
Use of Gas Chromatography and SPME Extraction for the Differentiation between Healthy and Paenibacillus larvae Infected Colonies of Bee Brood—Preliminary Research
 , ,
, ,
Abstract
:
1. Introduction
2. Material and Methods
2.1. Sampling
2.2. Extraction and Chromatographic Analysis of VOCs Emitted by Brood
2.3. Statistical Analysis of the Obtained Results
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alford, D.V. A study of the hibernation of Bumblebees (Hymenoptera: Bombidae) in Southern England. J. Anim. Ecol. 1969, 38, 149–170. [Google Scholar] [CrossRef]
- Richards, O.W. The biology of the social wasps (Hymenoptera, Vespidae). Biol. Rev. 1971, 46, 483–528. [Google Scholar] [CrossRef]
- Herrera, C.M. Floral biology, microclimate, and pollination by ectothermic bees in an early-blooming herb. Ecology 1995, 76, 218–228. [Google Scholar] [CrossRef]
- Wahengbam, J.; Raut, A.M.; Satinder, P.; Banu, A.N. Role of bumble bee in pollination. Ann. Biol. 2019, 35, 290–295. [Google Scholar]
- Topolska, G.; Gajda, A.; Imińska, U. Atlas of Bee Diseases; PWRiL: Warsaw, Poland, 2018. (In Polish) [Google Scholar]
- Genersch, E. Paenibacillus larvae and American Foulbrood—long since known and still surprising. J. Verbr. Lebensm. 2008, 3, 429–434. [Google Scholar] [CrossRef]
- Schulz, M.; Łoś, A.; Skowronek, P.; Staniszewski, A.; Iwański, B.; Wójcik, Ł.; Strachecka, A. American foulbrood—A dangerous bacterial honey bee disease. J. Anim. Sci. Biol. Bioecon. 2018, 36, 35–47. [Google Scholar]
- Ebeling, J.; Knispel, H.; Hertlein, G.; Fünfhaus, A.; Genersch, E. Biology of Paenibacillus larvae, a deadly pathogen of honey bee larvae. Appl. Microbiol. Biotechnol. 2016, 100, 7387–7395. [Google Scholar] [CrossRef]
- No 37/2010; Commission Regulation (EU) No 37/2010 of 22 December 2009 on pharmacologically active substances and their classification regarding maximum residue limits in foodstuffs of animal origin (Text with EEA relevance). Commission Regulation (EU): Brussels, Belgium, 2009.
- Lindström, A.; Korpela, S.; Fries, I. The distribution of Paenibacillus larvae spores in adult bees and honey and larval mortality, following the addition of American foulbrood diseased brood or spore-contaminated honey in honey bee (Apis mellifera) colonies. J. Invertebr. Pathol. 2008, 99, 82–86. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, E.; Müller, S.; Ensle, P.; Süssmuth, R.D.; Genersch, E. Elucidation of sevadicin, a novel nonribosomal peptide secondary metabolite produced by the honey bee pathogenic bacterium Paenibacillus larvae. Environ. Microbiol. 2014, 16, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Sood, S.; Steinmetz, H.; Beims, B.; Mohr, K.I.; Stadler, M.; Djukic, M.; von der Ohe, W.; Steinert, M.; Daniel, R.; Müller, R. Panilarvins: Iturin family lipopeptides from the honey bee pathogen Paenibacillus larvae. Chem. BioChem. 2014, 15, 1947–1955. [Google Scholar]
- Müller, S.; Garcia-Gonzalez, E.; Genersch, E.; Süssmuth, R. Involvement of secondary metabolites in the pathogenesis of the American foulbrood of honey bees caused by Paenibacillus larvae. Nat. Prod. Rep. 2015, 32, 765–778. [Google Scholar] [CrossRef] [Green Version]
- Gliński, Z.; Jarosz, J. Does Bacillus larvae produce an antibacterial substance in infected honey bee larvae? Apidologie 1992, 23, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gonzalez, E.; Poppinga, L.; Fünfhaus, A.; Hertlein, G.; Hedtke, K.; Jakubowska, A.; Genersch, E. Paenibacillus larvae chitindegrading protein PlCBP49 is a key virulence factor in American foulbrood of honey bees. PLoS Pathog. 2014, 10, e1004284. [Google Scholar] [CrossRef]
- Holst, E.C. A simple field test for American foulbrood. Amer. Bee 1946, 86, 34. [Google Scholar]
- Holst, E.C.; Sturtevant, A.P. Relation of proteolytic enzymes to phase of life cycle of Bacillus larvae, and two new culture media for this organism. J. Bacteriol. 1940, 40, 723–731. [Google Scholar] [CrossRef] [Green Version]
- Yue, D.; Nordhoff, M.; Wieler, L.H.; Genersch, E. Fluorescence in situ hybridization (FISH) analysis of the interactions between honeybee larvae and Paenibacillus larvae; the causative agent of American foulbrood of honeybees (Apis mellifera). Environ. Microbiol. 2008, 10, 1612–1620. [Google Scholar] [CrossRef]
