
Self-Regulation of Soil Enzyme Activity and Stoichiometry under Nitrogen Addition and Plastic Film Mulching in the Loess Plateau Area, Northwest China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatment
2.2. Sample Collection and Preservation
2.3. Soil Chemical Analysis
2.4. Soil Extracellular Enzyme Activity and Enzyme Stoichiometry Ratio
- EC:N = ln(βG):ln(NAG + LAP)
- EC:P = ln(βG):ln(AP)
- EN:P = ln(NAG + LAP):ln(AP)
- EC:N:P = ln(βG):ln(NAG + LAP):ln(AP)
2.5. Statistical Analysis
3. Results
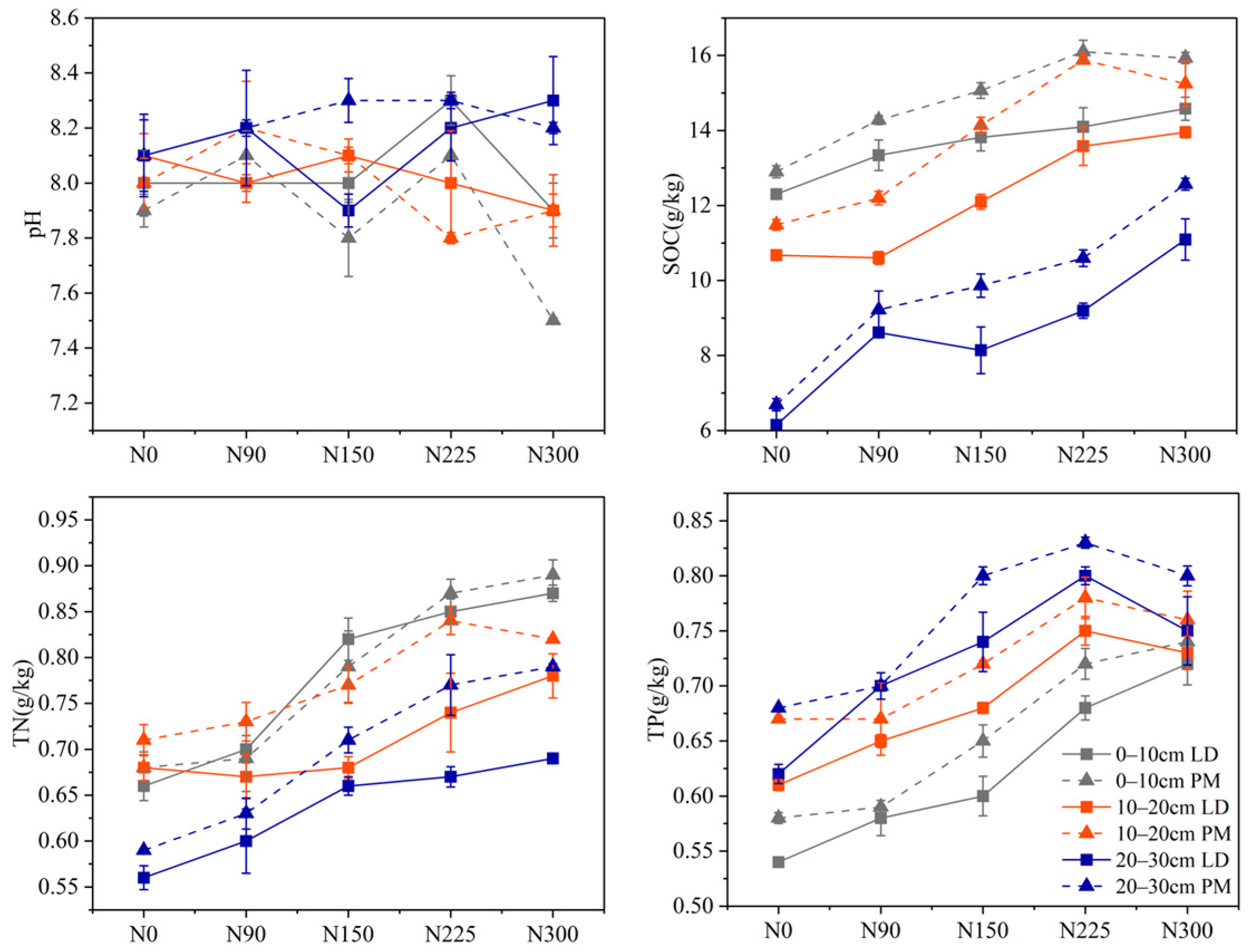
3.1. Soil Chemistry
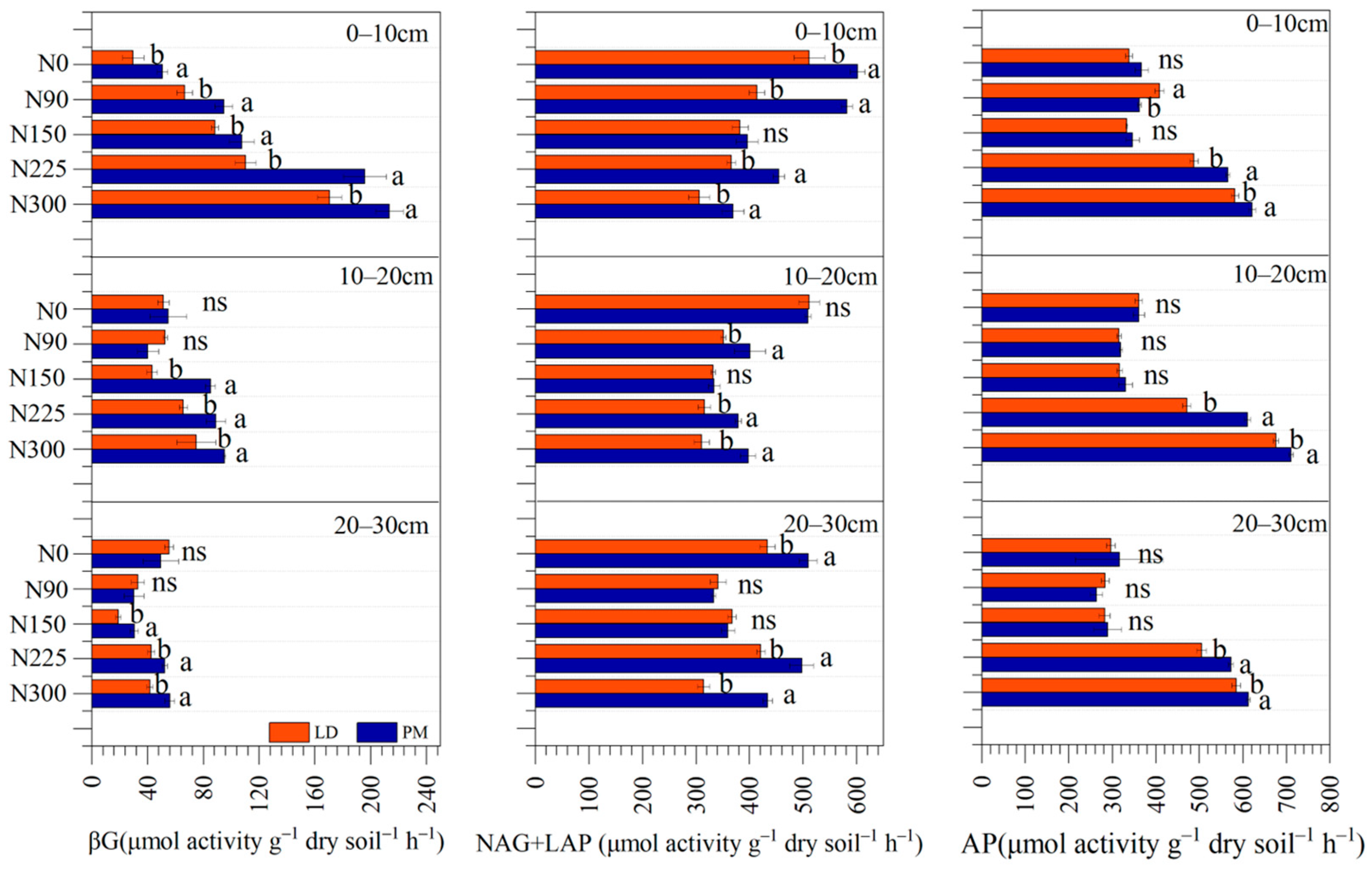
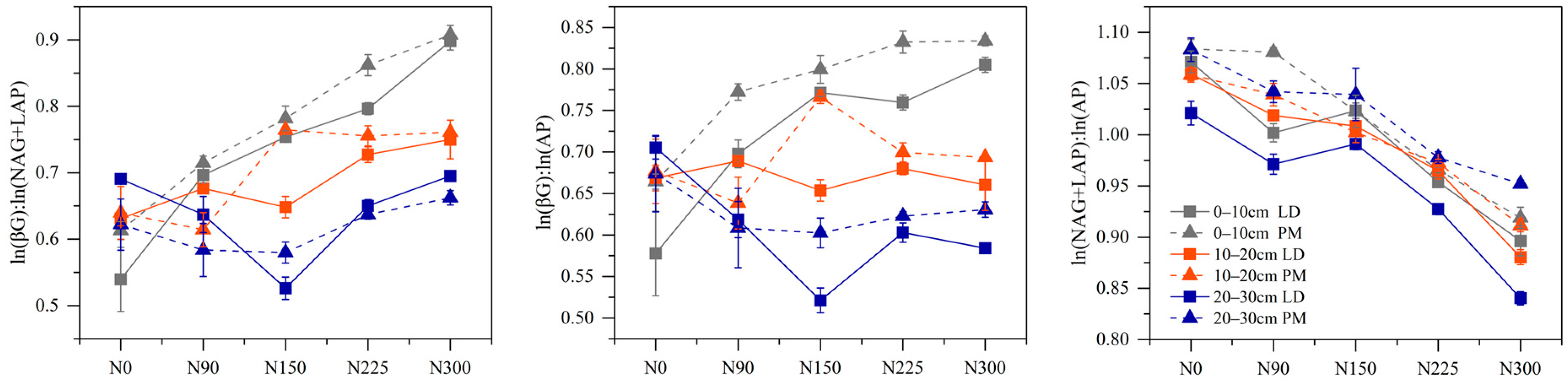
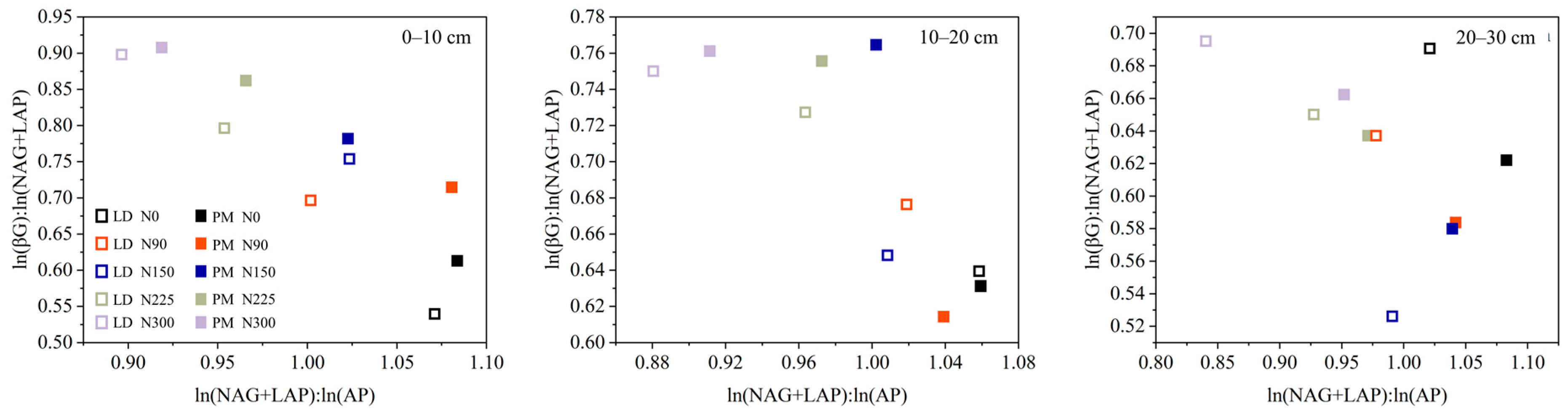
3.2. Soil Extracellular Enzyme Activities and Its Stoichiometry
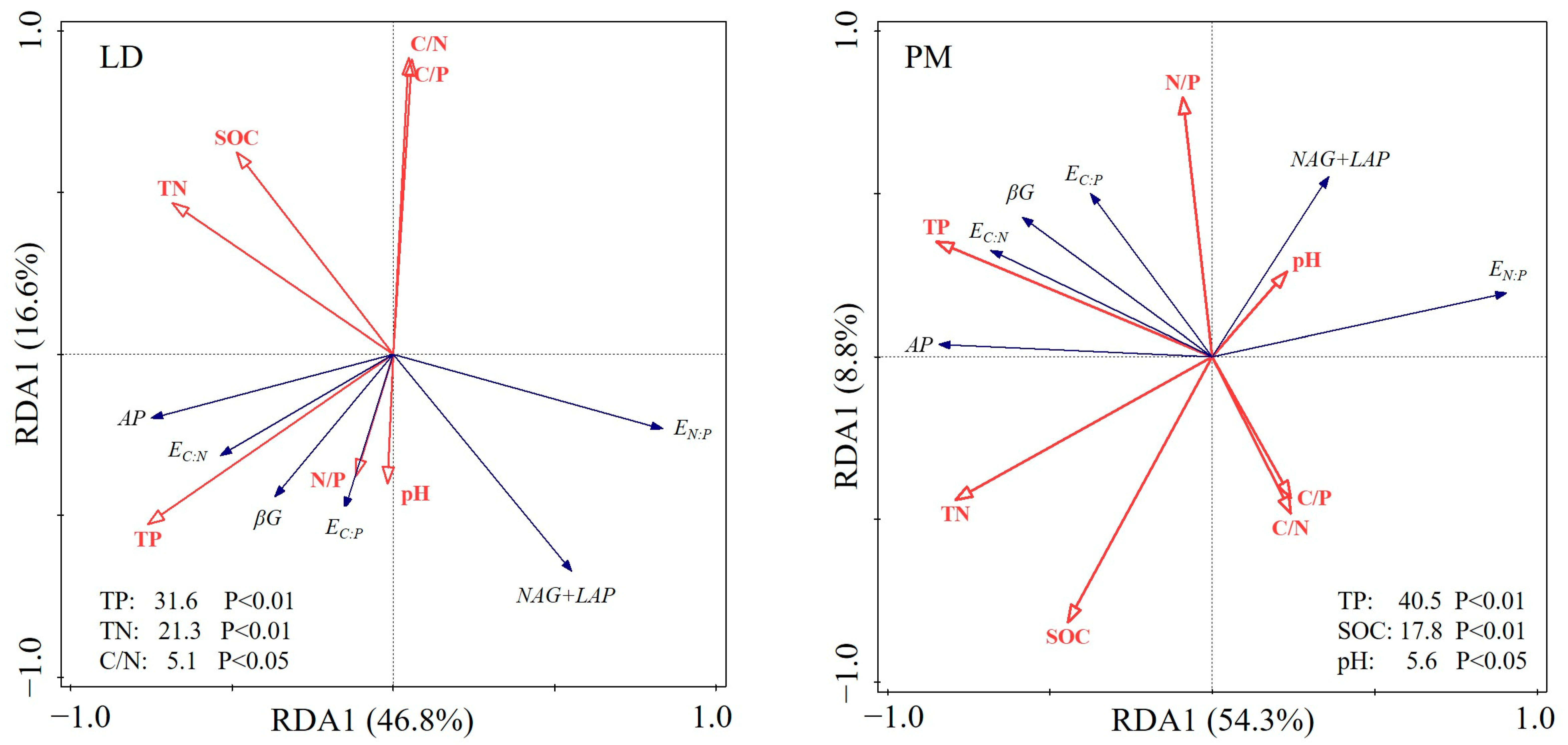
3.3. Relationship between Soil Chemical Properties and EA and ES
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sinsabaugh, R.L.; Gallo, M.E.; Lauber, C.; Waldrop, M.P.; Zak, D.R. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition. Biogeochemistry 2005, 75, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Zhong, Y.; Liu, J.; Zhu, G.; Shangguan, Z.; Yan, W. Effects of nitrogen enrichment on soil microbial characteristics: From biomass to enzyme activities. Geoderma 2020, 366, 114256. [Google Scholar] [CrossRef]
- Li, Y.; Nie, C.; Liu, Y.; Du, W.; He, P. Soil microbial community composition closely associates with specific enzyme activities and soil carbon chemistry in a long–term nitrogen fertilized grassland. Sci. Total Environ. 2019, 654, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L.; Follstad Shah, J.J. Ecoenzymatic stoichiometry and ecological theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 313–343. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Liu, S.; Shen, H.; Zhao, M.; Xu, L.; Xing, A.; Fang, J.; Sayer, E. Soil extracellular enzyme activity and stoichiometry in China’s forests. Funct. Ecol. 2020, 34, 1461–1471. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Hill, B.H.; Follstad Shah, J.J. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Gao, S.; Wang, P.; Qiu, J.; Shang, S. Impacts of simulated nitrogen deposition on soil enzyme activity in a northern temperate forest ecosystem depend on the form and level of added nitrogen. Eur. J. Soil Biol. 2021, 103, 103287. [Google Scholar] [CrossRef]
- Khan, S.A.; Mulvaney, R.L.; Ellsworth, T.R.; Boast, C.W. The myth of nitrogen fertilization for soil carbon sequestration. J. Environ. Qual. 2007, 36, 1821–1832. [Google Scholar] [CrossRef] [Green Version]
- Jian, S.; Li, J.; Chen, J.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta–analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Hou, E.; Chen, C.; Wen, D.; Liu, X. Phosphatase activity in relation to key litter and soil properties in mature subtropical forests in China. Sci. Total Environ. 2015, 515–516, 83–91. [Google Scholar] [CrossRef]
- Wang, B.; Xue, S.; Liu, G.B.; Zhang, G.H.; Li, G.; Ren, Z.P. Changes in soil nutrient and enzyme activities under different vegetations in the Loess Plateau area, Northwest China. Catena 2012, 92, 186–195. [Google Scholar] [CrossRef]
- Zhang, P.; Wei, T.; Cai, T.; Ali, S.; Han, Q.; Ren, X.; Jia, Z. Plastic–film mulching for enhanced water–use efficiency and economic returns from maize fields in semiarid China. Front. Plant Sci. 2017, 8, 512. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Yan, C.; Liu, Q.; Li, Z.; Yang, X.; Qi, R. Exploring optimal soil mulching to enhance yield and water use efficiency in maize cropping in China: A meta–analysis. Agric. Water Manag. 2019, 225, 105741. [Google Scholar] [CrossRef]
