
A Colorimetric LAMP Detection of Xylella fastidiosa in Crude Alkaline Sap of Olive Trees in Apulia as a Field-Based Tool for Disease Containment
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Preparation of Cloned Plasmid Standard
2.3. Plant Materials
2.4. Crude Plant Extracts Preparation
2.5. cLAMP Assay and Device
2.6. Analytical Sensitivity of cLAMP Assay
2.7. Evaluation of cLAMP Performance on Field-Grown Olive Samples
2.8. DNA Extraction and Real Time PCR (qPCR) Assay
2.9. Droplet Digital PCR Assay
2.10. Statistical Analysis
3. Results
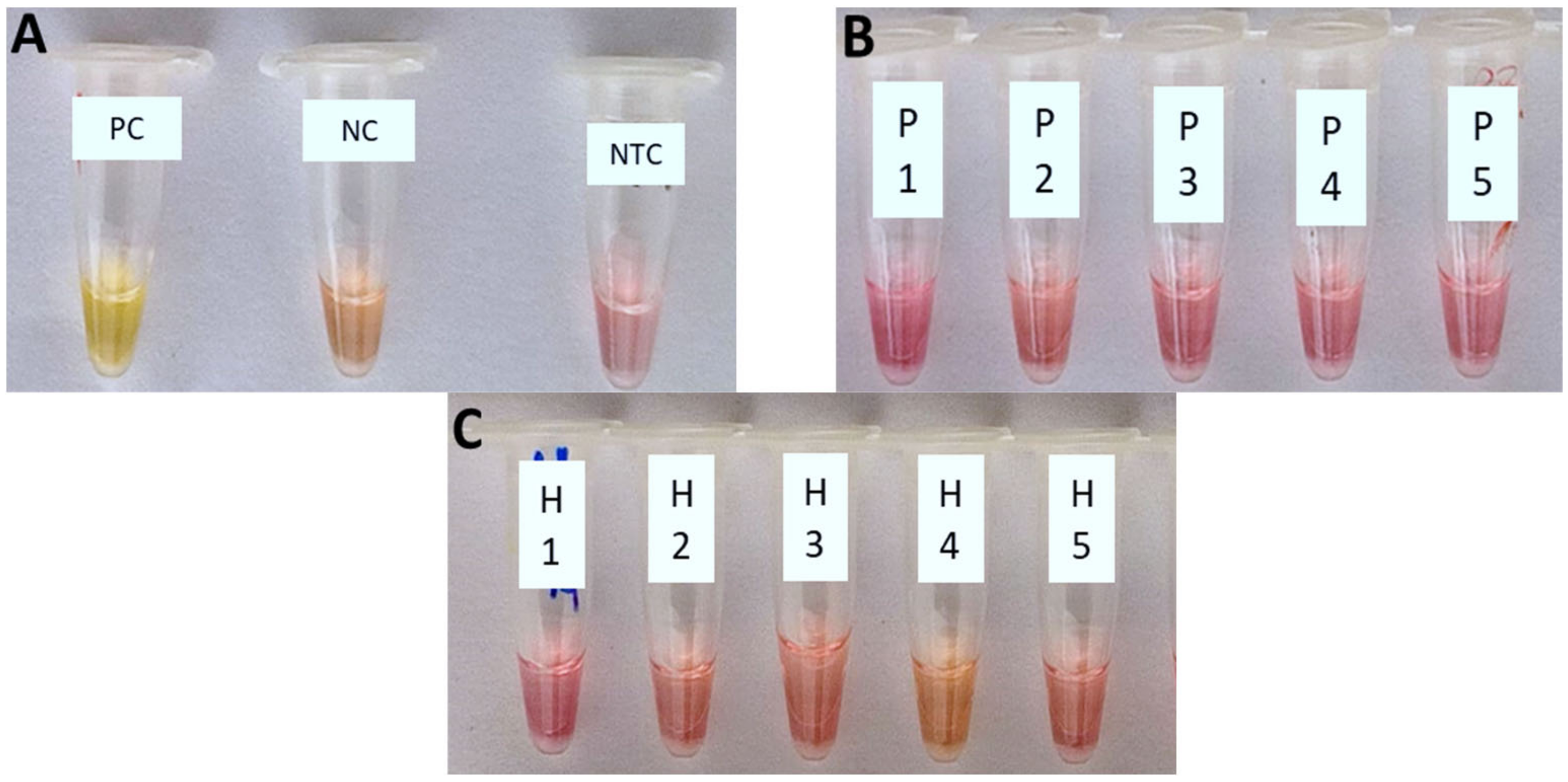
3.1. Specificity of Xfp Colorimetric LAMP Assay
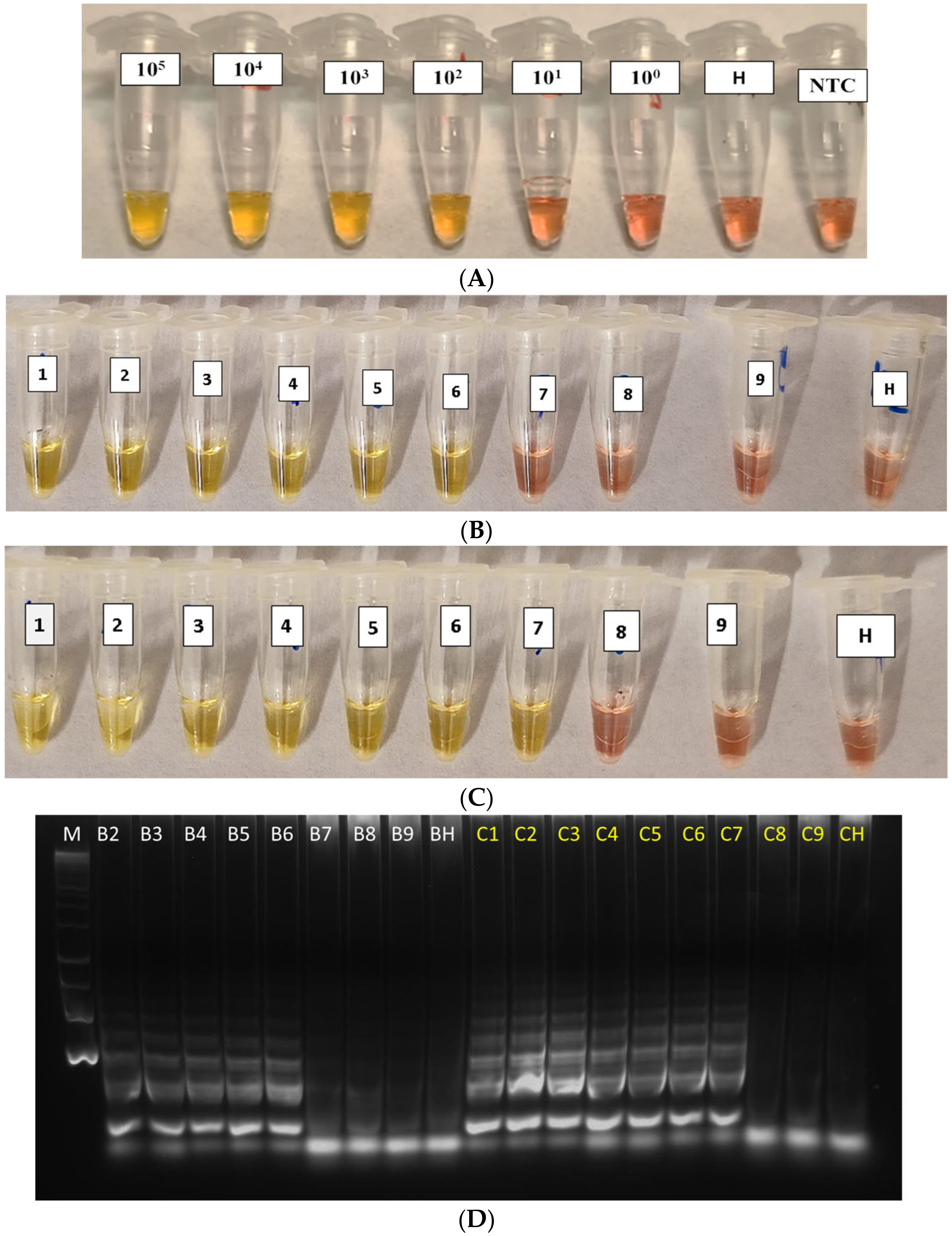
3.2. Sensitivity of Xfp Colorimetric LAMP Assay
3.3. Validation of cLAMP Assay on Field-Grown Olive Samples by Comparison with qPCR
3.4. Evaluation of ddPCR Performance and Comparison with cLAMP
3.5. Statistical Data Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wells, J.M.; Raju, B.C.; Hung, H.-Y.; Weisburg, W.G.; Mandelco-Paul, L.; Brenner, D.J. Xylella Fastidiosa Gen. Nov., Sp. Nov: Gram-Negative, Xylem-Limited, Fastidious Plant Bacteria Related to Xanthomonas Spp. Int. J. Syst. Bacteriol. 1987, 37, 136–143. [Google Scholar] [CrossRef]
- Landa, B.B.; Saponari, M.; Feitosa, O.R., Jr.; Giampetruzzi, A.; Vieira, F.J.D.; Mor, E.; Robatzek, S. Xylella Fastidiosa’s Relationships: The Bacterium, the Host Plants, and the Plant Microbiome. New Phytol. 2022, 234, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.L. Xylella Fastidiosa: Xylem-Limited Bacterial Pathogen of Plants. Annu. Rev. Phytopathol. 1989, 27, 271–290. [Google Scholar] [CrossRef]
- Purcell, A.H.; Hopkins, D.L. Fastidious Xylem-Limited Bacterial Plant Pathogens. Annu. Rev. Phytopathol. 1996, 34, 131–151. [Google Scholar] [CrossRef]
- Hill, B.L.; Purcell, A.M. Populations of Xylella Fastidiosa in Plants Required for Transmission by an Efficient Vector. Phytopathology 1997, 87, 1197–1201. [Google Scholar] [CrossRef]
