
Foliar Application of Salicylic Acid Mitigates Saline Stress on Physiology, Production, and Post-Harvest Quality of Hydroponic Japanese Cucumber
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
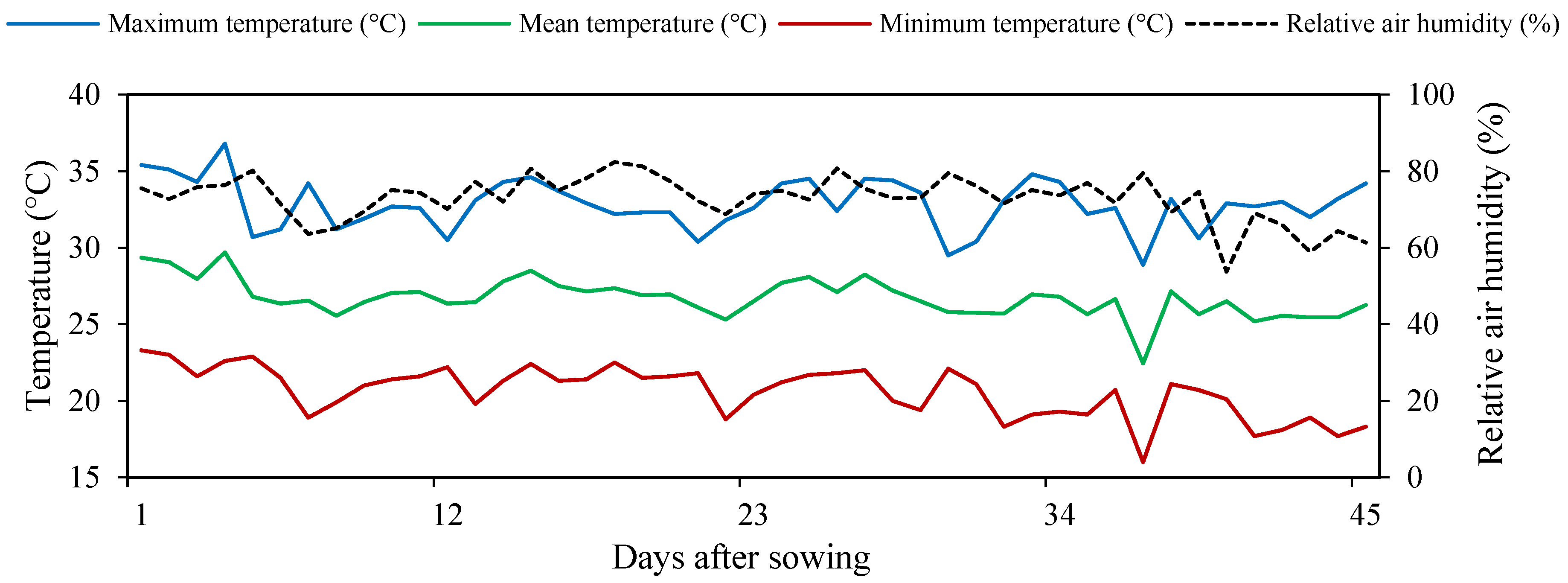
2.1. Experiment Site
2.2. Cultivar Studied
2.3. Experimental Design and Treatments
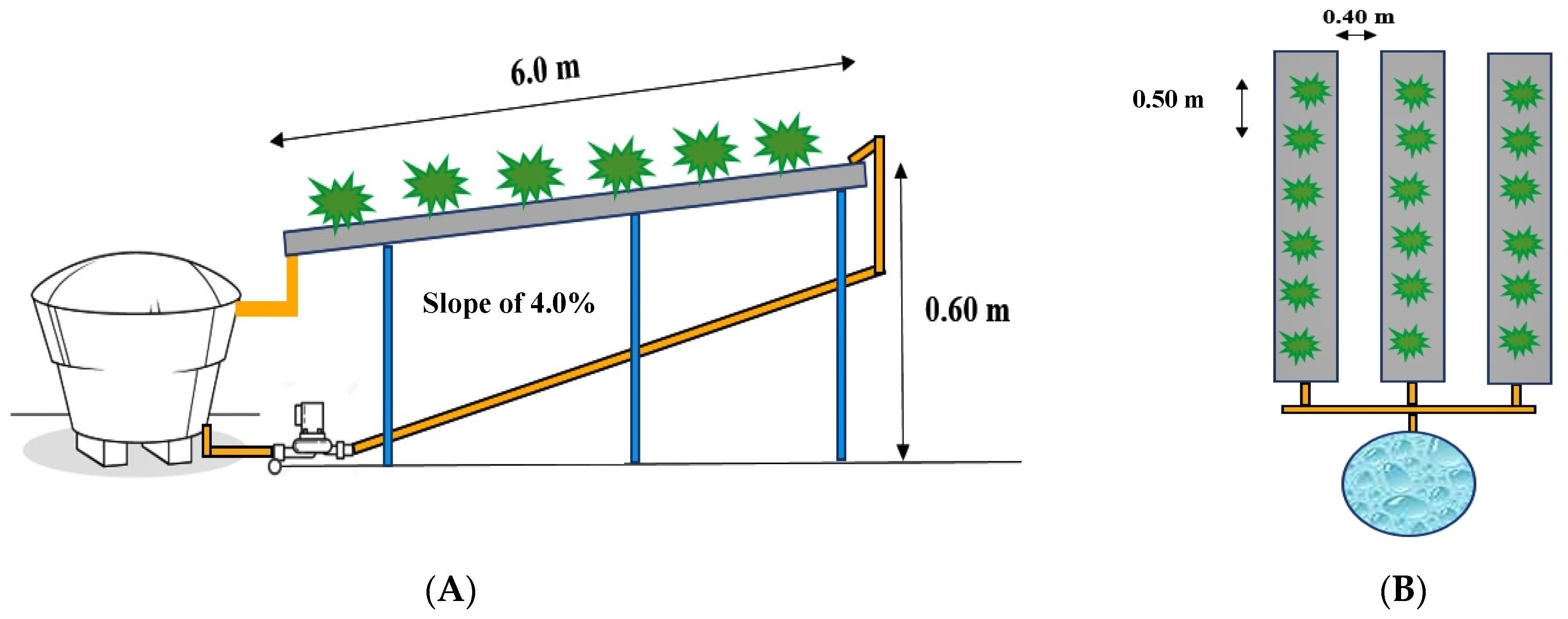