- Li, C.; Wen, X.; Wan, X.; Liu, Y.; Han, J.; Liao, Y.; Wu, W. Towards the highly effective use of precipitation by ridge–furrow with plastic film mulching instead of relying on irrigation resources in a dry semi–humid area. Field Crops Res. 2016, 188, 62–73. [Google Scholar] [CrossRef]
- Zheng, M.; Huang, J.; Chen, H.; Wang, H.; Mo, J. Responses of soil acid phosphatase and beta–glucosidase to nitrogen and phosphorus addition in two subtropical forests in southern China. Eur. J. Soil Biol. 2015, 68, 77–84. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, G.; Li, P.; Xue, S. Ecological stoichiometry of plant–soil–enzyme interactions drives secondary plant succession in the abandoned grasslands of Loess Plateau, China. Catena 2021, 202, 105302. [Google Scholar] [CrossRef]
- Tapia–Torres, Y.; Elser, J.J.; Souza, V.; García–Oliva, F. Ecoenzymatic stoichiometry at the extremes: How microbes cope in an ultra–oligotrophic desert soil. Soil Biol. Biochem. 2015, 87, 34–42. [Google Scholar] [CrossRef]
- Steinweg, J.M.; Dukes, J.S.; Wallenstein, M.D. Modeling the effects of temperature and moisture on soil enzyme activity: Linking laboratory assays to continuous field data. Soil Biol. Biochem. 2012, 55, 85–92. [Google Scholar] [CrossRef]
- Ding, F.; Ji, D.; Yan, K.; Dijkstra, F.A.; Bao, X.; Li, S.; Kuzyakov, Y.; Wang, J. Increased soil organic matter after 28 years of nitrogen fertilization only with plastic film mulching is controlled by maize root biomass. Sci. Total Environ. 2022, 810, 152244. [Google Scholar] [CrossRef]
