1. Introduction
As the world’s population grows, so will global food production, putting additional pressure on already-scarce resources such as clean irrigation water and arable land [
1,
2]. In addition, increasing soil salinity and dry conditions caused by climate change are regarded as the most critical and adverse environmental factors for plants, leading to enormous losses in plant production worldwide [
3]. Thus, it is of primary importance to study plant responses to salinity and drought so that the optimal conditions for plant production can be well understood.
Plants in their natural habitat are concurrently exposed to various environmental conditions, such as salinity, drought, and extreme temperatures, during their growing stages [
4,
5]. However, numerous studies conducted have focused on the individual effects of drought and salinity, whereas their interaction has not been taken into much consideration [
6,
7]. According to several studies on commercial crops, water deficit worsens the adverse effects of salinity by interfering with photosynthesis and nutrient uptake, which further inhibits growth [
8,
9]. Conversely, research on halophytic grass (
Panicum antidotale) showed that the negative effects of drought alone on plant growth and photosynthesis could be mitigated by the combination of low salinity and drought [
10]. Likewise, Alam et al. [
11] reported that the combined effect of drought and salinity did not show any significant effects on the shoot length of
Salsola imbricata (Fetid Saltwort). This then suggests that edible halophytes could be a suitable solution in saline areas with low rainfall. Moreover, agricultural land lost due to salinity would be regained to produce staple food.
Mesembryanthemum crystallinum L., also known as glacier lettuce or ice plant, is an edible annual succulent belonging to the Aizoaceae family [
12]. It is native to the southern coastal regions of Africa and is widely distributed along the coastal areas of Europe, the USA, Mexico, Chile, the Caribbean, and western Australia [
13,
14,
15]. The species is already consumed as a leafy vegetable in several countries, such as New Zealand, India, Germany, and the Netherlands [
13]. The medicinal value of the leaf extract of the species has been reported in Tunisian folkloric medical treatments for ocular infections and as a remedy for throat and mouth infections [
14,
16]. The juice from the leaves is also used to relieve lung irritation, water retention, painful urination, and inflammation of the respiratory system. Thus, the species was classified as a highly functional food [
17].
Moreover, the ice plant is regarded as a salt- and drought-tolerant species, and numerous laboratory experiments elucidating the physiological and molecular mechanisms behind the individual effects of salinity and drought have been published [
13,
18,
19,
20]. Yet, the combined effect of these stress factors on