2.4. Setting Up and Management of Experiment
2.5. Traits Analyzed
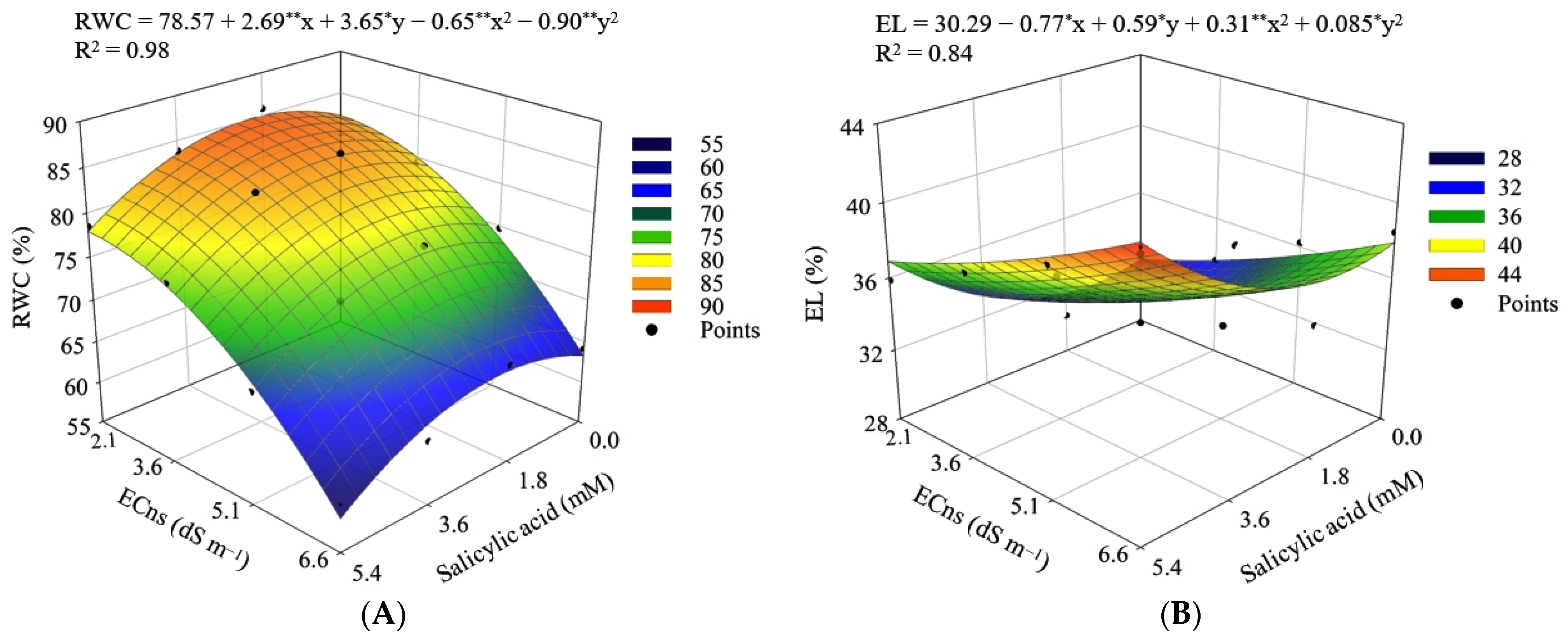
2.5.1. Relative Water Content
- RWC—relative water content (%);
- FM—fresh mass of leaves (g);
- TM—turgid mass (g);
- DM—dry mass (g).
2.5.2. Percentage of Electrolyte Leakage
- % EL—percentage of electrolyte leakage (%);
- Xi—initial electrical conductivity;
- Xf—final electrical conductivity.