- Bailey, L.; Ball, B.V. Honey Bee Pathology; Academic Press: New York, NY, USA, 1991. [Google Scholar]
- Genersch, E. American foulbrood in honeybees and its causative agent, Paenibacillus larvae. J. Invertebr. Pathol. 2010, 103, 10–19. [Google Scholar] [CrossRef]
- Ashiralieva, A.; Genersch, E. Reclassification; genotypes; and virulence of Paenibacillus larvae; the etiological agent of American foulbrood in honeybees—A review. Apidologie 2006, 37, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Sherman, P.W.; Seeley, T.D.; Reeve, H.K. Parasites, pathogens, and polyandry in social Hymenoptera. Am. Nat. 1988, 131, 602–610. [Google Scholar] [CrossRef]
- Palmer, K.A.; Oldroyd, B.P. Evidence for intra-colonial genetic variance in resistance to American foulbrood of honey bees (Apis mellifera): Further support for the parasite/pathogen hypothesis for the evolution of polyandry. Naturwissenschaften 2003, 90, 265–268. [Google Scholar]
- Alonso-Salces, R.M.; Cugnata, N.M.; Guaspari, E.; Pellegrini, M.; Aubone, I.; De Piano, F.G.; Antúnez, K.; Fuselli, S.R. Natural strategies for the control of Paenibacillus larvae; the causative agent of American foulbrood in honey bees: A review. Apidologie 2017, 48, 387–400. [Google Scholar] [CrossRef]
- Dostálková, S.; Urajová, P.; Činčárová, D.; Vránová, T.; Hrouzek, P.; Petřivalský, M.; Lukeš, M.; Čapková-Frydrychová, R.; Danihlík, J. Fatty acids and their derivatives from Chlorella vulgaris extracts exhibit in vitro antimicrobial activity against the honey bee pathogen Paenibacillus larvae. J. Apic. Res. 2021, 1–13. [Google Scholar] [CrossRef]
- Tutun, H.; Koç, N.; Kart, A. Plant essential oils used against some bee diseases. Turk. J. Agric. Food Sci. Technol. 2018, 6, 34–45. [Google Scholar] [CrossRef] [Green Version]
- St Clair, A.L.; Zhang, G.; Dolezal, A.G.; O’Neal, M.E.; Toth, A.L. Diversified farming in a monoculture landscape: Effects on honey bee health and wild bee communities. Environ. Entomol. 2020, 49, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Girard, M.; Chagnon, M.; Fournier, V. Pollen diversity collected by honey bees in the vicinity of Vaccinium spp. crops and its importance for colony development. Botany 2012, 90, 545–555. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Kobylińska, M. Regional diversity of beekeeping in Poland. Pol. Stat. 2021, 66, 25–38. (In Polish) [Google Scholar]
- Genersch, E.; Ashiralieva, A.; Fries, I. Strain-and genotype-specific differences in virulence of Paenibacillus larvae subsp. larvae, a bacterial pathogen causing American foulbrood disease in honeybees. Appl. Environ. Microbiol. 2005, 71, 7551–7555. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Lim, S.; Choi, Y.S.; Lee, M.L.; Kwon, H.W. Volatile disease markers of American foulbrood-infected larvae in Apis mellifera. J. Insect Physiol. 2020, 122, 104040. [Google Scholar] [CrossRef]
- Gochnauer, T.A.; Shearer, D.A. Volatile acids from honeybee larvae infected with Bacillus larvae and from a culture of the organism. J. Apicul. Res. 1981, 20, 104–109. [Google Scholar] [CrossRef]
- Bikaun, J.M.; Bates, T.; Bollen, M.; Flematti, G.R.; Melonek, J.; Praveen, P.; Grassl, J. Volatile biomarkers for non-invasive detection of American foulbrood, a threat to honey bee pollination services. Sci. Total. Environ. 2022, 845, 157123. [Google Scholar] [CrossRef] [PubMed]
- Liolios, V.; Kanelis, D.; Tananaki, C.; Rodopoulou, M.A. A comparative study of healthy and american foulbrood-infected bee brood (Apis mellifera L.) through the investigation of volatile compounds. Agriculture 2022, 12, 812. [Google Scholar] [CrossRef]
- Google Maps. Available online: www.google.pl/maps/ (accessed on 20 October 2022).
- Apiculture Factsheet. Ministry of Agriculture, Food and Fisheries. Available online: https://www2.gov.bc.ca/gov/content/industry/agriculture-seafood/animals-and-crops/animal-production/bees (accessed on 20 January 2023).
- Central Statistical Office; The Polish Statistician. 2021. Available online: https://ws.stat.gov.pl/WS/2021/2/gus_ws_2021_02.pdf.7 (accessed on 11 November 2022).