M. crystallinum remains unknown, as it has not been subject to much research, especially on the relative yields of minerals, phytochemicals, antioxidants, and proximate substances under different salinity levels and irrigation intervals. In addition, there is a dearth of information on the nutritional benefits of the ice plant and its potential use as a suitable vegetable for marginal areas. Thus, there is a need to study the combined effect of these stresses to support its cultivation in water-scarce regions that are affected by salinity. Therefore, this study assessed the combined effect of salinity and irrigation-interval-induced water stress on the plant growth, minerals, and proximate and phytochemical contents of
M. crystallinum. The findings are expected to be useful in the domestication of this species in southern Africa, where water scarcity and salinity are prevalent.
4. Discussion
Drought and salinity are known to be the most common coexisting factors affecting crop yield and productivity [
29]. Earlier reports have stated that the combined effects of drought and salinity are more detrimental than the individual effects of each stress on plant growth and yield [
30,
31]. Although the patterns of different plants’ responses to salinity and drought are similar, their thresholds to stress vary from one species to another. In
M. crystallinum, the individual effects of drought and salinity have been extensively studied, and the results have shown that the plant is drought- and salt-tolerant [
17,
32]. However, their combined effects have not yet been subject to much research. In this study, the individual and combined effects of salt and drought stress were evaluated on common ice plants. Results have shown that plants irrigated every two days without salinity had a longer leaf length. However, this was comparable to that of plants subjected to salinity and drought, suggesting that the combined effects on leaf length were not significant. These findings concur with the results reported by Alam et al. [
11] on
Salsola imbricata (Fetid Saltwort), where the combined effect of salt and water stress on shoot length was not significant. However, these results contradict the findings of Calone et al. [
33] on
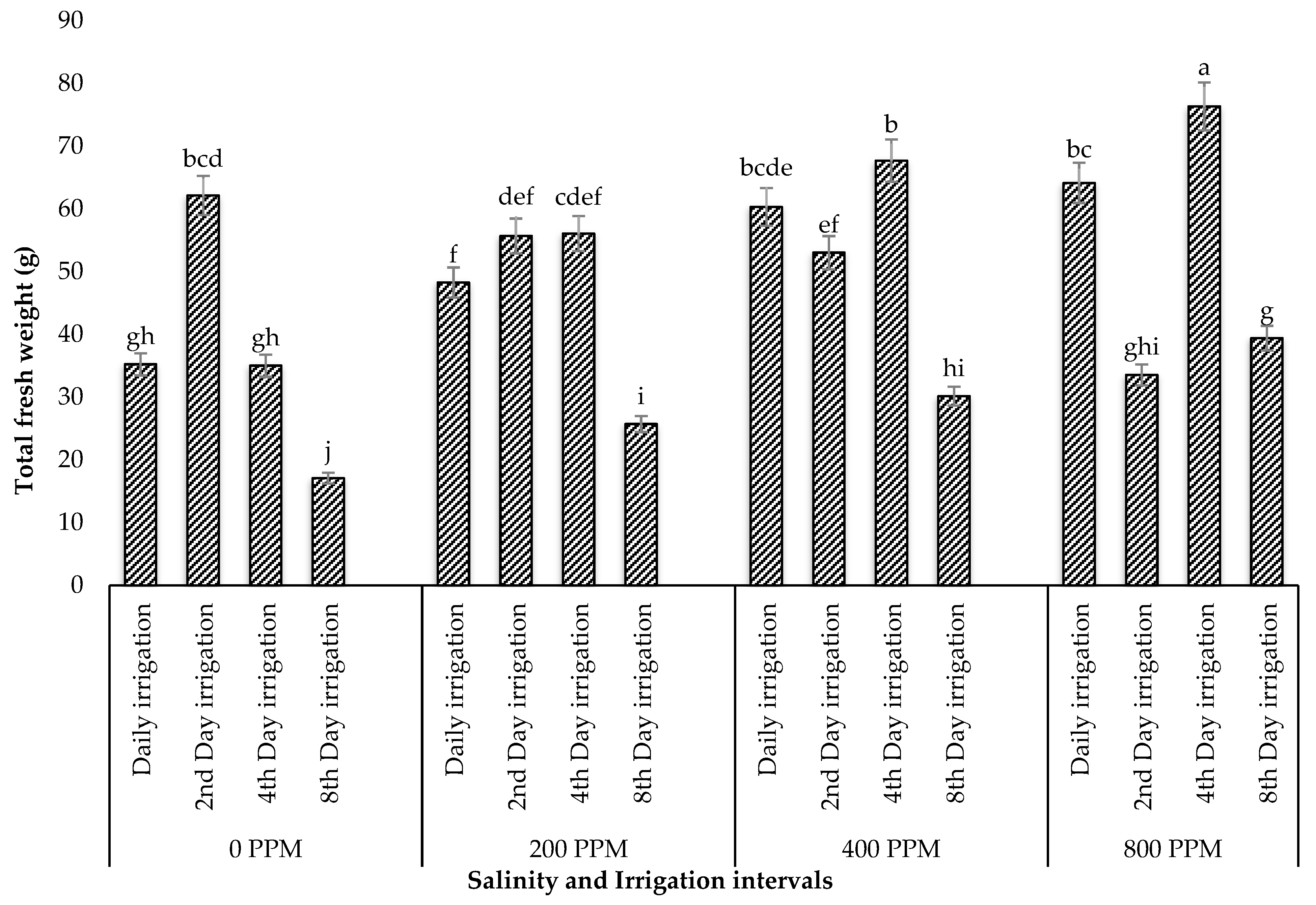
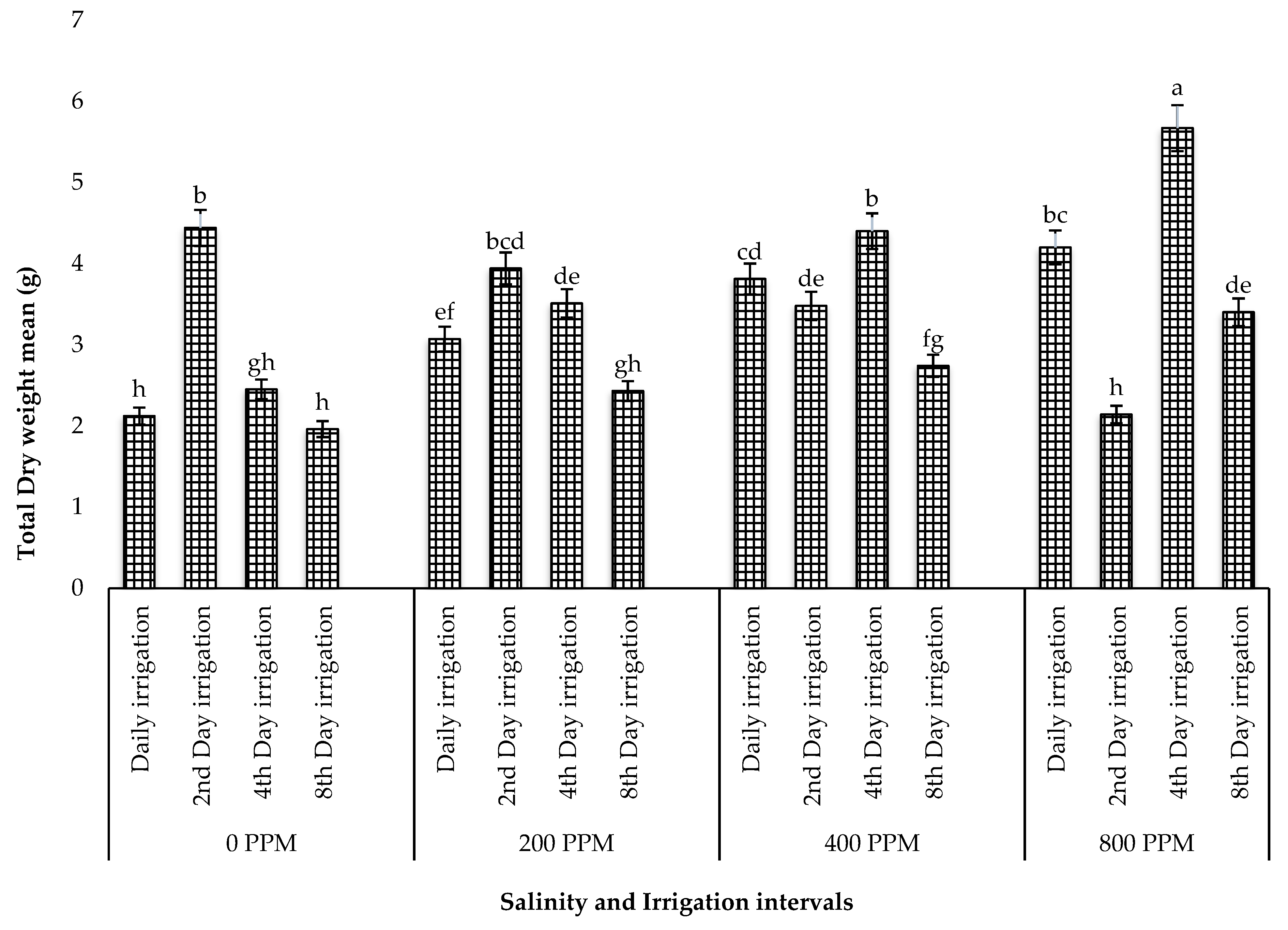
Limonium angustebracteatum, where the individual effect of salt stress did not have a negative effect on leaf length but was remarkably affected by both factors. This suggests that the combined tolerance to drought and salinity could be species-specific among halophytes. When assessing the number of leaves among treatments, plants irrigated every four days with 800 ppm salinity had the highest number of leaves. This increase also resulted in higher total fresh and dry weights. These results support the findings of Sogoni et al. [
24], who observed a significant increase in the leaf/branch number and total fresh and dry weights in
Tetragonia decumbens subjected to salinity and drought. This behaviour can be explained by the availability of epidermal bladder cells in the leaves of many halophytes, including the common ice plant [