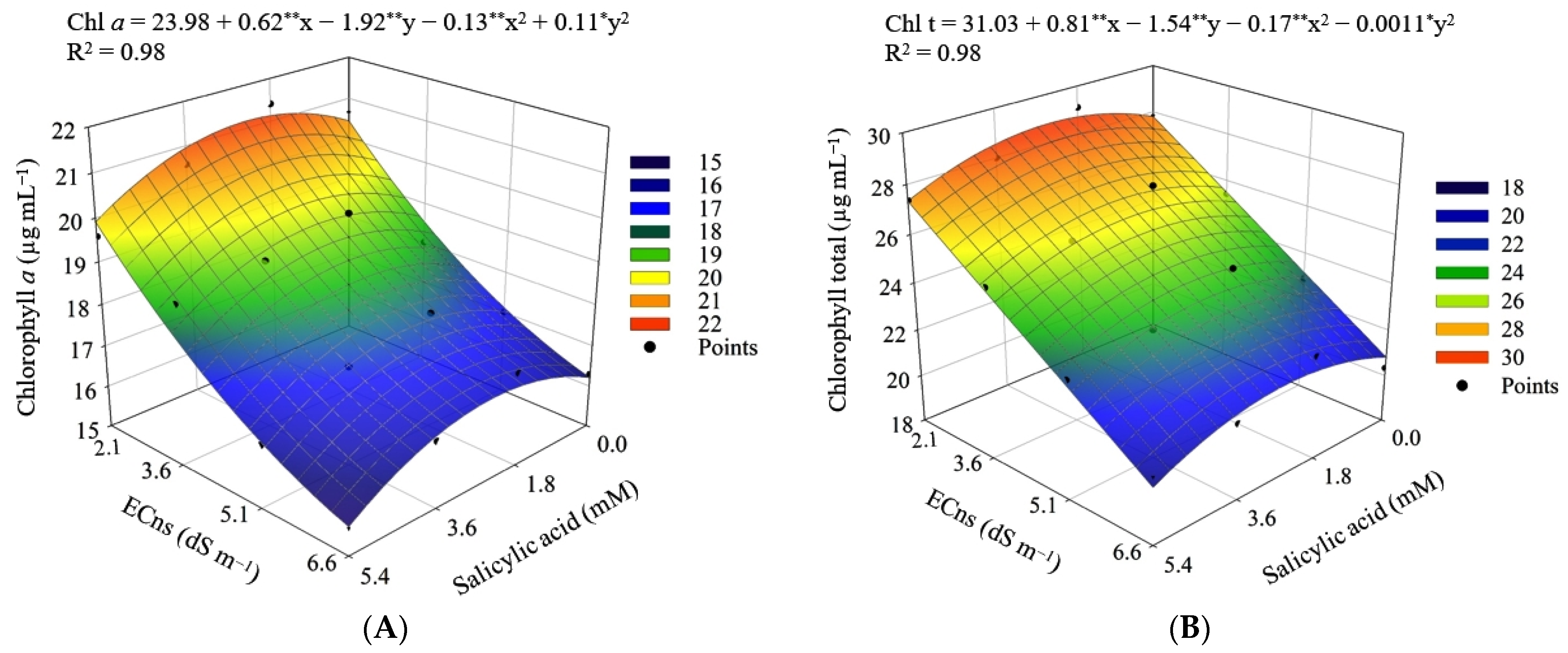
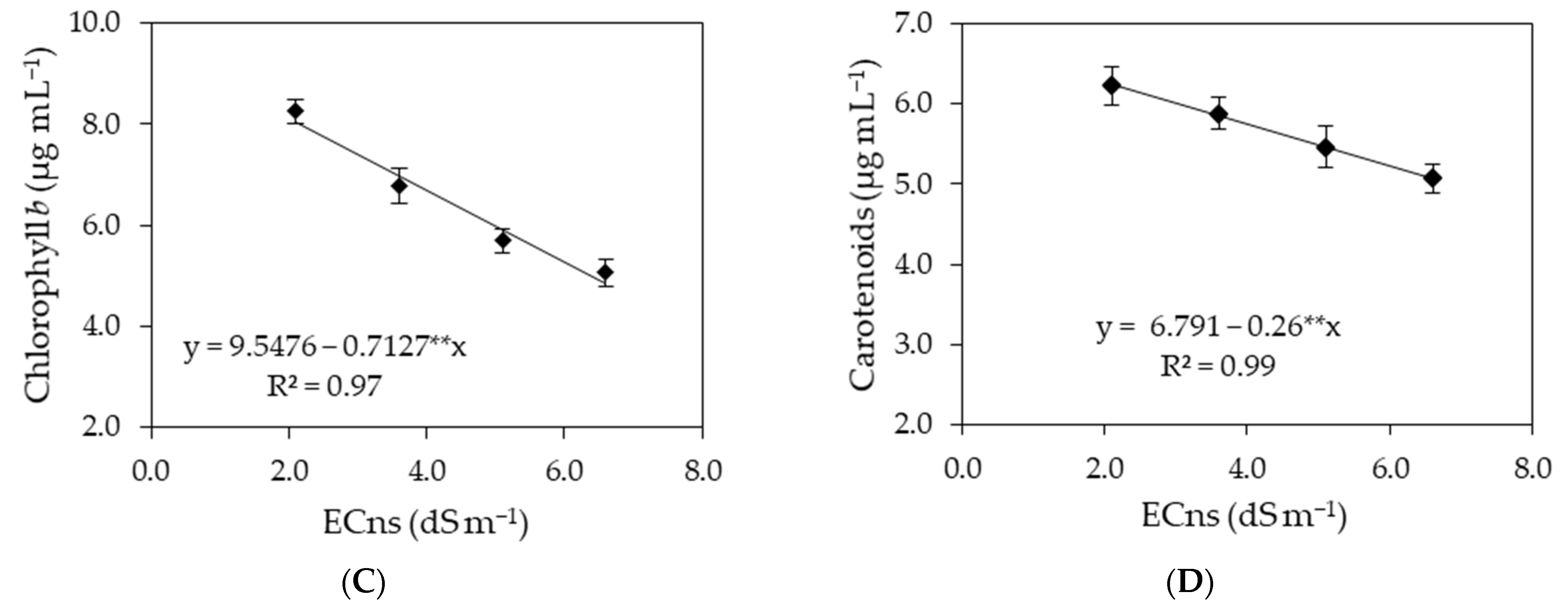
2.5.3. Photosynthetic Pigments
- Chl a—chlorophyll a;
- Chl b—chlorophyll b;
- Chl t—chlorophyll total;
- Car—carotenoids.
