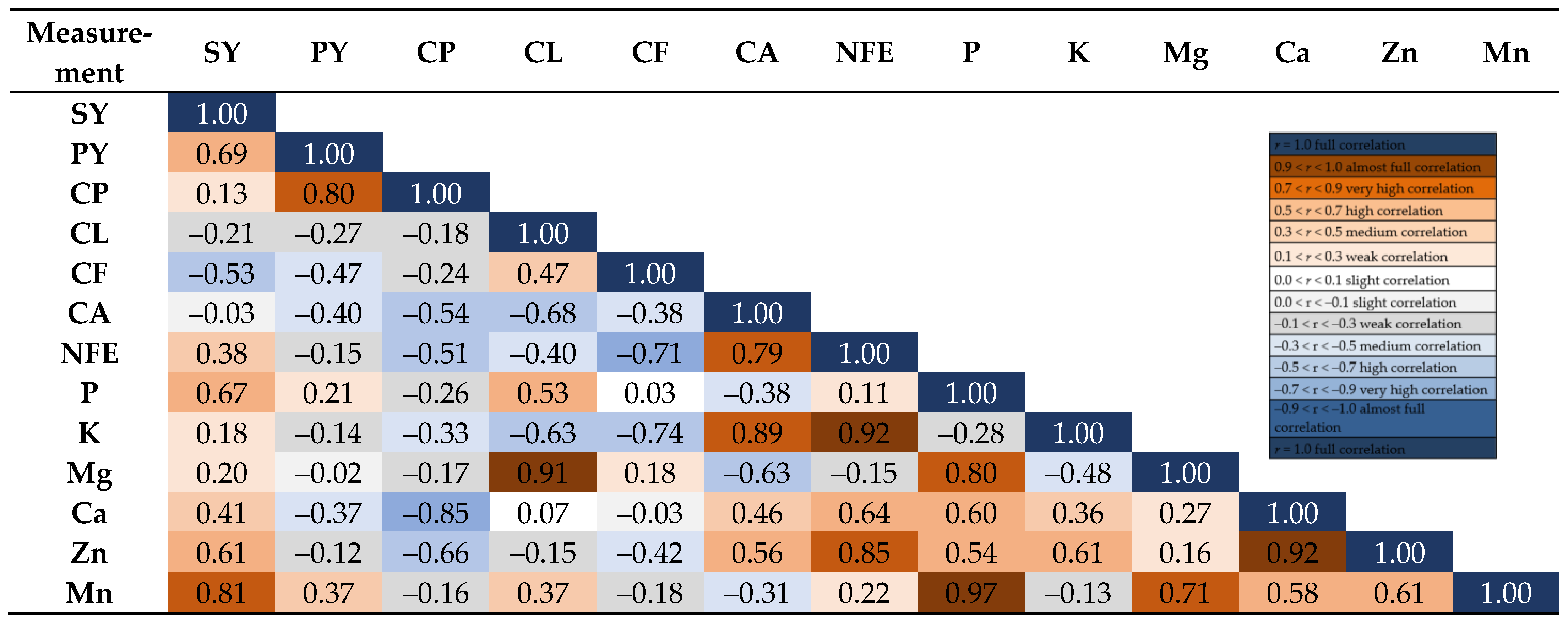
3. Results
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jarecki, W.; Migut, D. Comparison of yield and important seed quality traits of selected legume species. Agronomy 2022, 12, 2667. [Google Scholar] [CrossRef]
- Fox, R.H.; Myers, R.J.K.; Vallis, I. The nitrogen mineralization rate of legume residues in soil as influenced by their polyphenol, lignin, and nitrogen contents. Plant Soil 1990, 129, 251–259. [Google Scholar] [CrossRef]
- Chalk, P.M. Dynamics of biologically fixed N in legume-cereal rotation: A review. Aust. J. Agric. Res. 1998, 49, 303–316. [Google Scholar] [CrossRef]
- Isobe, K.; Ohte, N. Ecological perspectives on microbes involved in N-cycling. Microbes Environ. 2014, 29, 4–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.L.; Magthab, E.A.; Gleeson, D.B.; Hill, P.W.; Sánchez-Rodríguez, A.R.; Roberts, P.; Ge, T.; Murphy, D.V. Microbial competition for nitrogen and carbon is as intense in the subsoil as in the topsoil. Soil Biol. Biochem. 2018, 117, 72–82. [Google Scholar] [CrossRef]
- Peoples, M.B.; Craswell, E.T. Biological nitrogen fixation: Investments, expectations and actual contributions to agriculture. Plant Soil 1992, 141, 13–39. [Google Scholar] [CrossRef]
- Bebeli, P.J.; Lazaridi, E.; Chatzigeorgiou, T.; Suso, M.-J.; Hein, W.; Alexopoulos, A.A.; Canha, G.; van Haren, R.J.F.; Jóhannsson, M.H.; Mateos, C.; et al. State and progress of andean lupin cultivation in Europe: A review. Agronomy 2020, 10, 1038. [Google Scholar] [CrossRef]
- Mori, T.; Wachrinrat, C.; Staporn, D.; Aoyagi, R.; Meunpong, P.; Suebsai, W.; Boonsri, K.; Kitayama, K. Possibility of avoiding legumes-deriving boost of N2O emissions in tropical monoculture tree plantations. J. For. Res. 2023, 34, 565–573. [Google Scholar] [CrossRef]
- Jensen, R.J.; Joermsgaard, B.; Andersen, M.N.; Christiansen, J.L.; Morgensen, V.O.; Friis, P.; Peterson, C.T. The effect of lupins as compared with peas and oats on the yield of the subsequent winter barley crop. Eur. J. Agron. 2004, 20, 405–418. [Google Scholar] [CrossRef]