- Instruction of the Chief Veterinary Officer. Instruction of the Chief Veterinary Officer No. GIWpr 02010-23/2016; Publishing House of the Chief Veterinary Inspectorate: Warsaw, Poland, 2016. (In Polish) [Google Scholar]
- Hansen, H.; Brodsgaard, C.J. American foulbrood: A review of its biology; diagnosis and control. Bee World 1999, 80, 195–201. [Google Scholar] [CrossRef]
- Beekman, M.; Ratnieks, F. Long-range foraging by the honey bee; Apis mellifera L. Funct. Ecol. 2000, 14, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Crailsheim, K.; Stolberg, E. Influence of diet: Age and colony condition upon intestinal proteolytic activity and size of the hypopharyngeal glands in the honeybee (Apis mellifera L.). J. Insect Physiol. 1989, 35, 595–602. [Google Scholar] [CrossRef]
- Omar, E.; Abd-Ella, A.A.; Khodairy, M.M.; Moosbeckhofer, R.; Crailsheim, K.; Brodschneider, R. Influence of different pollen diets on the development of hypopharyngeal glands and size of acid gland sacs in caged honey bees (Apis mellifera). Apidologie 2017, 48, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Barascou, L.; Sene, D.; Barraud, A.; Michez, D.; Lefebvre, V.; Medrzycki, P.; Di Prisco, G.; Strobl, V.; Yañez, O.; Neumann, P.; et al. Pollen nutrition fosters honeybee tolerance to pesticides. R. Soc. Open Sci. 2021, 8, 210818. [Google Scholar] [CrossRef]
- Lan, J.; Ding, G.; Ma, W.; Jiang, Y.; Huang, J. Pollen source affects development and behavioral preferences in honey bees. Insects 2021, 12, 130. [Google Scholar] [CrossRef]
- Piana, M.L.; Paola, B.; Livia, P.O.; Sara, P. Pollen analysis of royal jelly: Contribution to analytical methods and characterization. Apiacta 2006, 41, 28–43. [Google Scholar]
- Ashour, Z.A.; Ali, M.A.M.; Abdelmegeed, S.M.; Amin, K.M. Detection of microorganisms (bacteria, fungi and yeasts) in royal jelly. Arab Univ. J. Agric. Sci. 2019, 26, 2533–2539. [Google Scholar] [CrossRef]
- Hroncova, Z.; Havlik, J.; Killer, J.; Doskocil, I.; Tyl, J.; Kamler, M.; Titera, D.; Hakl, J.; Mrazek, J.; Bunesova, V.; et al. Variation in honey bee gut microbial diversity affected by ontogenetic stage, age and geographic location. PLoS ONE 2015, 10, e0118707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slessor, K.N.; Winston, M.L.; Le Conte, Y. Pheromone communication in the honeybee (Apis mellifera L.). J. Chem. Ecol. 2005, 31, 2731–2745. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Mohammedi, A.; Robinson, G.E. Primer effects of a brood pheromone on honeybee behavioural development. Proc. R. Soc. Biol. Sci. 2001, 268, 163–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Conte, Y.; Arnold, G.; Trouiller, J.; Masson, C. Identification of a brood pheromone in honeybees. Naturwissenschaften 1990, 81, 462–465. [Google Scholar] [CrossRef]