- Lee, J.G.; Chae, H.G.; Hwang, H.Y.; Kim, P.J.; Cho, S.R. Effect of plastic film mulching on maize productivity and nitrogen use efficiency under organic farming in South Korea. Sci. Total Environ. 2021, 787, 147503. [Google Scholar] [CrossRef]
- Donald, R.Z.; William, E.H.; Andrew, J.B.; Kurt, S.P.; Alan, F.T. Simulated atmospheric NO3− deposition increases soil organic matter by slowing decomposition. Ecol. Appl. 2008, 18, 2016–2027. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wu, L.; Zhao, L.; Lu, X.; Fan, Q.; Zhang, F. Influence of continuous plastic film mulching on yield, water use efficiency and soil properties of rice fields under non–flooding condition. Soil Tillage Res. 2007, 93, 370–378. [Google Scholar] [CrossRef]
- Liu, C.; Jin, S.; Zhou, L.; Jia, Y.; Li, F.; Xiong, Y.; Li, X. Effects of plastic film mulch and tillage on maize productivity and soil parameters. Eur. J. Agron. 2009, 31, 241–249. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Zhao, J.; Zhang, W.; Xiao, K.; Wang, K. Nitrogen addition aggravates microbial carbon limitation: Evidence from ecoenzymatic stoichiometry. Geoderma 2018, 329, 61–64. [Google Scholar] [CrossRef]
- Dong, W.; Si, P.; Liu, E.; Yan, C.; Zhang, Z.; Zhang, Y. Influence of film mulching on soil microbial community in a rainfed region of northeastern China. Sci. Rep. 2017, 7, 8468. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.G.; Chae, H.G.; Das, S.; Kim, G.W.; Kim, P.J. Cover crops offset recalcitrant soil organic carbon losses under plastic–film mulching by altering microbial functional genes. Biol. Fertil. Soils 2022. [Google Scholar] [CrossRef]