17]. These bladder cells are known to have water and NaCl storage functions for osmotic adjustments in the vacuole, which enables these species to dilute excessive salt and enhance plant growth [
17].
Drought and salt stress have been reported as major factors responsible for senescence mechanisms in plants, which cause a reduction in the chlorophyll content of many plant species [
34]. Nevertheless, the effect of these abiotic factors on chlorophyll reduction varies from species to species [
35]. In the present study, leaf chlorophyll contents among tested treatments were comparable to that of the control during the growing weeks and were only reduced in the last week before harvest. This reduction in chlorophyll content did not have any negative effects on plant growth. These findings were also observed by Atzori et al. [
2] in a field experiment, where the common ice plant was not negatively affected by the reduction in the photosynthetic apparatus as the plant aged under increased seawater irrigation. These findings confirm that the species can tolerate both salt and drought stress under cultivation.
The nutritional quality of commercial crops around the world has been heavily affected by salinity and drought conditions, with a more pronounced effect in arid regions [
36]. This has triggered an increasing global interest in the investigation of the nutritional and nutraceutical value of halophytes to tackle malnutrition and increase food security in countries affected by drought and salinity [
37]. Halophytes have been proven to possess important healthy minerals with several phytochemical compounds essential for human consumption [
38,
39]. In the present study, high yields of minerals present in the leaves of the common ice plant under salinity and drought were found to be comparable to those in other edible halophytes, such as
Aster tripolium,
Sarcocornia perennis,
Salicornia ramosissima, and
Arthrocnemum macrostachyum, which are already consumed and sold in restaurants and supermarkets in other parts of the world [
34,
38,
40]. The variation observed in the results shows that drought and salinity modulate mineral composition, since most minerals tested were higher than the recommended dietary intake allowance (RDA). For instance, 2000 mg of potassium is required for an adult, a limit that was obtained in all tested treatments, including the control. When compared to the previous literature on other leafy vegetables subjected to salinity and drought, such as New Zealand spinach, water spinach,
Salicornia, and
Sarcocornia, the common ice plant was shown to have a high content of potassium [
1,
41,
42]. This mineral is the most prevalent intracellular cation and plays a crucial role in excitable tissues such as the heart, neurons, and skeletal muscles, as it is essential for action potentials and electrical excitability [
43]. Therefore, the daily consumption of the leaves of the common ice plant would be a good source of dietary potassium.