- Wagoner, K.M.; Millar, J.G.; Schal, C.; Rueppell, O. Cuticular pheromones stimulate hygienic behavior in the honey bee (Apis mellifera). Sci. Rep. 2020, 10, 7132. [Google Scholar] [CrossRef]
- Mihai, C.M.; Mărghitaş, L.A.; Dezmirean, D.S.; Chirilă, F.; Moritz, R.F.A.; Schlüns, H. Interactions among flavonoids of propolis affect antibacterial activity against the honeybee pathogen Paenibacillus larvae. J. Invertebr. Pathol. 2012, 110, 68–72. [Google Scholar] [CrossRef]
- Wilson, M.B.; Brinkman, D.; Spivak, M.; Gardner, G.; Cohen, J.D. Regional variation in composition and antimicrobial activity of US propolis against Paenibacillus larvae and Ascosphaera apis. J. Invertebr. Pathol. 2015, 124, 44–50. [Google Scholar] [CrossRef]
- Jaouani, I.; Abbassi, M.S.; Alessandria, V.; Bouraoui, J.; Ben Salem, R.; Kilani, H.; Mansouri, R.; Messadi, L.; Cocolin, L. High inhibition of Paenibacillus larvae and Listeria monocytogenes by Enterococcus isolated from different sources in Tunisia and identification of their bacteriocin genes. Lett. Appl. Microbiol. 2014, 59, 17–25. [Google Scholar] [CrossRef]
- Kešnerová, L.; Emery, O.; Troilo, M.; Liberti, J.; Erkosar, B.; Engel, P. Gut microbiota structure differs between honeybees in winter and summer. ISME J. 2020, 14, 801–814. [Google Scholar] [CrossRef] [Green Version]
- Castelli, L.; Branchiccela, B.; Garrido, M.; Invernizzi, C.; Porrini, M.; Romero, H.; Santos, E.; Zunino, P.; Antúnez, K. Impact of nutritional stress on honeybee gut microbiota immunity and Nosema ceranae infection. Microb. Ecol. 2020, 80, 908–919. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Gonzaléz, M.J.; Porrini, M.P.; Eguaras, M.J.; Audisio, M.C.; Marioli, J.M. Synergistic effect of surfactin from Bacillus subtilis C4 and Achyrocline satureioides extracts on the viability of Paenibacillus larvae. World J. Microbiol. Biotechnol. 2012, 28, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Trytek, M.; Kowalski, P.; Fiedurek, J. Presence; biological role and biotransformation of limonene. Biotechnologia. 2003, 2, 206–217. (In Polish) [Google Scholar]
- Effmert, U.; Kalderás, J.; Warnke, R.; Piechulla, B. Volatile mediated interactions between bacteria and fungi in the soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Wahed, A.A.; Farag, M.A.; Eraqi, W.A.; Mersal, G.A.M.; Zhao, C.; Khalifa, S.A.M.; El-Seedi, H.R. Unravelling the beehive air volatiles profile as analysed via solid-phase microextraction (SPME) and chemometrics. J. King Saud Univ. Sci. 2021, 33, 101449. [Google Scholar] [CrossRef]