- Preissel, S.; Reckling, M.; Schläfke, N.; Zander, P. Magnitude and farm-economic value of grain pre-crop benefits in Europe. A review. Field Crops Res. 2015, 175, 64–79. [Google Scholar] [CrossRef] [Green Version]
- Faligowska, A.; Szymańska, G.; Panasiewicz, K.; Szukała, J.; Koziara, W.; Ratajczak, K. The long-term effect of legumes as forecrops on the productivity of rotation (winter rape-winter wheat-winter wheat) with nitrogen fertilization. Plant Soil Environ. 2019, 65, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Szymańska, G.; Faligowska, A.; Panasiewicz, K.; Szukała, J.; Ratajczak, K.; Sulewska, H. The long-term effect of legumes as forecrops on the productivity of rotation winter triticale–winter rape with nitrogen fertilisation. Acta Agric. Scand. Sec. B Soil Plant Sci. 2020, 70, 128–134. [Google Scholar] [CrossRef]
- Nemecek, T.; von Richthofen, J.-S.; Dubois, G.; Casta, P.; Charles, R.; Pahl, H. Environmental impacts of introducing grain leg umes into European crop rotations. Eur. J. Agron. 2008, 28, 380–393. [Google Scholar] [CrossRef]
- Kalembasa, D.; Szukała, J.; Symanowicz, B.; Kalembasa, S.; Faligowska, A.; Becher, M. Amount of biologically nitrogen fixed by faba bean and its uptake by winter wheat determined by 15N ID method. Arch. Agron. Sci. 2021, 67, 1875–1888. [Google Scholar] [CrossRef]
- Monteiro, A.; Miranda, C.; Trindade, H. Mediterranean Lupines as an Alternative Protein Source to Soybean. Biol. Life Sci. Forum 2021, 3, 38. [Google Scholar] [CrossRef]
- Panasiewicz, K. Chemical Composition of Lupin (Lupinus spp.) as Influenced by Variety and Tillage System. Agriculture 2022, 12, 263. [Google Scholar] [CrossRef]
- Wilmowicz, E.; Kućko, A.; Alché, J.D.D.; Czeszewska-Rosiak, G.; Florkiewicz, A.B.; Kapusta, M.; Karwaszewski, J. Remodeling of cell wall components in root nodules and flower abscission zone under drought in yellow lupine. Int. J. Mol. Sci. 2022, 23, 1680. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef] [Green Version]