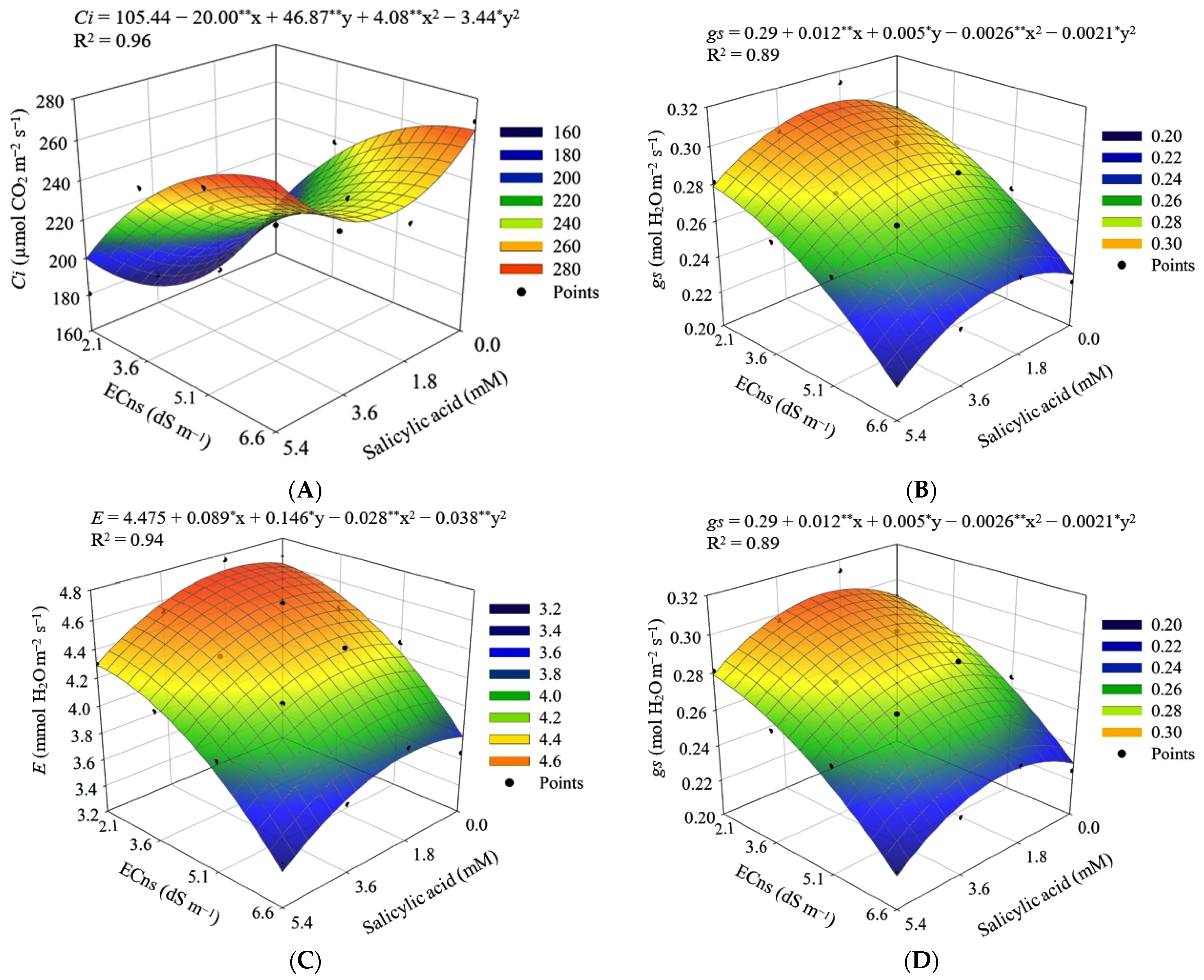
2.5.4. Leaf Gas Exchange Parameters
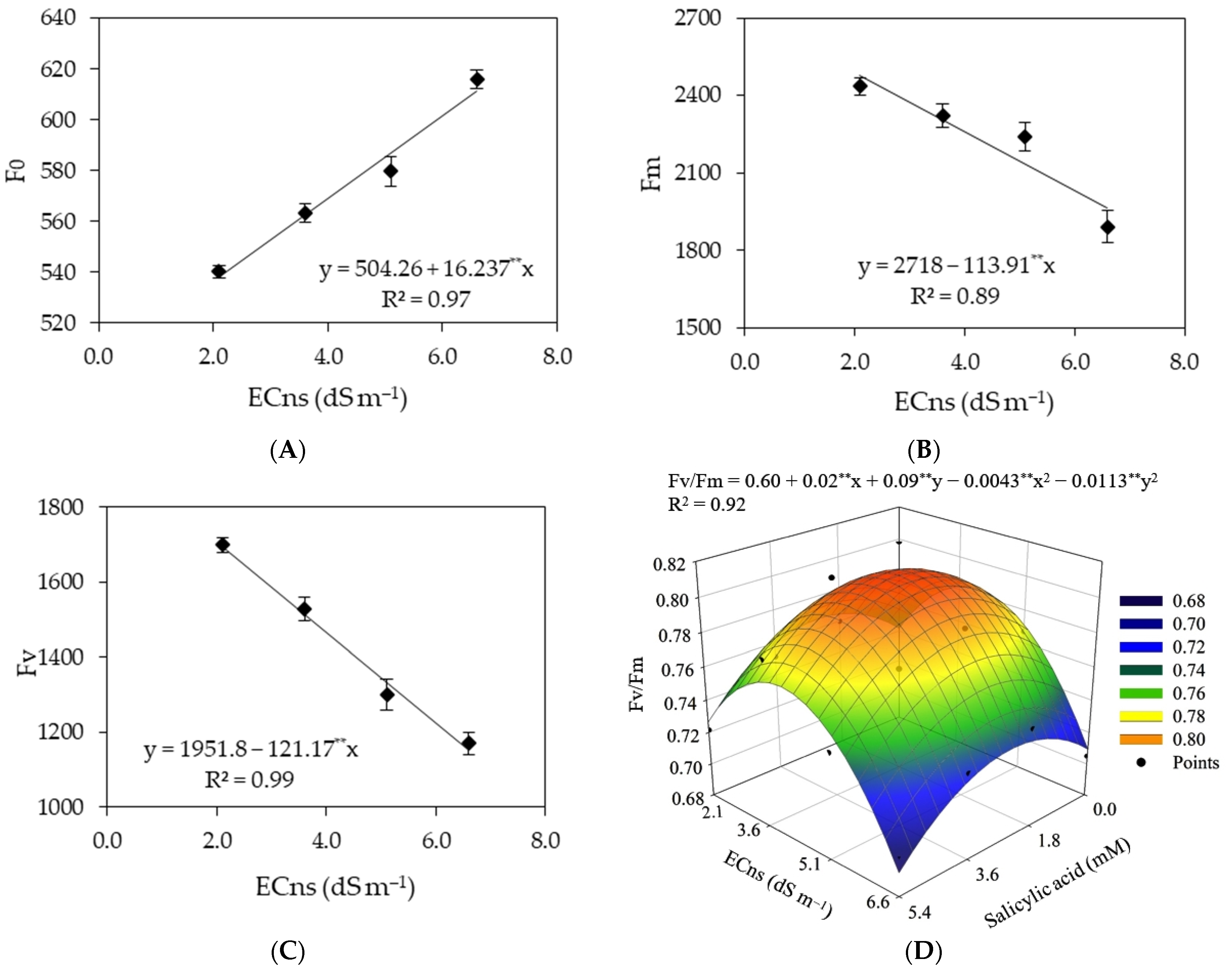
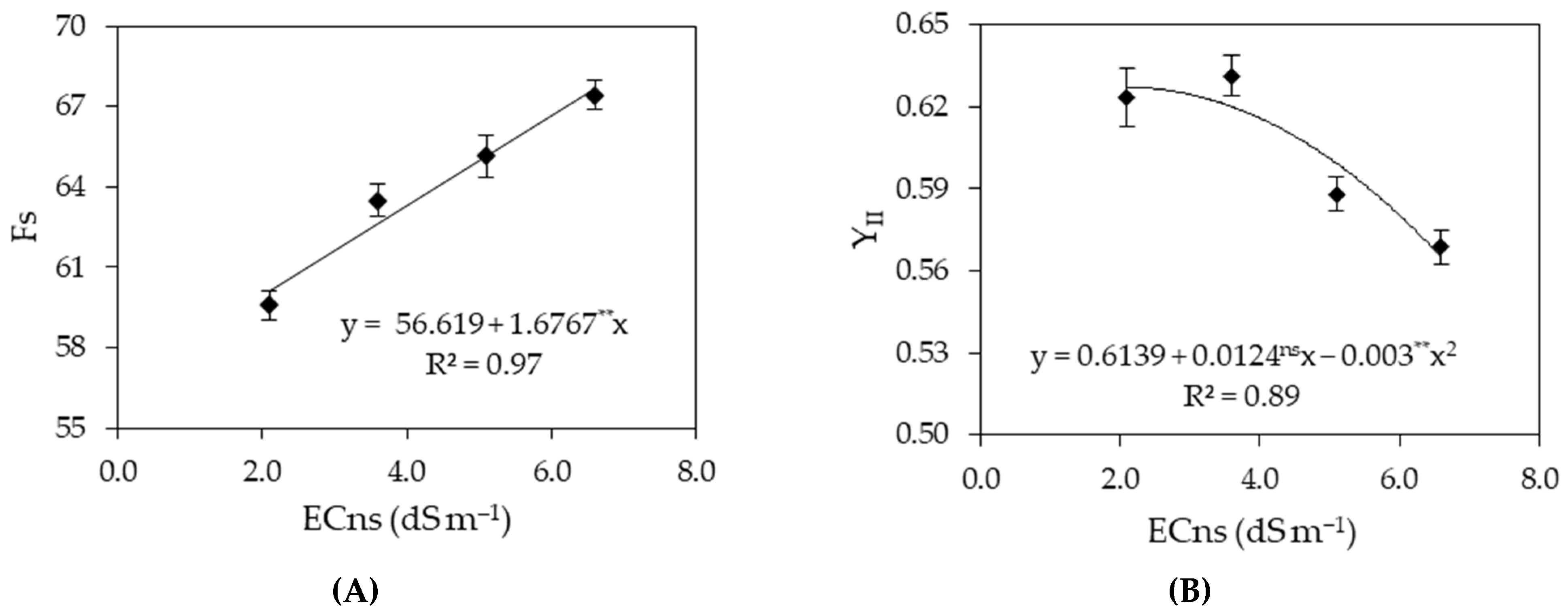
2.5.5. Chlorophyll Fluorescence
2.5.6. Production Components
2.5.7. Post-Harvest
2.6. Data Analysis
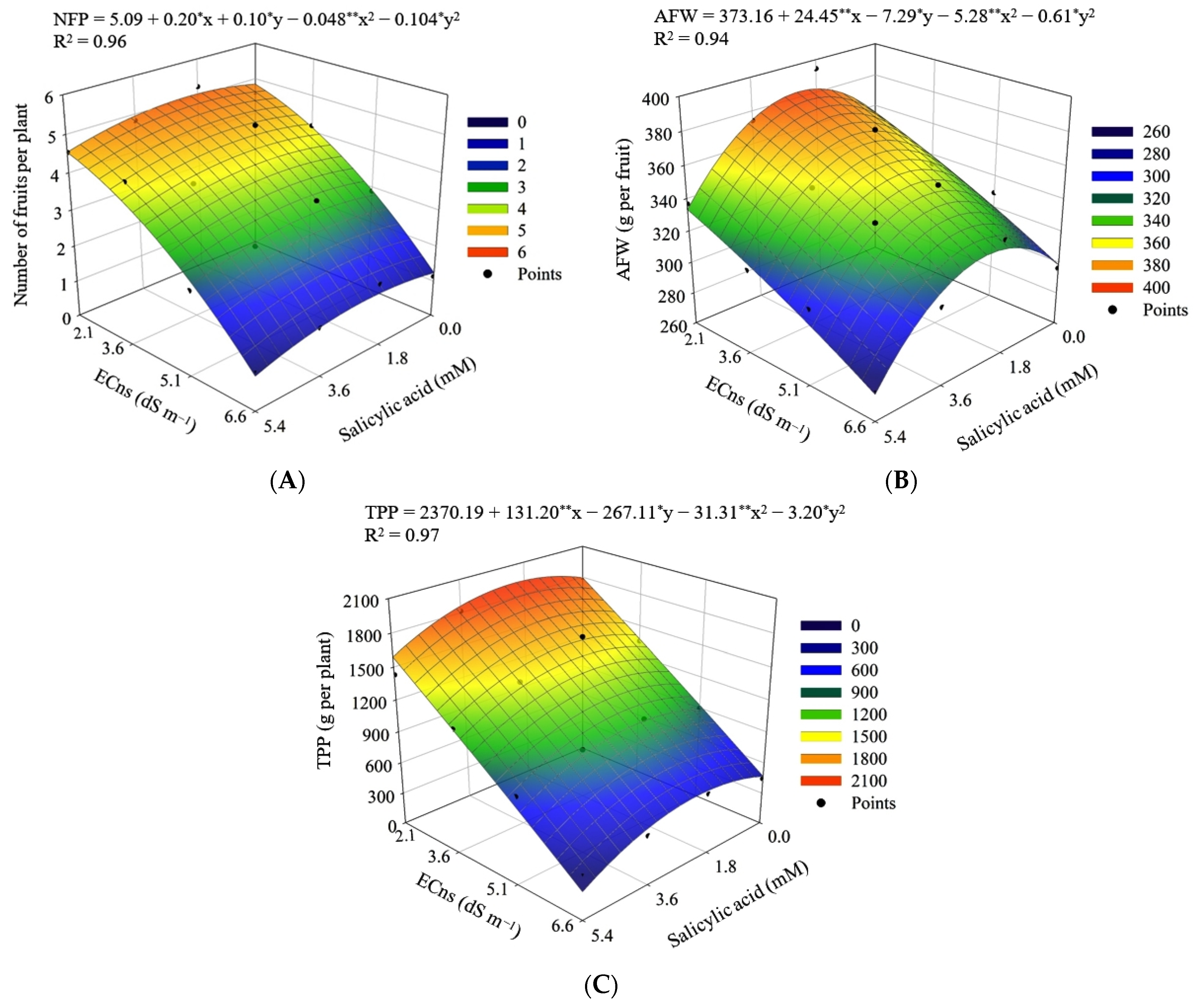
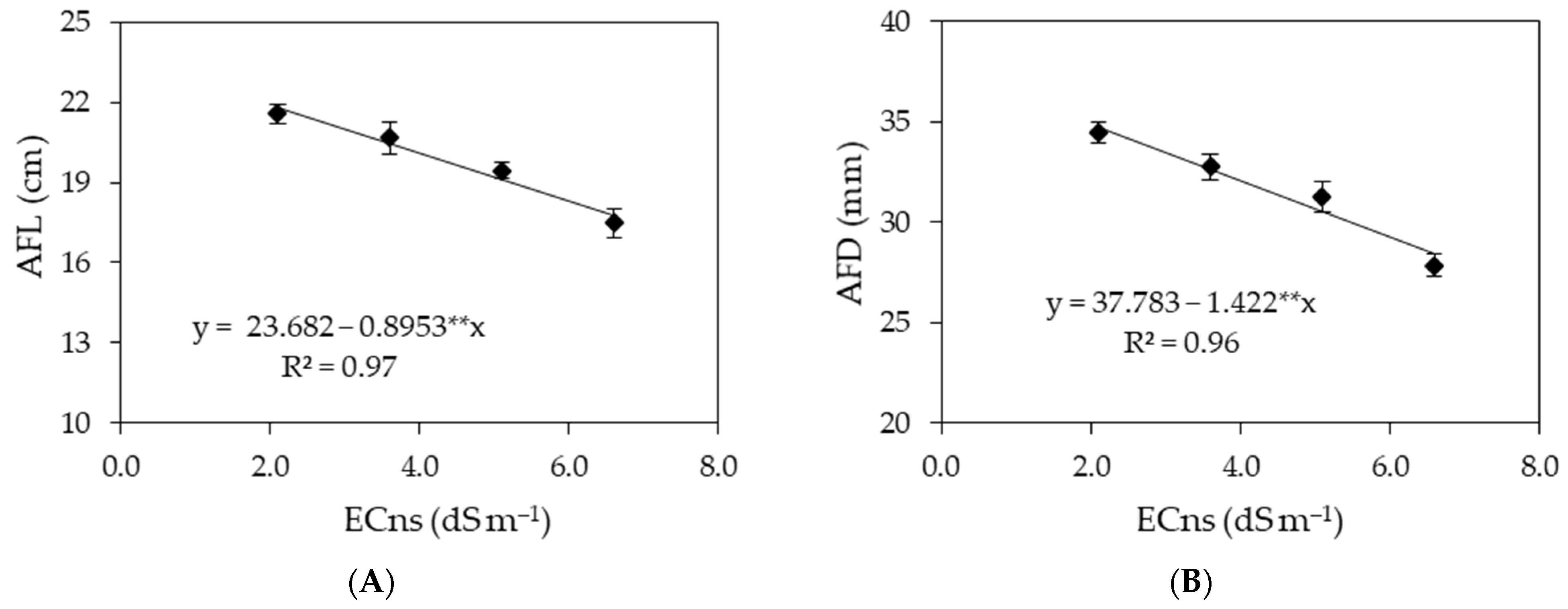
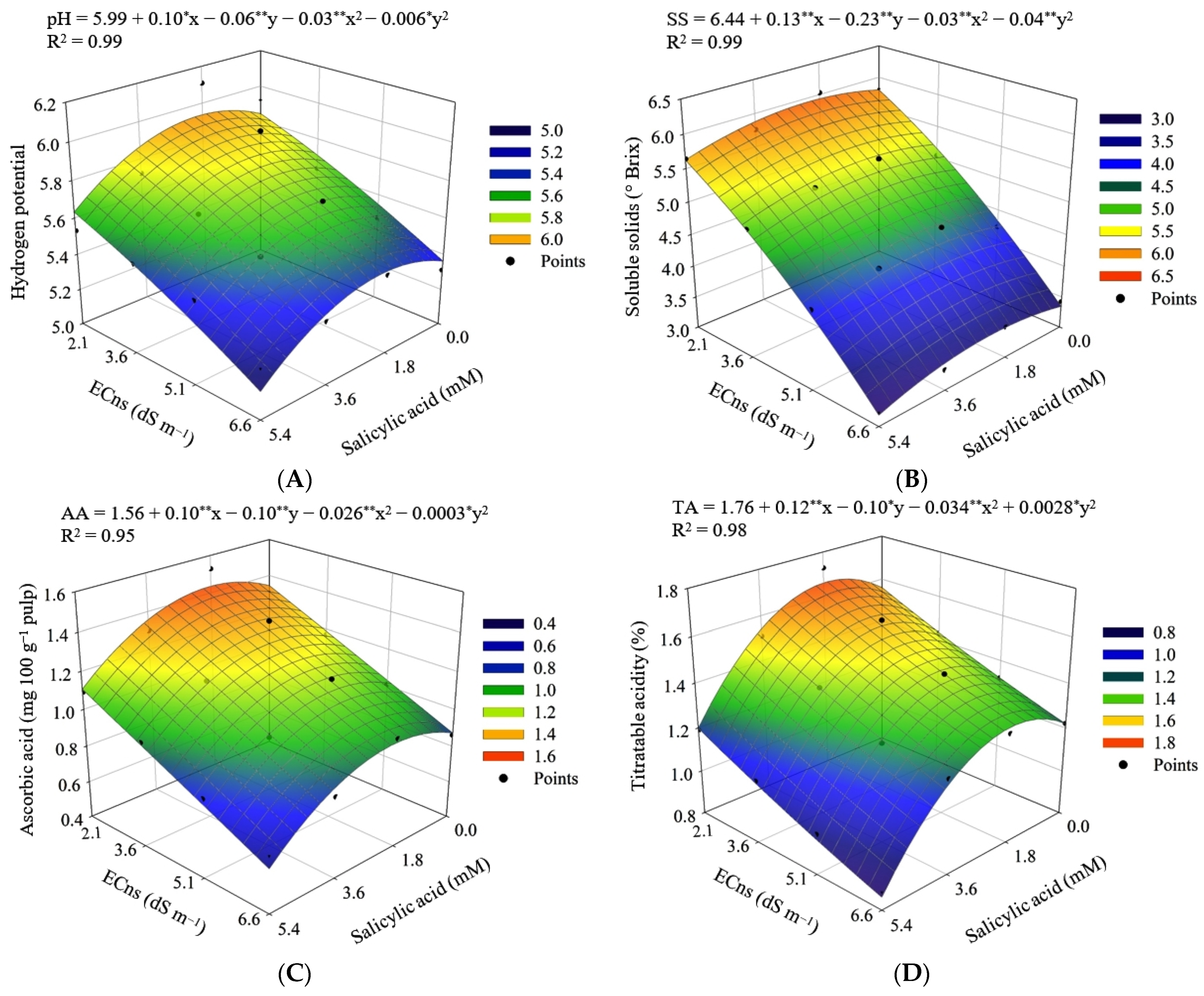
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Li, X.; Simunek, J.; Shi, H.; Chen, N.; Hu, Q.; Tian, T. Evaluating soil salt dynamics in a field drip-irrigated with brackish water and leached with freshwater during different crop growth stages. Agric. Water Manag. 2021, 244, e106601. [Google Scholar] [CrossRef]
- Zhang, W.; Du, T. Fresh/brackish watering at growth period provided a trade-off between lettuce growth and resistance to NaCl-induced damage. Sci. Hortic. 2022, 304, e111283. [Google Scholar] [CrossRef]