- Delbianco, A.; Gibin, D.; Pasinato, L.; Morelli, M. Update of the Xylella Spp. Host Plant Database—Systematic Literature Search up to 30 June 2021. EFSA J. 2022, 20, e07039. [Google Scholar] [CrossRef]
- PM 7/24 (4) Xylella Fastidiosa. EPPO Bull. 2019, 49, 175–227. [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef]
- Tomlinson, J.A.; Ostoja-Starzewska, S.; Webb, K.; Cole, J.; Barnes, A.; Dickinson, M.; Boonham, N. A Loop-Mediated Isothermal Amplification-Based Method for Confirmation of Guignardia Citricarpa in Citrus Black Spot Lesions. Eur. J. Plant Pathol. 2013, 136, 217–224. [Google Scholar] [CrossRef]
- Becherer, L.; Borst, N.; Bakheit, M.; Frischmann, S.; Zengerle, R.; Von Stetten, F. Loop-Mediated Isothermal Amplification (LAMP)-Review and Classification of Methods for Sequence-Specific Detection. Anal. Methods 2020, 12, 717–746. [Google Scholar] [CrossRef]
- Baldi, P.; La Porta, N. Molecular Approaches for Low-Cost Point-of-Care Pathogen Detection in Agriculture and Forestry. Front. Plant Sci. 2020, 11, 570862. [Google Scholar] [CrossRef]
- Gadkar, V.J.; Goldfarb, D.M.; Gantt, S.; Tilley, P.A.G. Real-Time Detection and Monitoring of Loop Mediated Amplification (LAMP) Reaction Using Self-Quenching and De-Quenching Fluorogenic Probes. Sci. Rep. 2018, 8, 5548. [Google Scholar] [CrossRef]
- Wang, D.; Yu, J.; Wang, Y.; Zhang, M.; Li, P.; Liu, M.; Liu, Y. Development of a Real-Time Loop-Mediated Isothermal Amplification (LAMP) Assay and Visual LAMP Assay for Detection of African Swine Fever Virus (ASFV). J. Virol. Methods 2020, 276, 113775. [Google Scholar] [CrossRef]
- Papadakis, G.; Pantazis, A.K.; Fikas, N.; Chatziioannidou, S.; Tsiakalou, V.; Michaelidou, K.; Pogka, V.; Megariti, M.; Vardaki, M.; Giarentis, K.; et al. Portable Real-Time Colorimetric LAMP-Device for Rapid Quantitative Detection of Nucleic Acids in Crude Samples. Sci. Rep. 2022, 12, 3775. [Google Scholar] [CrossRef]
- Cibecchini, G.; Cecere, P.; Tumino, G.; Morcia, C.; Ghizzoni, R.; Carnevali, P.; Terzi, V.; Pompa, P.P. A Fast, Naked-Eye Assay for Varietal Traceability in the Durum Wheat Production Chain. Foods 2020, 9, 1691. [Google Scholar] [CrossRef]