- Pudełko, K.; Żarnicka, J. Diversity in symbiotic specificity of bacterial strains nodulating lupins in Poland. Pol. J. Agron. 2010, 2, 50–56. [Google Scholar]
- Prusiński, J.; Baturo-Cieśniewska, A.; Borowska, M. Response of soybean (Glycine max (L.) Merrill) to mineral nitrogen fertilization and Bradyrhizobium japonicum seed inoculation. Agronomy 2020, 10, 1300. [Google Scholar] [CrossRef]
- Księżak, J.; Bojarszczuk, J. The effect of mineral N fertilization and Bradyrhizobium japonicum seed inoculation on productivity of soybean (Glycine max (L.) Merrill). Agriculture 2022, 12, 110. [Google Scholar] [CrossRef]
- Ibrahim, H.M.; El-Sawah, A.M. The mode of integration between azotobacter and rhizobium affect plant growth, yield, and physiological responses of pea (Pisum sativum L.). J. Soil Sci. Plant Nutr. 2022, 22, 1238–1251. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Revision 4; Horwitz, W., Latimer, G.W., Jr., Eds.; AOAC International: Gaithersburg, MD, USA, 2011. [Google Scholar]
- Van Soest, P.J. Use of detergents in the analysis of fibrous feeds. II. A rapid method of determination of fibre and lignin. J. AOAC 1963, 4.6, 82. [Google Scholar]
- Erbas, M.; Certel, M.; Uslu, M.K. Some chemical properties of white lupin seeds (Lupinus albus L.). Food Chem. 2005, 89, 341–345. [Google Scholar] [CrossRef]
- Kebede, E. Contribution, utilization, and improvement of legumes-driven biological nitrogen fixation in agricultural systems. Front. Sustain. Food Syst. 2021, 5, 767998. [Google Scholar] [CrossRef]
- Rubiales, D.; Annicchiarico, P.; Vaz Patto, M.C.; Julier, B. Legume breeding for the agroecological transition of global agri-food systems: A european perspective. Front. Plant Sci. 2021, 12, 782574. [Google Scholar] [CrossRef] [PubMed]