- Mendonça, A.J.T.; da Silva, A.A.R.; de Lima, G.S.; Soares, L.A.A.; Oliveira, V.K.N.; Gheyi, H.R.; Fernandes, P.D. Salicylic acid modulates okra tolerance to salt stress in hydroponic system. Agriculture 2022, 12, 1687. [Google Scholar] [CrossRef]
- Bachani, J.; Mahanty, A.; Aftab, T.; Kumar, K. Insight into calcium signalling in salt stress response. S. Afr. J. Bot. 2022, 151, 1–8. [Google Scholar] [CrossRef]
- Sousa, H.C.; de Sousa, G.G.; Cambissa, P.B.; Lessa, C.I.; Goes, G.F.; Silva, F.D.B.; Viana, T.V.A. Gas exchange and growth of zucchini crop subjected to salt and water stress. Rev. Bras. Eng. Agricola Ambient. 2022, 26, 815–822. [Google Scholar] [CrossRef]
- Najar, R.; Aydi, S.; Sassi-Aydi, S.; Zarai, A.; Abdelly, C. Effect of salt stress on photosynthesis and chlorophyll fluorescence in Medicago truncatula. Plant Biosyst. 2019, 153, 88–97. [Google Scholar] [CrossRef]
- Fang, S.; Hou, X.; Liang, X. Response mechanisms of plants under saline-alkali stress. Front. Plant Sci. 2021, 12, e667458. [Google Scholar] [CrossRef]
- Arif, Y.; Sami, F.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salicylic acid in relation to other phytohormones in plant: A study towards physiology and signal transduction under challenging environment. Environ. Exp. Bot. 2020, 175, 104040. [Google Scholar] [CrossRef]
- Poór, P. Effects of salicylic acid on the metabolism of mitochondrial reactive oxygen species in plants. Biomolecules 2020, 10, 341. [Google Scholar] [CrossRef]
- Horváth, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Poór, P.; Borbély, P.; Bódi, N.; Bagyánszki, M.; Tari, I. Effects of salicylic acid on photosynthetic activity and chloroplast morphology under light and prolonged darkness. Photosynthetica 2019, 57, 367–376. [Google Scholar] [CrossRef]