The composition of magnesium in the analysed leaves of all treatments ranged from 340 to 1135 mg/100 g, and these values are higher than the RDA of 55 mg/100 g, proving that this species is a rich source of magnesium. The values attained in this study are comparable to those recorded by Patricia et al. [
44] and Mih et al. [
45] on wild vegetables consumed by the people of Lebialem Highlands, Southwestern Cameroon, and Northern Cote D’Ivoire. Magnesium is well known for preventing a number of illnesses, such as cardiovascular disorders, and its deficiency is also linked to the aetiology of diabetes mellitus [
46,
47]. Additionally, it is needed in the human body as an intracellular electrolyte and as a co-factor for the creation of numerous enzymes, proteins, and nucleic acids [
48]. Thus, the consumption of this species will help in mitigating several illnesses.
When assessing the composition of calcium within the tested samples, most treatments meet the RDA of 1000 mg in respect of drought and salinity exposure. These results substantiate those obtained by Davis et al. [
49] on
Sambucus nigra exposed to varying environmental stresses, where the leaves and flowers had calcium that ranged from 500 to 1228 mg/100 g. The common ice plant has proven to be a good source of calcium, which has the ability to retain extracellular fluids, build bones and teeth, and transmit nerve impulses, blood clotting, and muscle contraction [
50,
51]. However, when examining the phosphorus composition within samples, all treatments fell short of the RDA of 700 mg. These results are in agreement with the findings of Jimoh et al. [
22] on
Amaranthus caudatus, where the examined samples had lower phosphorus than the RDA of 700 mg.
The accumulation of sodium (Na) in the leaves and roots of many halophytes in bio-saline agriculture has been reported to cause a decline in cations such as K
+, Ca
2+, and Mg
2+ [
42]. In this study, none of these cations declined, supporting the existence of a clear-cut nutrient absorption system that allows large NaCl compartmentation. This then enables the species to accumulate high Na content that is beyond the RDA of 200–500 mg proposed for healthy living organisms and might cause health problems. Caparrota et al. [
52] recommended the boiling technique, as it reduced the Na composition of spinach leaves cultivated under seawater irrigation. Thus, it will be vital to cook or boil ice plant leaves before consumption when cultivated under saline conditions.
A variety of micronutrients, including iron, zinc, aluminium, copper, and manganese, are crucial for human nutrition [
53]. However, their daily consumption is required in small quantities of not less than 20 mg, which accounts for less than 0.01% of body weight [
54]. The micronutrients accumulated in the leaves of the ice plant were below the recommended daily allowance of 20 mg, except iron, which was above the RDA of 20 mg in all treatments, including the control. Iron has been reported to be the most commonly deficient micronutrient in school children, and its deficiency has been implicated in anaemia, fatigue, and blood-related diseases [
55,
56]. Most people receive iron by eating vegetables, particularly spinach [
39]. Using spinach as a comparison, it is clear that ice plant leaves are a richer source of iron, displaying levels 5–10 times higher than those found in spinach.
When assessing the proximate composition of the leaves of the ice plant, variation among treatments were observed. Salinity and drought are known to impair plant nutrition; however, in this study, the ash content of the samples increased with increasing salinity and drought conditions. Ash has been used to measure the nutritional value of food and is believed to be an indicator of the mineral contents that have been conserved in food items. The ash content of the tested samples ranged from 35 to 52%, which is higher than the 5% reported for other wild vegetables and corresponds to the composition found in processed foods [
57]. These results concur with Ntuli’s [
53] research on two species of water spinach (
Ipomea plebeian R.Br. and
Ipomea wightii (Wall.) Choisy), where the ash level was reported to range from 20 to 38%. The plant’s high ash value suggests that it is an abundant source of dietary fibre.
Most wild vegetables, including halophytes, have been reported to have low levels of unsaturated fats that range from 2 to 4% [
21]. The amount of crude fat in the analysed samples concurs with this finding of [
21] since it was lower than 3% in all tested treatments, including the control. Excessive fat in food can lead to increased cholesterol, which is a major cause of cardiovascular disorders. Thus, the consumption of this leafy vegetable will be suitable for the management of weight loss and diseases caused by excess fat content. When assessing the protein content within samples, it ranged from 9 to 15%. This is similar to what was reported by Ajayi et al. [
58] on
Amaranthus cruentus (11.32%) and
Solanum nigrum (15.06%). This implies that the ice plant can be a good natural source of protein and could reduce protein malnutrition in children, especially in developing nations.