- Drais, M.I.; Maheshwari, Y.; Selvaraj, V.; Varvaro, L.; Yokomi, R.; Djelouah, K. Development and Validation of a Loop-Mediated Isothermal Amplification Technique (LAMP) for the Detection of Spiroplasma Citri, the Causal Agent of Citrus Stubborn Disease. Eur. J. Plant Pathol. 2019, 155, 125–134. [Google Scholar] [CrossRef]
- Stehlíková, D.; Beran, P.; Cohen, S.P.; Čurn, V. Development of Real-Time and Colorimetric Loop Mediated Isothermal Amplification Assay for Detection of Xanthomonas Gardneri. Microorganisms 2020, 8, 1301. [Google Scholar] [CrossRef]
- Bertacca, S.; Caruso, A.G.; Trippa, D.; Marchese, A.; Giovino, A.; Matic, S.; Noris, E.; Ambrosio, M.I.F.S.; Alfaro, A.; Panno, S.; et al. Development of a Real-Time Loop-Mediated Isothermal Amplification Assay for the Rapid Detection of Olea Europaea Geminivirus. Plants 2022, 11, 660. [Google Scholar] [CrossRef]
- Sadallah, A.; Minutillo, S.A.; Valentini, F.; Raimondo, M.L.; Lops, F.; Carlucci, A.; Ippolito, A.; D’Onghia, A.M. A Real Time Loop-Mediated Isothermal Amplification (RealAmp) Assay for Rapid Detection of Pleurostoma Richardsiae in Declining Olive Plants. Phytopathol. Mediterr. 2022, 61, 259–267. [Google Scholar] [CrossRef]
- Hansen, Z.R.; Knaus, B.J.; Tabima, J.F.; Press, C.M.; Judelson, H.S.; Grünwald, N.J.; Smart, C.D. Loop-Mediated Isothermal Amplification for Detection of the Tomato and Potato Late Blight Pathogen, Phytophthora Infestans. J. Appl. Microbiol. 2016, 120, 1010–1020. [Google Scholar] [CrossRef]
- Kogovšek, P.; Hodgetts, J.; Hall, J.; Prezelj, N.; Nikolić, P.; Mehle, N.; Lenarčič, R.; Rotter, A.; Dickinson, M.; Boonham, N.; et al. LAMP Assay and Rapid Sample Preparation Method for On-Site Detection of Flavescence Dorée Phytoplasma in Grapevine. Plant Pathol. 2015, 64, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Kogovšek, P.; Mehle, N.; Pugelj, A.; Jakomin, T.; Schroers, H.J.; Ravnikar, M.; Dermastia, M. Rapid Loop-Mediated Isothermal Amplification Assays for Grapevine Yellows Phytoplasmas on Crude Leaf-Vein Homogenate Has the Same Performance as QPCR. Eur. J. Plant Pathol. 2017, 148, 75–84. [Google Scholar] [CrossRef]