- Księżak, J.; Bojarszczuk, M. The Productivity of Selected Species and Cultivars of Legumes Grown for Seeds in Organic Production System. Legume Crops—Characterization and Breeding for Improved Food Security; IntechOpen: London, UK, 2019; p. 82686. [Google Scholar] [CrossRef] [Green Version]
- Podleśna, A.; Podleśny, J.; Doroszewski, A. Usefulness of selected weather indices to evaluation of yellow lupine yielding possibility. Agric. Water Manag. 2014, 146, 201–207. [Google Scholar] [CrossRef]
- Csajbók, J.; Kutasy, E.T.; Melash, A.A.; Virág, I.C.; Ábrahám, É.B. Agro-biological traits of soybean cultivars, their yield quantity and quality under Central European conditions. Zemdirb. Agric. 2022, 109, 107–114. [Google Scholar] [CrossRef]
- Raza, S.; Jørnsgård, B.; Abdel-Wahab, A.; Christiansen, J. Effect of combined inoculation strains on growth of lupin on newly reclaimed land in Egypt. Biol. Fertil. Soils 2001, 34, 319–324. [Google Scholar] [CrossRef]
- Tounsi-Hammami, S.; Dhane-Fitouri, S.; Le Roux, C.; Hammami, Z.; Jeddi, F.B. Potential of native inoculum to improve the nodulation and growth of white lupin in Tunisia. Ann. del’ INRA 2020, 93, 102–114. [Google Scholar]
- Szpunar-Krok, E.; Wondołowska-Grabowska, A.; Bobrecka-Jamro, D.; Jańczak-Pieniążek, M.; Kotecki, A.; Kozak, M. Effect of nitrogen fertilisation and inoculation with Bradyrhizobium japonicum on the fatty acid profile of soybean (Glycine max (L.) Merrill) seeds. Agronomy 2021, 11, 941. [Google Scholar] [CrossRef]
- Szpunar-Krok, E.; Wondołowska-Grabowska, A. Quality evaluation indices for soybean oil in relation to cultivar, application of N fertiliser and seed inoculation with Bradyrhizobium japonicum. Foods 2022, 11, 762. [Google Scholar] [CrossRef] [PubMed]
- Zetochová, E.; Vollmannová, A.; Tirdiľová, I. Effect of inoculation on the content of biogenic elements in the white lupine and grass pea. Potravinarstvo Slovak J. Food Sci. 2020, 14, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Trivedi, A.; Nath, C.h.P.; Gupta, D.S.; Hazra, K.K. A comprehensive review on grain legumes as climate-smart crops: Challenges and prospects. Environ. Chall. 2022, 7, 100479. [Google Scholar] [CrossRef]