Dietary fibre is essential for controlling bowel movements, preventing cardiovascular disease, and slowing the absorption of cholesterol. The neutral detergent fibre (NDF) present in the tested samples ranged from 20 to 29% and was higher than the values reported in other wild vegetables, such as
Amaranthus cruentus (8.45%),
Celosia argentea (23%), and
Solanum nigrum (9.56%) [
55,
58]. The NDF concentration found in ice plant leaves is a sign that the species may assist in regulating intestinal transit, increasing dietary bulk, and lowering the risk of various metabolic illnesses such as colon cancer, obesity, and diabetes that are brought on by the insufficient intake of crude fibre. Moreover, the tested samples possessed a lower moisture content that ranged from 7 to 10%, suggesting that the leaves of this species might have lower microbial contamination and chemical degradation, which are normally associated with high moisture content [
59]. These lower values imply that the leaves of the ice plant may have a lengthy storage life, benefiting producers and sellers.
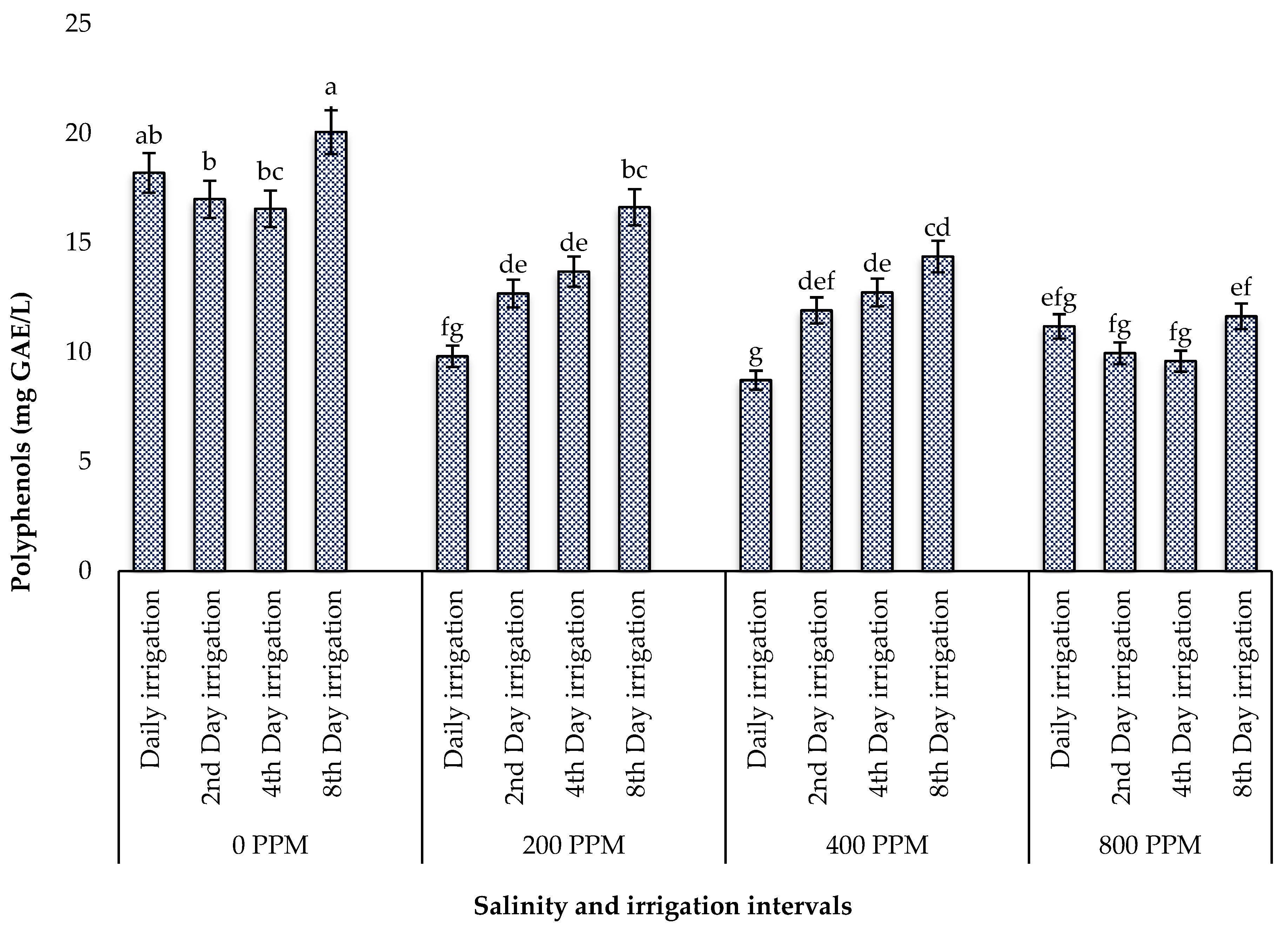
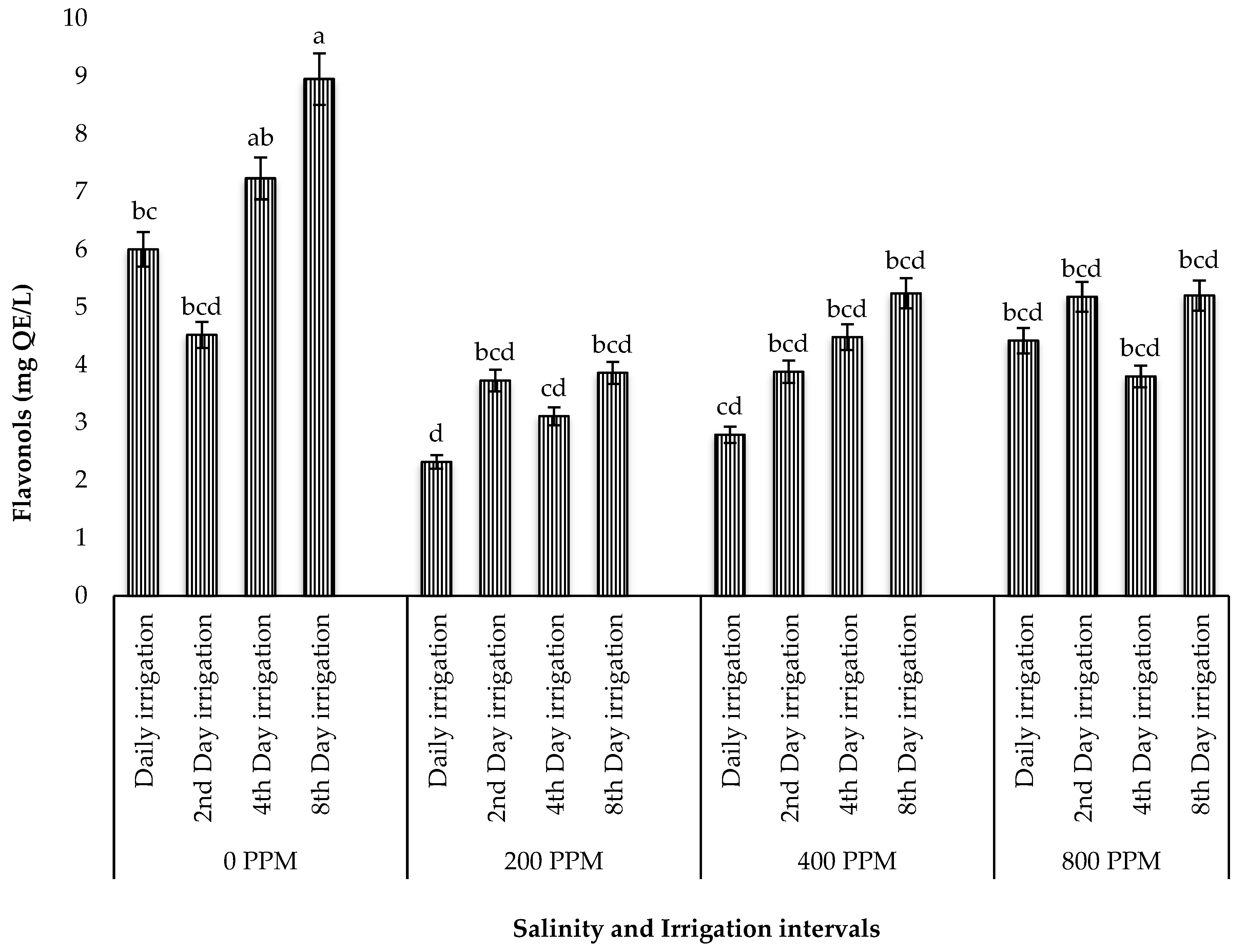
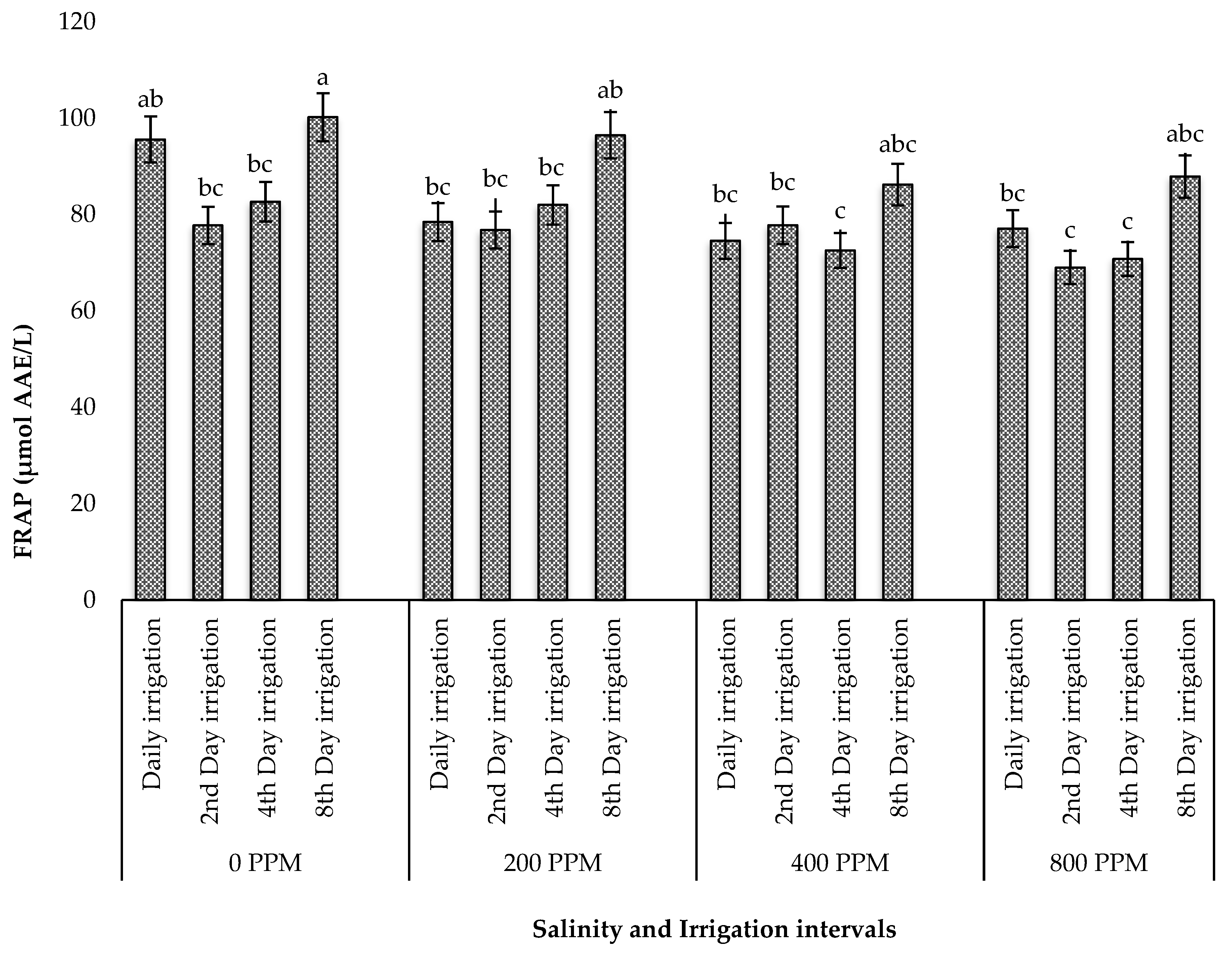
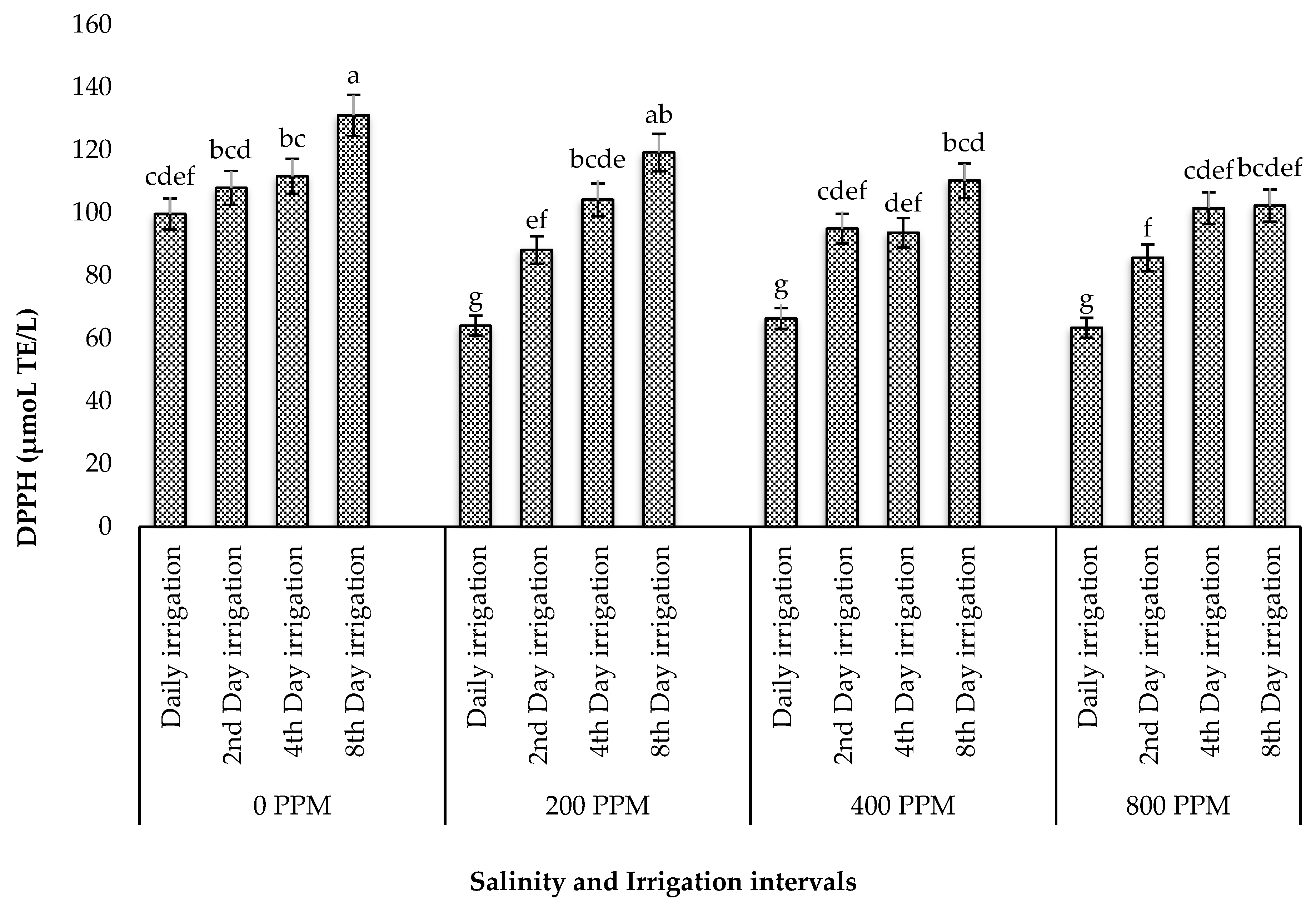
Abiotic factors such as salinity and drought stress are known to increase the production of reactive oxygen species (ROS), triggering oxidative stress and the activation of antioxidant mechanisms in plants [
60]. During this process, many plant species alter their growth and produce metabolites, such as phenolic compounds, which act as reducing agents, hydrogen donors, and singlet oxygen quenchers [
61]. These phenolic compounds are of great intrinsic importance in human nutrition since they scavenge free radicals and suppress lipid peroxidation in human tissues, which prevents potential issues brought on by the excessive consumption of synthetic additives. In the present study, the individual effect of drought stress optimized the yield of polyphenols and flavonols, while its interaction with salinity reduced these compounds. These findings contradict those of Alam et al. [