- Muñoz–Romero, V.; Benítez–Vega, J.; López–Bellido, R.J.; Fontán, J.M.; López–Bellido, L. Effect of tillage system on the root growth of spring wheat. Plant Soil 2009, 326, 97–107. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.; Li, J.; Zhou, X.; Cao, J.; Wang, R.; Wang, Y.; Shelton, S.; Jin, Z.; Walker, L.M.; et al. Costimulation of soil glycosidase activity and soil respiration by nitrogen addition. Glob. Chang. Biol. 2017, 23, 1328–1337. [Google Scholar] [CrossRef]
- Jing, X.; Chen, X.; Tang, M.; Ding, Z.; Jiang, L.; Li, P.; Ma, S.; Tian, D.; Xu, L.; Zhu, J.; et al. Nitrogen deposition has minor effect on soil extracellular enzyme activities in six Chinese forests. Sci. Total Environ. 2017, 607–608, 806–815. [Google Scholar] [CrossRef]
- Zhou, L.; Feng, H. Plastic film mulching stimulates brace root emergence and soil nutrient absorption of maize in an arid environment. J. Sci. Food Agric. 2020, 100, 540–550. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Zheng, M.; Jiang, L.; Luo, Y. Patterns and mechanisms of responses by soil microbial communities to nitrogen addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Fatemi, F.R.; Fernandez, I.J.; Simon, K.S.; Dail, D.B. Nitrogen and phosphorus regulation of soil enzyme activities in acid forest soils. Soil Biol. Biochem. 2016, 98, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Ajwa, H.A.; Dell, C.J.; Rice, C.W. Changes in enzyme activities and microbial biomass of tallgrass prairie soil as related to burning and nitrogen fertilization. Soil Biol. Biochem. 1999, 31, 769–777. [Google Scholar] [CrossRef]
- Cusack, D.F.; Silver, W.L.; Torn, M.S.; Burton, S.D.; Firestone, M.K. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology 2011, 92, 621–632. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Yu, S.; Zhang, L.; Dong, K.; Feng, B. Mulching practices manipulate the microbial community diversity and network of root-associated compartments in the Loess Plateau. Soil Tillage Res. 2022, 223, 105476. [Google Scholar] [CrossRef]