- Zveushe, O.K.; de Dios, V.R.; Zhang, H.; Zeng, F.; Liu, S.; Shen, S.; Kang, Q.; Zhang, Y.; Huang, M.; Sarfaraz, A.; et al. Effects of Co-Inoculating Saccharomyces spp. with Bradyrhizobium japonicum on Atmospheric Nitrogen Fixation in Soybeans (Glycine max (L.). Plants 2023, 12, 681. [Google Scholar] [CrossRef]
- de Borja Reis, A.F.; Rosso, L.M.; Purcell, L.C.; Naeve, S.; Casteel, S.N.; Kovács, P.; Archontoulis, S.; Davidson, D.; Ciampitti, I.A. Environmental factors associated with nitrogen fixation prediction in soybean. Front. Plant Sci. 2021, 12, 1013. [Google Scholar] [CrossRef]
- Campo, R.J.; Araujo, R.S.; Hungria, M. Molybdenum-enriched soybean seeds enhance N accumulation, seed yield, and seed protein content in Brazil. Field Crops Res. 2009, 110, 219–224. [Google Scholar] [CrossRef]
- Hungria, M.; Vargas, M.A.T. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Chalasani, D.; Basu, A.; Pullabhotla, S.V.S.R.N.; Jorrin, B.; Neal, A.L.; Poole, P.S.; Rao Podile, A.; Tkacz, A. Poor competitiveness of bradyrhizobium in pigeon pea root colonization in Indian soils. ASM J. 2021, 12, 4. [Google Scholar] [CrossRef]
- Podar, D.; Maathuis, F.J.M. Primary nutrient sensors in plants. iScience 2022, 25, 104029. [Google Scholar] [CrossRef]
- Marinkovic, J.; Bjelic, D.; Tintor, B.; Miladinovic, J.; Dukic, V.; Dordevic, V. Effects of soybean co-inoculation with plant growth promoting rhizobacteria in field trial. Rom. Biotechnol. Lett. 2018, 23, 13401–13408. [Google Scholar]
- Gresta, F.; Abbate, V.; Avola, G.; Magazzù, G.; Chiofalo, B. Lupin seed for the crop-livestock food chain. Ital. J. Agron. 2010, 4, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, K.; Biswas, B.; Kerr, P.G.; Blanchard, C.H.; Nabila, A.; Golder, M.; Gulzarul Aziz, M.; Farahnaky, A. Comparative assessment of nutritional, thermal, rheological and functional properties of nine Australian lupin cultivars. Sci. Rep. 2021, 11, 21515. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frías, J.; Martínez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.M.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 705. [Google Scholar] [CrossRef] [PubMed] [Green Version]






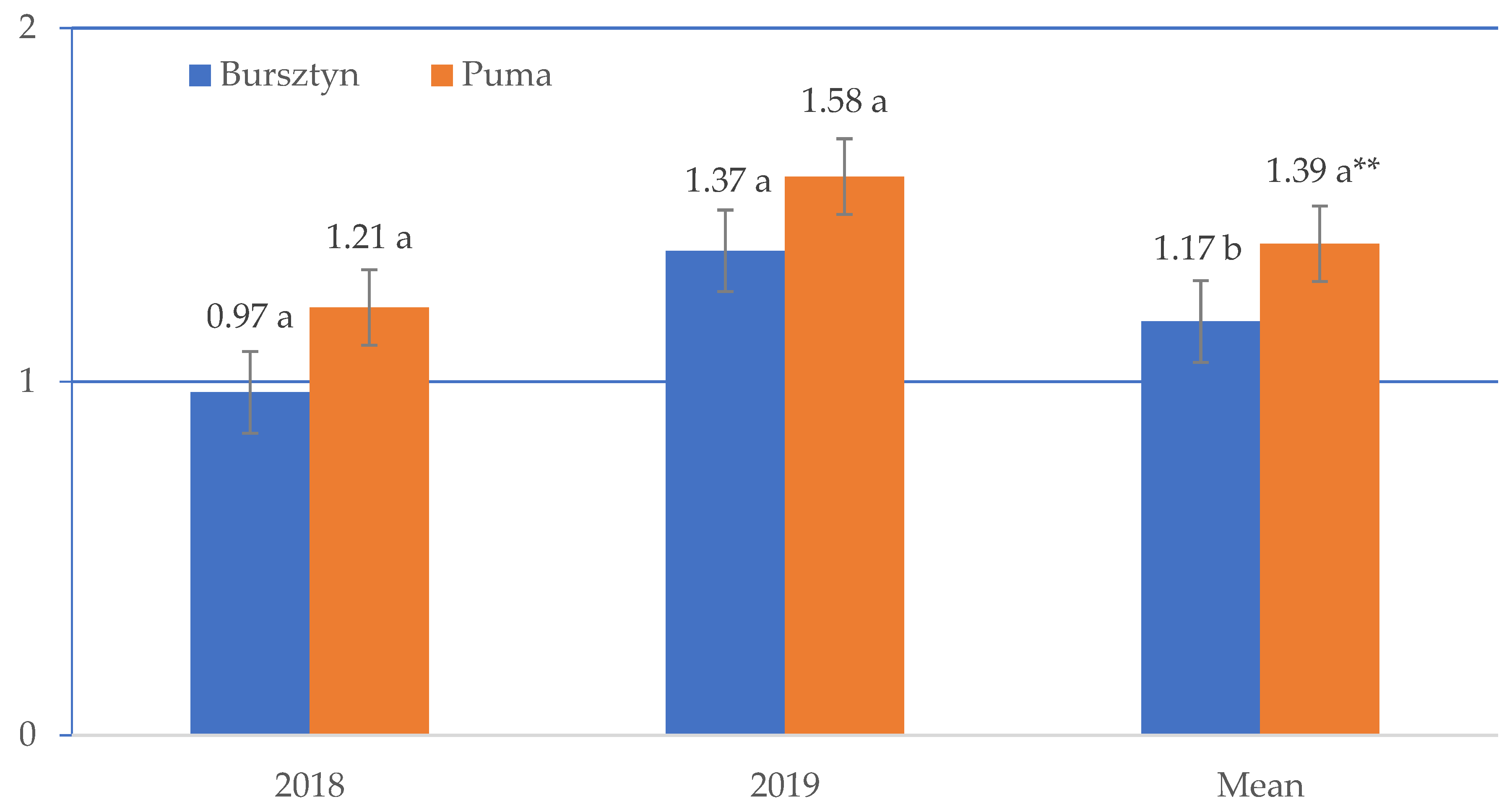{kind=link}
{kind=link}
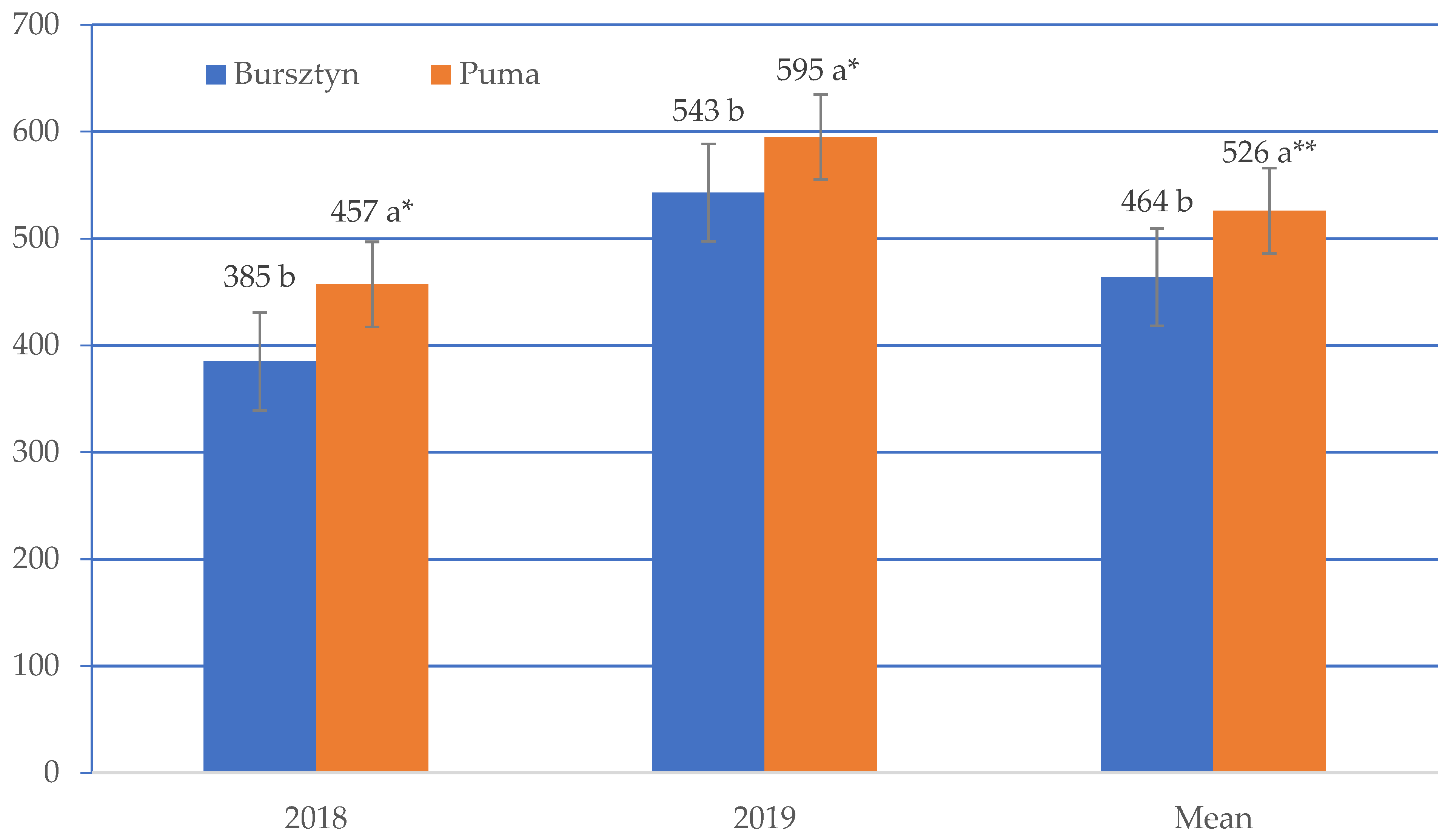{kind=link}
{kind=link}
{kind=link}
| Month | Precipitation Sum (mm) | Air Temperature (°C) | ||||
|---|---|---|---|---|---|---|
| Year | Year | |||||
| 2018 | 2019 | Long-Term Average | 2018 | 2019 | Long-Term Average | |
| April | 55.5 ** | 6.5 ** | 47.8 | 12.8 * | 9.6 * | 8.2 |
| May | 22.3 | 86.3 | 33.8 | 16.6 | 12.1 | 14.4 |
| Jun | 23.8 | 6.9 | 61.5 | 19.4 | 22.0 | 17.1 |
| July | 84.2 | 71.8 | 78.0 | 20.5 | 19.1 | 18.8 |
| Sum/Mean | 185.8 | 171.5 | 221.2 | 17.3 | 15.7 | 14.6 |
| Specification | PH | DMN | PD | NP | NS | NSP | MTS |
|---|---|---|---|---|---|---|---|
| Variety | |||||||
| Bursztyn | 53.4 | 1.4 | 66.7 | 8.2 | 24.4 | 3.0 | 125.6 |
| Puma | 60.1 | 1.0 | 77.6 | 6.5 | 20.9 | 3.2 | 133.3 |
| LSD value | 4.36 ** | 0.38 * | 1.49 ** | 1.35 * | 3.29 * | NS | 2.13 ** |
| Inoculant | |||||||
| Control | 56.1 | 1.2 | 71.6 | 7.5 | 22.6 | 3.0 | 129.1 |
| Nitragina | 56.7 | 1.2 | 72.1 | 7.2 | 22.3 | 3.1 | 128.4 |
| Nitroflora I | 57.3 | 1.1 | 74.0 | 6.6 | 21.0 | 3.2 | 130.6 |
| Nitroflora II | 57.4 | 1.3 | 69.0 | 7.2 | 21.3 | 3.0 | 128.9 |
| HiStick | 56.4 | 1.2 | 73.9 | 8.2 | 25.9 | 3.2 | 130.2 |
| LSD value | NS | NS | 3.09 * | 1.03 * | 2.96 * | NS | NS |
| Specification | Crude Protein | Crude Lipid | Crude Fiber | Crude Ash | N-Free Extract |
|---|---|---|---|---|---|
| Variety | |||||
| Bursztyn | 466.8 | 49.5 | 169.9 | 47.3 | 266.2 |
| Puma | 444.6 | 55.8 | 172.8 | 46.9 | 279.6 |
| LSD value | 3.97 ** | 4.32 * | NS | NS | NS |
| Inoculant | |||||