- Espina, L.; Gelaw, T.K.; de Lamo-Castellví, S.; Pagán, R.; García-Gonzalo, D. Mechanism of bacterial inactivation by (+)-Limonene and its potential use in food preservation combined processes. PLoS ONE 2013, 8, e56769. [Google Scholar] [CrossRef] [Green Version]
- Pașca, C.; Matei, J.A.; Diaconeasa, Z.; Rotaru, A.; Erler, S.; Dezmirean, D.S. Biologically active extracts from different medicinal plants tested as potential additives against bee pathogens. Antibiotics 2021, 10, 960. [Google Scholar] [CrossRef]
- Ansari, M.J.; Al-Ghamdi, A.; Usmani, S.; Al-Waili, N.; Nuru, A.; Sharma, D.; Khan, K.A.; Kaur, M.; Omer, M. In vitro evaluation of the effects of some plant essential oils on Paenibacillus larvae; the causative agent of American foulbrood. Biotechnol. Biotechnol. Equip. 2016, 30, 49–55. [Google Scholar] [CrossRef]
- Cavaleiro, C.; Pinto, E.; Gonçalves, M.J.; Salgueiro, L. Antifungal activity of Juniperus essential oils against dermatophyte: Aspergillus and Candida strains. J. Appl. Microbiol. 2006, 100, 1333–1338. [Google Scholar] [CrossRef]
- Valterová, I.; Vrkoč, J.; Norin, T. The enantiomeric composition of monoterpene hydrocarbons in the defensive secretions of Nasutitermes termites (Isoptera): Inter- and intraspecific variations. Chemoecology 1993, 4, 120–123. [Google Scholar] [CrossRef]
- Harborne, J.B. Preferencje Żywieniowe Owadów. In Ekologia Biochemiczna; Harborne, J.B., Ed.; PWN: Warszawa, Poland, 1997; pp. 154–188. (In Polish) [Google Scholar]
- da Silveira, S.M.; Luciano, F.B.; Fronza, N.; Cunha, A.; Scheuermann, G.N.; Vieira, C.R.W. Chemical composition and antibacterial activity of Laurus nobilis essential oil towards foodborne pathogens and its application in fresh Tuscan sausage stored at 7 °C. LWT Food Sci. Technol. 2014, 59, 86–93. [Google Scholar] [CrossRef]
- Merghni, A.; Marzouki, H.; Hentati, H.; Aouni, M.; Mastouri, M. Antibacterial and antibiofilm activities of Laurus nobilis L. essential oil against Staphylococcus aureus strains associated with oral infections. Curr. Res. Transl. Med. 2016, 64, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Yiin, L.M.; Millette, J.R.; Vette, A.; Ilacqua, V.; Quan, C.; Gorczynski, J.; Kendall, M.; Chen, L.C.; Weisel, C.P.; Buckley, B.; et al. Comparisons of the dust/smoke particulate that settled inside the surrounding buildings and outside on the streets of southern New York City after the collapse of the World Trade Center, September 11, 2001. J. Air Waste Manag. Assoc. 2004, 54, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Safara, S.; Harighi, B.; Bahramnejad, B.; Ahmadi, S. Antibacterial activity of endophytic bacteria against sugar beet root rot agent by volatile organic compound production and induction of systemic resistance. Front. Microbiol. 2022, 2, 921762. [Google Scholar] [CrossRef] [PubMed]