- Matić, S.; Candian, V.; D’Errico, C.; Pierro, R.; Panno, S.; Davino, S.; Noris, E.; Tedeschi, R. In-Field LAMP Detection of Flavescence Dorée Phytoplasma in Crude Extracts of the Scaphoideus Titanus Vector. Agronomy 2022, 12, 1645. [Google Scholar] [CrossRef]
- Harper, S.J.; Ward, L.I.; Clover, G.R.G. Development of LAMP and Real-Time PCR Methods for the Rapid Detection of Xylella Fastidiosa for Quarantine and Field Applications. Phytopathology 2010, 100, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Aglietti, C.; Luchi, N.; Pepori, A.L.; Bartolini, P.; Pecori, F.; Raio, A.; Capretti, P.; Santini, A. Real-Time Loop-Mediated Isothermal Amplification: An Early-Warning Tool for Quarantine Plant Pathogen Detection. AMB Express 2019, 9, 50. [Google Scholar] [CrossRef]
- Yaseen, T.; Drago, S.; Valentini, F.; Elbeaino, T.; Stampone, G.; D’Onghia, A.M. On-Site Detection of Xylella Fastidiosa in Host Plants and in “Spy Insects” Using the Real-Time Loop-Mediated Isothermal Amplification Method. Phytopathol. Mediterr. 2015, 54, 488–496. [Google Scholar]
- Elbeaino, T.; Incerti, O.; Dakroub, H.; Valentini, F.; Huang, Q. Development of an FTP-LAMP Assay Based on TaqMan Real-Time PCR and LAMP for the Specific Detection of Xylella Fastidiosa De Donno and Mulberry Strains in Both Plants and Insect Vectors. J. Microbiol. Methods 2020, 175, 105992. [Google Scholar] [CrossRef]
- Kim, S.; Park, Y.; Kim, G. Development of Diagnostic Technology of Xylella Fastidiosa Using Loop-Mediated Isothermal Amplification and PCR Methods. Res. Plant Dis. 2021, 27, 38–44. [Google Scholar] [CrossRef]
- Waliullah, S.; Di Genova, D.; Oliver, J.E.; Ali, M.E. Development of a CAPS Marker and a LAMP Assay for Rapid Detection of Xylella Fastidiosa Subsp. Multiplex and Differentiation from X. Fastidiosa Subsp. Fastidiosa on BlueBerry. Int. J. Mol. Sci. 2022, 23, 1937. [Google Scholar] [CrossRef]
- Chiriacò, M.S.; Luvisi, A.; Primiceri, E.; Sabella, E.; De Bellis, L.; Maruccio, G. Development of a Lab-on-a-Chip Method for Rapid Assay of Xylella Fastidiosa Subsp. Pauca Strain CoDiRO. Sci. Rep. 2018, 8, 7376. [Google Scholar] [CrossRef]