- Olander, L.P.; Vitousek, P.M. Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 2000, 49, 175–191. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Guo, X.; Wang, X.; Zhang, Y.; Li, P.; Zhang, X. Ecoenzymatic stoichiometry and microbial nutrient limitation in rhizosphere soil in the arid area of the northern Loess Plateau, China. Soil Biol. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Perroni, Y.; García–Oliva, F.; Tapia–Torres, Y.; Souza, V. Relationship between soil P fractions and microbial biomass in an oligotrophic grassland–desert scrub system. Ecol. Res. 2014, 29, 463–472. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, X.; Zheng, B.; Yue, S.; Zhang, X.; Zhai, B.; Wang, Z.; Zheng, W.; Li, Z.; Zamanian, K.; et al. Effects of plastic and straw mulching on soil microbial P limitations in maize fields: Dependency on soil organic carbon demonstrated by ecoenzymatic stoichiometry. Geoderma 2021, 388, 114928. [Google Scholar] [CrossRef]
- Xu, H.; Qu, Q.; Chen, Y.; Liu, G.; Xue, S. Responses of soil enzyme activity and soil organic carbon stability over time after cropland abandonment in different vegetation zones of the Loess Plateau of China. Catena 2021, 196, 104812. [Google Scholar] [CrossRef]
- Saiya–Corka, K.R.; Sinsabaugha, R.L.; Zakb, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0–10 cm | 10–20 cm | 20–30 cm | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | SOC | TN | TP | pH | SOC | TN | TP | pH | SOC | TN | TP | |
| N | <0.01 | <0.01 | <0.05 | <0.01 | =0.09 | <0.01 | <0.01 | <0.01 | <0.05 | <0.05 | <0.05 | <0.01 |
| P | <0.01 | =0.12 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.17 | <0.01 | <0.01 | <0.01 |