- Boontawan, A.; Boontawan, P. Isolation and characterization of Jatropha oil-degradation by Enterococcus faecalis and Burkholderia cenocepacia W-1 under anaerobic condition. Afr. J. Biotechnol. 2011, 10, 13841–13851. [Google Scholar]
- Spada, F.P.; de Alencar, S.M.; Purgatto, E. Comprehensive chocolate aroma characterization in beverages containing jackfruit seed flours and cocoa powder. Future Foods. 2022, 6, 100158. [Google Scholar] [CrossRef]
- El-Bastawesy, A.M.; Mohamed, R.H. Evaluation of cardamom oil role as antimicrobial anticarcenogenic and anti- inflammatory agents. Egypt. J. Agric. Res. 2005, 83, 789–809. [Google Scholar] [CrossRef]
- Nurraihana, H.; Norfarizan-Hanoon, N.A. Phytochemistry, pharmacology and toxicology properties of Strobilanthes crispus. Int. Food Res. J. 2013, 20, 2045–2056. [Google Scholar]
- Hailu, Y.M.; Atlabachew, M.; Chandravanshi, B.S.; Redi-Abshiro, M. Composition of essential oil and antioxidant activity of Khat (Catha edulis Forsk), Ethiopia. Chem. Int. 2017, 3, 25–31. [Google Scholar]
- El-Shaboury, G.; Haroun, S.; Shaker, K.; Badrorcid, A. Systematics implications of GC-MS analysis of secondary metabolites in the ethanol extract of Solanum species from South West Saudi Arabia. Egypt. J. Bot. 2017, 57, 429–444. [Google Scholar]
- Chowdhury, M.T.I.; Solaiman, A.H.M.; Zeba, N.; Hasanuzzaman, M.; Mustafiz, S.S.B.; Nahar, K. Determining the status of chlorinated pesticide residue in some leafy vegetables. J. Ecobiotechnol. 2009, 1, 41–45. [Google Scholar]
- Clewell, R.A.; Kremer, J.J.; Williams, C.C.; Campbell, J.L.; Andersen, M.E.; Borghoff, S.J. Tissue exposures to free and glucuronidated monobutylyphthalate in the pregnant and fetal rat following exposure to di-n-butylphthalate: Evaluation with a PBPK model. Toxicol. Sci. 2008, 103, 241–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omidpanah, S.; Saeidnia, S.; Saeedi, M.; Hadjiakhondi, A.; Manayi, A. Phthalate contamination of some plants and herbal products. Bol. Latinoam. Caribe Plantas Med. Aromat. 2018, 17, 61–67. [Google Scholar]
- Shaheed, K.A.; Al Garaawi, N.I.; Alsultany, A.K.; Abbas, Z.H.; Khshayyish, I.K.; Al Khazali, M.T. Analysis of bioactive phytochemical compound of (Cyperus iria L.) By using gas chromatography –mass spectrometry. IOP Conf. Ser. Earth Environ. Sci. 2019, 388, 012064. [Google Scholar] [CrossRef]