- Sarcina, L.; Macchia, E.; Loconsole, G.; D’Attoma, G.; Bollella, P.; Catacchio, M.; Leonetti, F.; Di Franco, C.; Elicio, V.; Scamarcio, G.; et al. Fast and Reliable Electronic Assay of a Xylella Fastidiosa Single Bacterium in Infected Plants Sap. Adv. Sci. 2022, 9, 2203900. [Google Scholar] [CrossRef]
- Dupas, E.; Legendre, B.; Olivier, V.; Poliakoff, F.; Manceau, C.; Cunty, A. Comparison of Real-Time PCR and Droplet Digital PCR for the Detection of Xylella Fastidiosa in Plants. J. Microbiol. Methods 2019, 162, 86–95. [Google Scholar] [CrossRef]
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification Of Dna Sequences Related To Xylella Fastidiosa In Oleander, Almond And Olive Trees Exhibiting Leaf Scorch Symptoms In Apulia (Southern Italy). J. Plant Pathol. 2013, 95, 668. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Saponari, M.; Almeida, R.P.P.; Essakhi, S.; Boscia, D.; Loconsole, G.; Saldarelli, P. Complete Genome Sequence of the Olive-Infecting Strain Xylella Fastidiosa Subsp. Pauca De Donno. Genome Announc. 2017, 5, e00569-17. [Google Scholar] [CrossRef]
- D’attoma, G.; Morelli, M.; Saldarelli, P.; Saponari, M.; Giampetruzzi, A.; Boscia, D.; Savino, V.N.; Fuente, L.D.L.; Cobine, P.A. Ionomic Differences between Susceptible and Resistant Olive Cultivars Infected by Xylella Fastidiosa in the Outbreak Area of Salento, Italy. Pathogens 2019, 8, 272. [Google Scholar] [CrossRef]
- Baccari, C.; Antonova, E.; Lindow, S. Biological Control of Pierce’s Disease of Grape by an Endophytic Bacterium. Phytopathology 2019, 109, 248–256. [Google Scholar] [CrossRef]
- Sun, X.; Ma, W.; Xu, Y.; Jin, X.; Ni, H. Complete Genome Sequence of Rahnella Aquatilis MEM40, a Plant Growth-Promoting Rhizobacterium Isolated from Rice Rhizosphere Soil, with Antagonism against Magnaporthe Oryzae and Fusarium Graminearum. Microbiol. Resour. Announc. 2020, 9, e00651-20. [Google Scholar] [CrossRef]