| Control | 451.7 | 49.1 | 164.0 | 48.4 | 286.6 |
| Nitragina | 451.2 | 53.8 | 183.6 | 47.1 | 264.0 |
| Nitroflora I | 459.1 | 53.6 | 160.6 | 47.0 | 279.5 |
| Nitroflora II | 447.1 | 54.8 | 175.6 | 46.8 | 275.4 |
| HiStick | 469.4 | 52.1 | 173.0 | 46.2 | 259.1 |
| LSD value | 19.88 ** | NS | 15.0 * | NS | NS |
| Phosphorus | Potassium | Magnesium | Calcium | Zinc *** | Manganese *** | |
|---|---|---|---|---|---|---|
| Control | 0.3 | 18.1 | 1.1 | 6.5 | 22.0 | 79.4 |
| Nitragina | 0.3 | 17.0 | 1.3 | 6.3 | 19.0 | 72.4 |
| Nitroflora I | 0.4 | 17.6 | 1.4 | 6.3 | 20.6 | 83.1 |
| Nitroflora II | 0.5 | 17.1 | 1.5 | 6.6 | 22.3 | 95.0 |
| HiStick | 0.4 | 16.8 | 1.3 | 6.1 | 18.6 | 82.6 |
| LSD value | 0.13 * | 1.19 ** | 0.15 ** | NS | NS | 12.89 * |
| Variety | Inoculant | ||||
|---|---|---|---|---|---|
| Control | Nitragina | Nitroflora I | Nitroflora II | HiStick | |
| Seed Yield | |||||
| Bursztyn | 1.18 | 1.09 | 1.16 | 1.20 | 1.21 |
| Puma | 1.40 | 1.39 | 1.40 | 1.41 | 1.37 |
| Mean | 1.29 | 1.24 | 1.28 | 1.30 | 1.39 |
| LSD value | I—NS; V × I—0.100 * | ||||
| Protein Yield | |||||
| Bursztyn | 455 | 444 | 467 | 461 | 491 |
| Puma | 533 | 503 | 529 | 524 | 538 |
| Mean | 494 | 475 | 498 | 492 | 515 |
| LSD value | I –27.5 *; V × I—38.8 * | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faligowska, A. Response of New Yellow Lupin Varieties to Inoculation with Bradyrhizobium sp. Lupinus under Central European Conditions. Agriculture 2023, 13, 1261. https://doi.org/10.3390/agriculture13061261
Faligowska A. Response of New Yellow Lupin Varieties to Inoculation with Bradyrhizobium sp. Lupinus under Central European Conditions. Agriculture. 2023; 13(6):1261. https://doi.org/10.3390/agriculture13061261
Chicago/Turabian StyleFaligowska, Agnieszka. 2023. "Response of New Yellow Lupin Varieties to Inoculation with Bradyrhizobium sp. Lupinus under Central European Conditions" Agriculture 13, no. 6: 1261. https://doi.org/10.3390/agriculture13061261