- Bhagat, A.P.; Bhuktar, A.S. Phytochemical and pharmacognostic investigation on Cardiospermum halicacabum L. (Sapindaceae). J. Pharmacogn. Phytochem. 2020, 9, 1691–1700. [Google Scholar]
- Joshi, U.P.; Wagh, R.D. GC-MS Analysis of phytochemical compounds present in the bark extracts of Ehretia laevis Roxb. Int. J. Res. Dev. Pharm. Life Sci. 2018, 7, 3150–3154. [Google Scholar] [CrossRef]
- Sharma, N.; Bahuguna, V.; Rawat, R.; Lalremruati, F.; Singh, A. Complete plant regeneration of Valeriana wallichii DC. on auxin enriched medium and phytochemical analysis. Plant Sci. Today 2020, 7, 542–550. [Google Scholar] [CrossRef]
- m-Phenylenediamne Compound Summary. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/m-Phenylenediamine. (accessed on 12 February 2023).
- Anderson, F.A. Final report on the safety assessment of m-phenylenediamine and m-phenylenediamine sulfate. Int. J. Toxicol. 1997, 16, 59–115. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
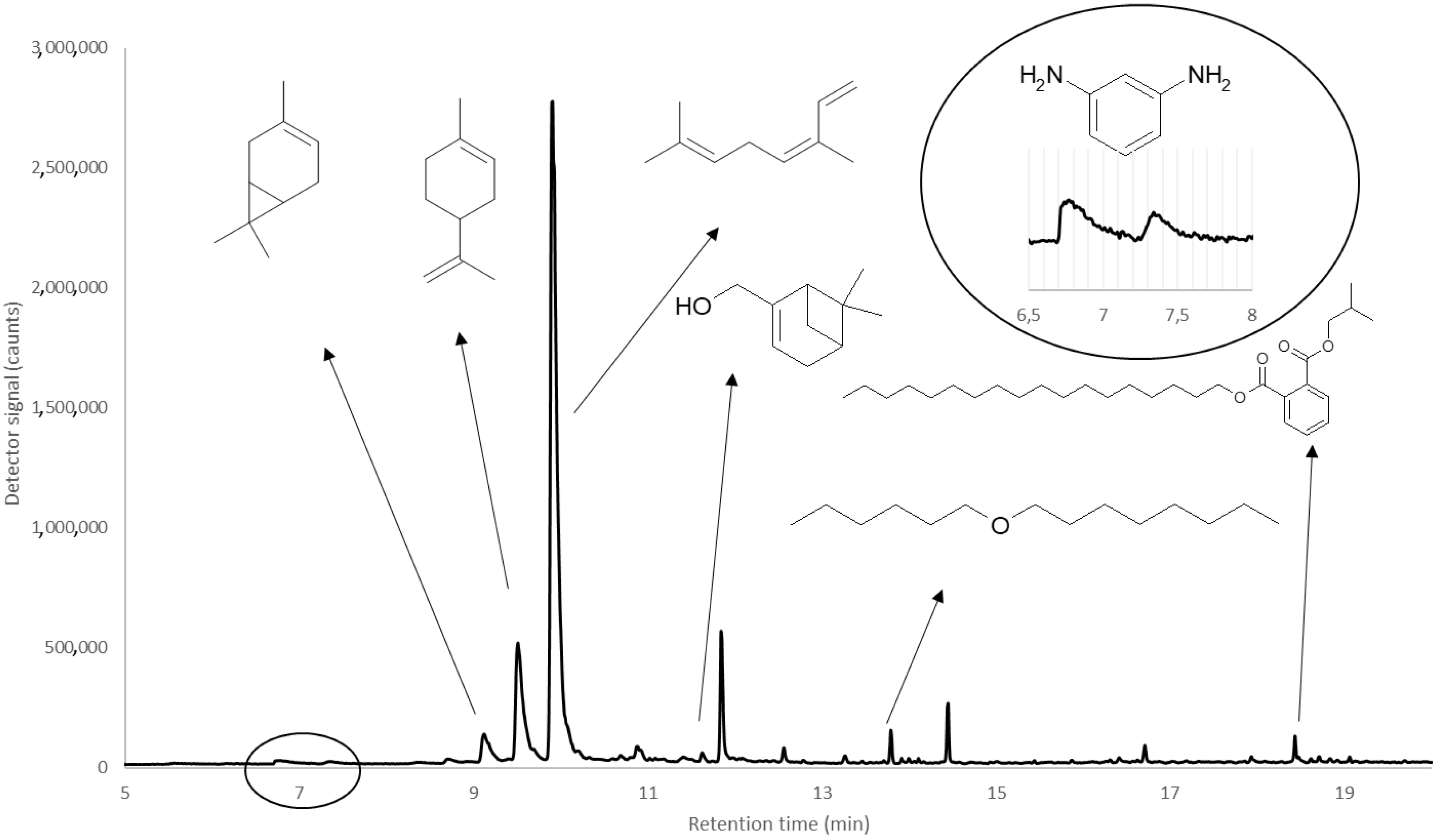
| Compound Name | m-Phenylene Diamine | 3-Carene | Limonene | Ocimene | 2-Pinen-10-ol | Hexyl Octyl ether | Phthalic Acid, Isobutyl Octadecyl Ester |
|---|---|---|---|---|---|---|---|
| Molecular weight (g/mol) | 108 | 136 | 136 | 136 | 152 | 214 | 474 |
| Calculated Kováts retention index * | 212 | 1016 | 1034 | 1052 | 1105 | 1325 | 1520 |
| Reference Kováts retention index | 200–220 | 1004–1017 | 1031–1056 | 1043–1097 | 1106 | 1302 | - |
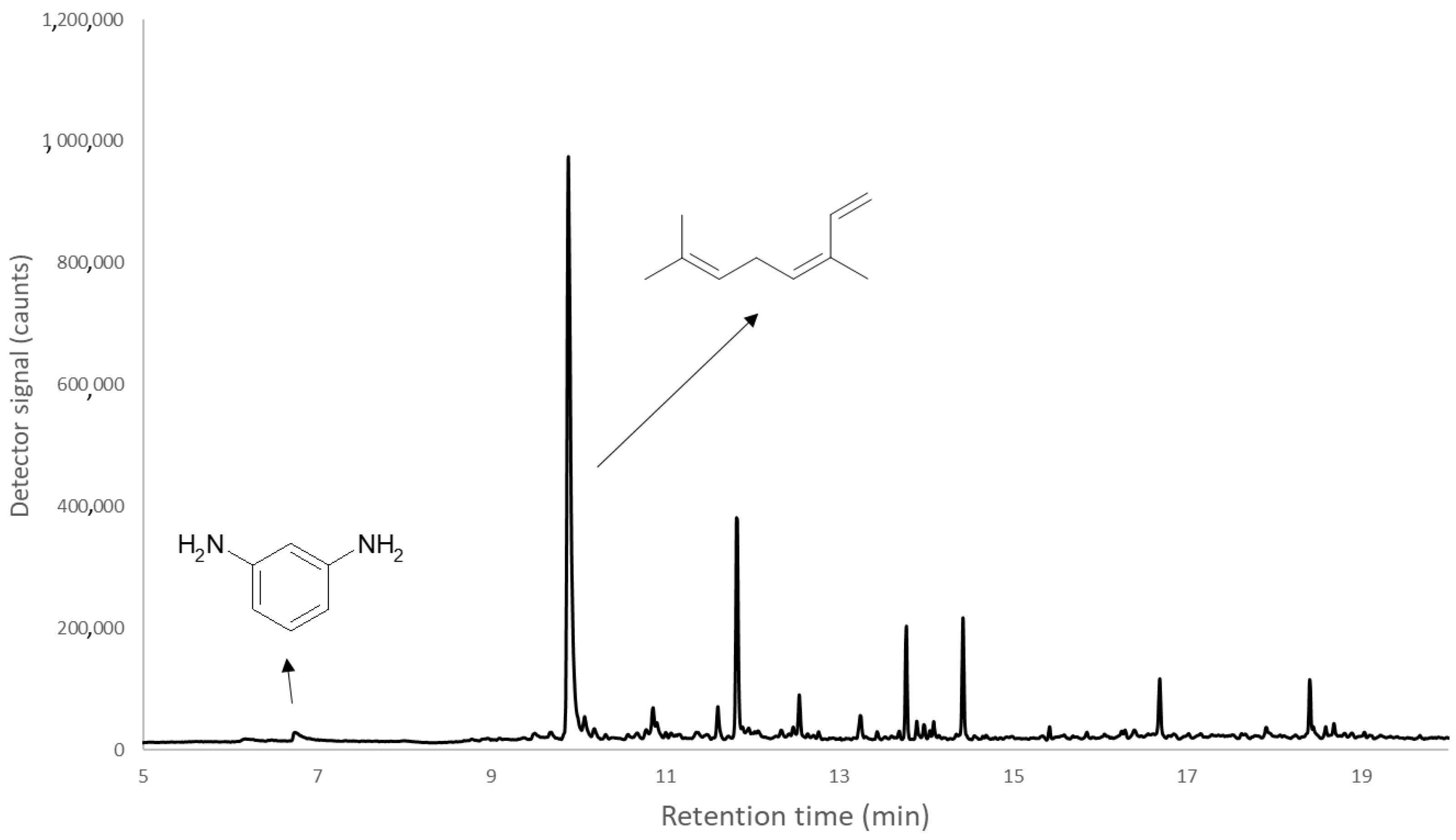
| Order Number | P. larvae Infection | m-Phenylene Diamine | 3-Carene | Limonene | Ocimene | 2-Pinen-10-ol | Hexyl Octyl Ether | Phthalic Acid, Isobutyl Octadecyl Ester |
|---|---|---|---|---|---|---|---|---|
| 1 | - | <LOD * | 6.543 | 19.468 | 59.351 | 0.824 | 2.075 | <LOD |
| 2 | - | <LOD | 8.800 | 45.525 | <LOD | <LOD | 5.252 | 3.375 |
| 3 | - | <LOD | 6.131 | 15.363 | 59.926 | 0.817 | 1.535 | <LOD |
| 4 | - | <LOD | 7.908 | 17.470 | 67.981 | 0.691 | 1.212 | <LOD |
| 5 | - | <LOD | 5.228 | 13.233 | 73.066 | 0.836 | 0.771 | <LOD |
| 6 | - | <LOD | <LOD | <LOD | 14.288 | <LOD | 12.728 | Trace |
| 7 | - | <LOD | 6.668 | 16.213 | 49.362 | 1.503 | 2.648 | 1.524 |
| 8 | - | <LOD | 5.810 | 18.133 | 32.959 | 1.150 | 2.834 | Trace |
| 9 | - | Trace ** | 2.617 | 9.500 | 53.577 | 0.290 | 0.773 | 0.480 |
| 10 | - | <LOD | 5.432 | 18.000 | 46.589 | <LOD | 4.280 | Trace |
| 11 | - | <LOD | 7.069 | 39.625 | 9.021 | <LOD | 4.661 | Trace |
| 12 | + | Trace | 9.702 | 53.232 | <LOD | <LOD | 3.933 | 2.079 |
| 13 | + | <LOD | 3.650 | 12.827 | 44.854 | 1.971 | 4.781 | Trace |
| 14 | + | <LOD | 1.188 | 19.502 | 50.843 | <LOD | 2.848 | <LOD |