- Loconsole, G.; Potere, O.; Boscia, D.; Altamura, G.; Djelouah, K.; Elbeaino, T.; Frasheri, D.; Lorusso, D.; Palmisano, F.; Pollastro, P.; et al. Detection of Xylella Fastidiosa in Olive Trees By Molecular And Serological Methods. J. Plant Pathol. 2014, 96, 7–14. [Google Scholar] [CrossRef]
- PM 7/76 (5) Use of EPPO Diagnostic Standards. EPPO Bull. 2018, 48, 373–377. [CrossRef]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef]
- Cleophas, T.J.; Zwinderman, A.H. Modern Bayesian Statistics in Clinical Research; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-92746-6. [Google Scholar]
- Altman, D.G. Practical Statistics for Medical Research; Chapman & Hall/CRC Press: New York, NY, USA, 1999. [Google Scholar]
- Saponari, M.; Giampetruzzi, A.; Loconsole, G.; Boscia, D.; Saldarelli, P. Xylella Fastidiosa in Olive in Apulia: Where We Stand. Phytopathology 2019, 109, 175–186. [Google Scholar] [CrossRef] [PubMed]
- González-González, E.; Lara-Mayorga, I.M.; Rodríguez-Sánchez, I.P.; Zhang, Y.S.; Martínez-Chapa, S.O.; Santiago, G.T.; Alvarez, M.M. Colorimetric Loop-Mediated Isothermal Amplification (LAMP) for Cost-Effective and Quantitative Detection of SARS-CoV-2: The Change in Color in LAMP-Based Assays Quantitatively Correlates with Viral Copy Number. Anal. Methods Adv. Methods Appl. 2021, 13, 169–178. [Google Scholar] [CrossRef] [PubMed]
- García-Bernalt Diego, J.; Fernández-Soto, P.; Márquez-Sánchez, S.; Santos Santos, D.; Febrer-Sendra, B.; Crego-Vicente, B.; Muñoz-Bellido, J.L.; Belhassen-García, M.; Corchado Rodríguez, J.M.; Muro, A. SMART-LAMP: A Smartphone-Operated Handheld Device for Real-Time Colorimetric Point-of-Care Diagnosis of Infectious Diseases via Loop-Mediated Isothermal Amplification. Biosensors 2022, 12, 424. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| qPCR Cq Value | cLAMP Positives /Nr Analysed Plants (Per Cq Value) | Diagnostic Sensitivity % | Diagnostic Specificity % |
|---|---|---|---|
| 18–21 | 22/28 | 78.57 | / |
| 22 | 24/29 | 82.76 | / |
| 23 | 21/26 | 80.77 | / |
| 24 | 16/29 | 55.17 | / |
| 25 | 19/39 | 48.72 | / |
| 26 | 14/29 | 48.28 | / |
| 27 | 13/28 | 46.43 | / |
| 28 | 3/9 | 33.33 | / |