11] on
Salsola imbricata (Fetid Saltwort) subjected to both salinity and drought, where an increase in proline accumulation acted as an antioxidative defence system to maintain a balance between ROS over-accumulation and their elimination to keep ROS at the signalling level required for plant growth [
62]. The positive effect of salinity in drought-stressed halophytes has also been reported in
Atriplex halimus L., where the accumulation of antioxidants was reduced in samples subjected to the combined effect of salinity and drought [
63]. This improved drought tolerance under salinity may be due to osmotic adjustment through higher Na
+ and proline accumulation and antioxidative enzymes [
64]. Nevertheless, the leaves of ice plants subjected to both salinity and water stress possessed more polyphenols and flavonols than other promising edible halophytes in South Africa, such as
Chenopodium album,
Trachyandra divaricata,
Trachyandra ciliata [
21,
65,
66,
67], and
Tetragonia decumbens [
24,
68]. This suggests that the leaves of this plant may be a good source of nutritional antioxidants.
In summary, increased salt concentrations did not influence the high yields of acid detergent fibre (ADF), crude fat, protein, neutral detergent fibre (NDF), and phytochemicals and antioxidants in M. crystallinum, as these phytonutrients were recorded in high amounts in plants subjected to irrigation intervals only, although a combination of salinity and irrigation intervals resulted in the highest ash and moisture contents. These findings validate earlier reports that M. crystallinum can excrete salt through its foliar epidermal bladder, thereby normalizing salt-induced stress, which could trigger the increased accumulation of phytochemicals, antioxidants, and proximate nutrients in the species.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}