| 15 | + | <LOD | 8.687 | 47.592 | <LOD | <LOD | 2.954 | 3.418 |
| 16 | + | <LOD | <LOD | <LOD | 19.330 | <LOD | 2.944 | 6.667 |
| 17 | + | <LOD | <LOD | <LOD | 81.000 | 0.753 | 2.115 | Trace |
| 18 | + | Trace | <LOD | <LOD | 42.099 | <LOD | 4.181 | 2.949 |
| 19 | + | Trace | <LOD | <LOD | 71.145 | 0.639 | <LOD | Trace |
| 20 | + | <LOD | <LOD | <LOD | 59.698 | Trace | 5.891 | <LOD |
| 21 | + | <LOD | <LOD | <LOD | 87.240 | 0.767 | <LOD | <LOD |
| 22 | + | <LOD | <LOD | <LOD | 77.311 | Trace | <LOD | <LOD |
| 23 | + | Trace | <LOD | <LOD | 46.623 | 0.672 | <LOD | <LOD |
| 24 | + | <LOD | <LOD | <LOD | 23.191 | <LOD | 12.923 | <LOD |
| 25 | + | <LOD | <LOD | <LOD | 53.884 | Trace | 2.699 | Trace |
| Compound | p-Value for χ2 Test * |
|---|---|
| m-phenylenediamine | 0.481 |
| 3-Carene | 0.007 ** |
| Limonene | 0.007 ** |
| Ocimene | 0.823 |
| 2-pinen-10-ol | 0.934 |
| hexyl octyl ether | 0.167 |
| phthalic acid, isobutyl octadecyl ester | 0.934 |
| Compound | U Mann–Whitney Statistics | p | Stimulant |
|---|---|---|---|
| 3-Carene | 2.72 | 0.007 * | n ** |
| Limonene | 2.20 | 0.028 * | n |
| Ocimene | −0.36 | 0.722 | - |
| 2-Pinen-10-ol | 1.10 | 0.270 | - |
| Hexyl octyl ether | 0.03 | 0.978 | - |
| Phthalic acid, isobutyl octadecyl ester | −0.03 | 0.977 | - |
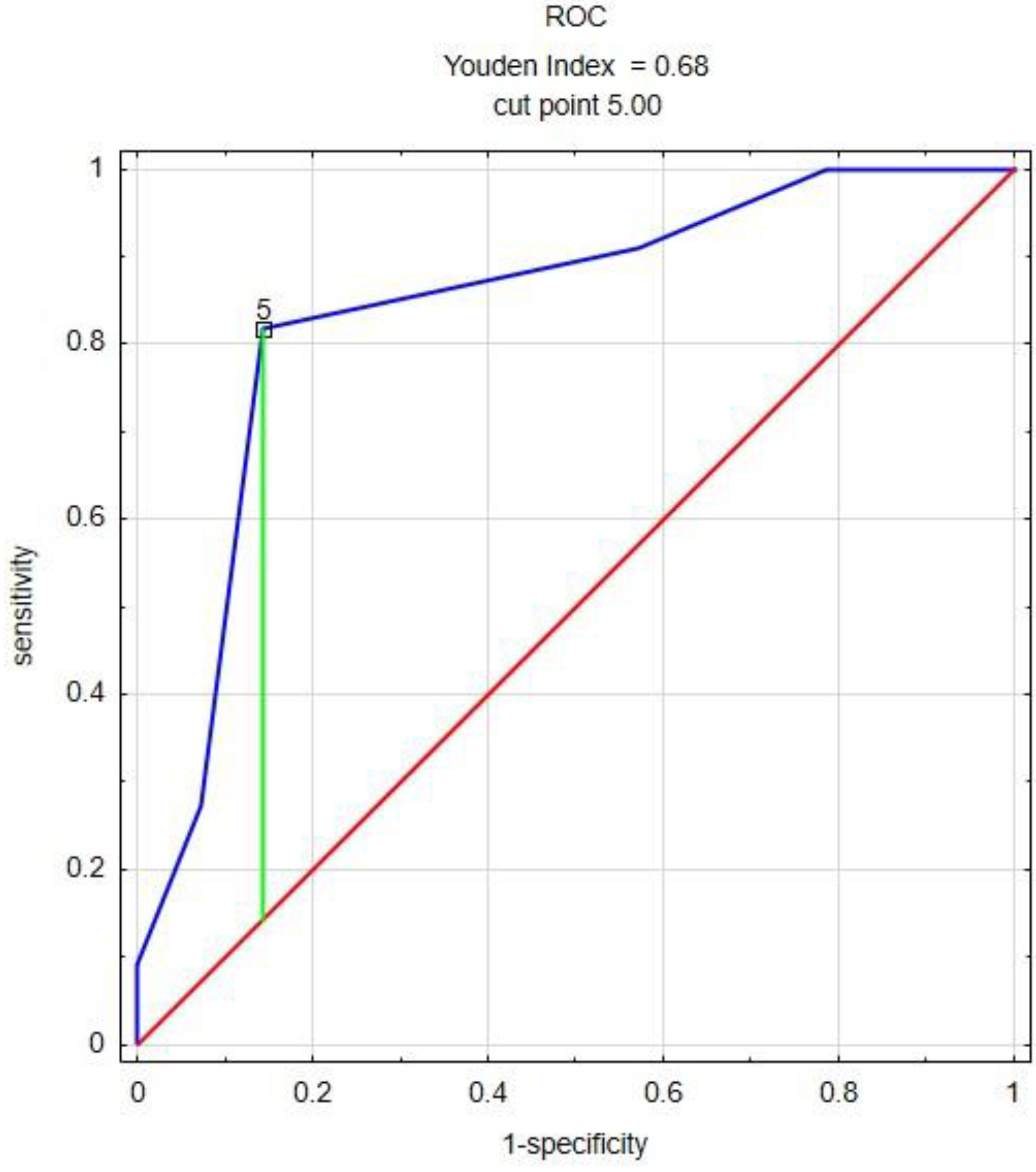
| Compound | Cut Point | AUC * | SE ** | AUC Lower 95% | AUC Upper 95% | p |
|---|---|---|---|---|---|---|
| 3-Carene | 2.617 | 0.812 | 0.098 | 0.619 | 1.000 | 0.0015 |
| Limonene | 9.5 | 0.753 | 0.108 | 0.541 | 0.965 | 0.0191 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piechowicz, B.; Kwiatek, A.; Sadło, S.; Zaręba, L.; Koziorowska, A.; Kloc, D.; Balawejder, M. Use of Gas Chromatography and SPME Extraction for the Differentiation between Healthy and Paenibacillus larvae Infected Colonies of Bee Brood—Preliminary Research. Agriculture 2023, 13, 487. https://doi.org/10.3390/agriculture13020487
Piechowicz B, Kwiatek A, Sadło S, Zaręba L, Koziorowska A, Kloc D, Balawejder M. Use of Gas Chromatography and SPME Extraction for the Differentiation between Healthy and Paenibacillus larvae Infected Colonies of Bee Brood—Preliminary Research. Agriculture. 2023; 13(2):487. https://doi.org/10.3390/agriculture13020487
Chicago/Turabian StylePiechowicz, Bartosz, Aleksandra Kwiatek, Stanisław Sadło, Lech Zaręba, Anna Koziorowska, Daniela Kloc, and Maciej Balawejder. 2023. "Use of Gas Chromatography and SPME Extraction for the Differentiation between Healthy and Paenibacillus larvae Infected Colonies of Bee Brood—Preliminary Research" Agriculture 13, no. 2: 487. https://doi.org/10.3390/agriculture13020487