| 29–32 | 7/27 | 25.93 | / |
| 33–37 | 3/11 | / | 72.73 |
| N/A | 6/27 | / | 77.78 |
| Total | 148/282 |
| qPCR Cq Value | ddPCR Positive /Nr. Analysed Plants (Per Cq Value) | Diagnostic Sensitivity % | Diagnostic Specificity % |
|---|---|---|---|
| 18–21 | 9/10 | 90.00 | / |
| 22 | 7/9 | 77.78 | / |
| 23 | 12/12 | 100.00 | / |
| 24 | 8/10 | 80.00 | / |
| 25 | 12/14 | 85.71 | / |
| 26 | 13/15 | 86.67 | / |
| 27 | 10/13 | 76.92 | / |
| 28–32 | 9/22 | 40.91 | / |
| NA | 1/10 | / | 90.00 |
| Total | 81/115 | / |
| qPCR Cq Value | No. ddPCR-Tested Samples | Mean of Positive Droplet ± SD | cLAMP Positive Samples | % PA in cLAMP Assay |
|---|---|---|---|---|
| 19–21 | 10 | 247.0 ± 312.87 | 7 | 70.00 |
| 22 | 9 | 140.2 ± 155.55 | 6 | 66.67 |
| 23 | 12 | 56.4 ± 54.92 | 10 | 83.33 |
| 24 | 10 | 50.4 ± 51.63 | 6 | 60.00 |
| 25 | 14 | 29.9 ± 44.58 | 5 | 35.71 |
| 26 | 15 | 13.2 ± 12.51 | 5 | 33.33 |
| 27 | 13 | 9.4 ± 7.36 | 7 | 53.85 |
| 28–32 | 22 | 5.7 ± 7.33 | 7 | 31.82 |
| NA | 10 | 0.9 ± 1.38 | 2 | 80.00 |
| Total | 115 | 55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amoia, S.S.; Loconsole, G.; Ligorio, A.; Pantazis, A.K.; Papadakis, G.; Gizeli, E.; Minafra, A. A Colorimetric LAMP Detection of Xylella fastidiosa in Crude Alkaline Sap of Olive Trees in Apulia as a Field-Based Tool for Disease Containment. Agriculture 2023, 13, 448. https://doi.org/10.3390/agriculture13020448
Amoia SS, Loconsole G, Ligorio A, Pantazis AK, Papadakis G, Gizeli E, Minafra A. A Colorimetric LAMP Detection of Xylella fastidiosa in Crude Alkaline Sap of Olive Trees in Apulia as a Field-Based Tool for Disease Containment. Agriculture. 2023; 13(2):448. https://doi.org/10.3390/agriculture13020448
Chicago/Turabian StyleAmoia, Serafina Serena, Giuliana Loconsole, Angela Ligorio, Alexandros K. Pantazis, George Papadakis, Electra Gizeli, and Angelantonio Minafra. 2023. "A Colorimetric LAMP Detection of Xylella fastidiosa in Crude Alkaline Sap of Olive Trees in Apulia as a Field-Based Tool for Disease Containment" Agriculture 13, no. 2: 448. https://doi.org/10.3390/agriculture13020448