| N*P | <0.01 | =0.13 | =0.15 | =0.43 | <0.01 | <0.01 | =0.35 | =0.33 | <0.01 | =0.94 | =0.22 | =0.08 |
| 0–10 cm | 10–20 cm | 20–30 cm | |||||||
|---|---|---|---|---|---|---|---|---|---|
| βG | NAG + LAP | AP | βG | NAG + LAP | AP | βG | NAG + LAP | AP | |
| N | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.05 | <0.01 | <0.05 |
| P | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| N*P | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.05 | <0.01 | <0.01 |
| 0–10 cm | 10–20 cm | 20–30 cm | |||||||
|---|---|---|---|---|---|---|---|---|---|
| EC:N | EC:P | EN:P | EC:N | EC:P | EN:P | EC:N | EC:P | EN:P | |
| N | <0.01 | <0.01 | <0.01 | <0.05 | <0.05 | <0.05 | <0.01 | <0.05 | <0.01 |
| P | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| N*P | <0.05 | <0.05 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Wang, M.; Sun, C.; Wu, H.; Zhao, X.; Liu, E.; Dong, W.; Yan, M. Self-Regulation of Soil Enzyme Activity and Stoichiometry under Nitrogen Addition and Plastic Film Mulching in the Loess Plateau Area, Northwest China. Agriculture 2023, 13, 938. https://doi.org/10.3390/agriculture13050938
Liu M, Wang M, Sun C, Wu H, Zhao X, Liu E, Dong W, Yan M. Self-Regulation of Soil Enzyme Activity and Stoichiometry under Nitrogen Addition and Plastic Film Mulching in the Loess Plateau Area, Northwest China. Agriculture. 2023; 13(5):938. https://doi.org/10.3390/agriculture13050938
Chicago/Turabian StyleLiu, Meixia, Menglu Wang, Congwei Sun, Hui Wu, Xueqing Zhao, Enke Liu, Wenyi Dong, and Meiling Yan. 2023. "Self-Regulation of Soil Enzyme Activity and Stoichiometry under Nitrogen Addition and Plastic Film Mulching in the Loess Plateau Area, Northwest China" Agriculture 13, no. 5: 938. https://doi.org/10.3390/agriculture13050938