- Veloso, L.L.S.A.; de Lima, G.S.; da Silva, A.A.R.; Souza, L.P.; de Lacerda, C.N.; Silva, I.J.; Fernandes, P.D. Attenuation of salt stress on the physiology and production of bell peppers by treatment with salicylic acid. Semin. Ciênc. Agrar. 2021, 42, 2751–2768. [Google Scholar] [CrossRef]
- Da Silva, A.A.R.; Sousa, P.F.N.; de Lima, G.S.; Soares, L.A.A.; Gheyi, H.R.; Azevedo, C.A.V. Hydrogen peroxide reduces the effect of salt stress on growth and postharvest quality of hydroponic mini watermelon. Water Air Soil Pollut. 2022, 233, 198. [Google Scholar] [CrossRef]
- Sousa, V.F.O.; Santos, A.S.; Sales, W.S.; Silva, A.J.; Gomes, F.A.L.; Dias, T.J.; Araújo, J.R.E.S. Exogenous application of salicylic acid induces salinity tolerance in eggplant seedlings. Braz. J. Biol. 2022, 84, e257739. [Google Scholar] [CrossRef]
- Soares, M.D.M.; Dantas, M.V.; de Lima, G.S.; Oliveira, V.K.N.; Soares, L.A.A.; Gheyi, H.R.; Fernandes, P.D. Physiology and yield of ‘Gaúcho’ melon under brackish water and salicylic acid in hydroponic cultivation. Arid. Land Res. Manag. 2022, 37, 134–153. [Google Scholar] [CrossRef]
- Silva, T.I.; Silva, J.S.; Dias, M.G.; Martins, J.V.S.; Ribeiro, W.S.; Dias, T.J. Salicylic acid attenuates the harmful effects of salt stress on basil. Rev. Bras. Eng. Agricola Ambient. 2022, 26, 399–406. [Google Scholar] [CrossRef]
- Kaloterakis, N.; van Delden, S.H.; Hartley, S.; De Deyn, G.B. Silicon application and plant growth promoting rhizobacteria consisting of six pure Bacillus species alleviate salinity stress in cucumber (Cucumis sativus). Sci. Hortic. 2021, 288, e110383. [Google Scholar] [CrossRef]
- Silva, A.A.R.; Veloso, L.L.S.A.; de Lima, G.S.; Soares, L.A.A.; Chaves, L.H.G.; Silva, F.A.; Fernandes, P.D. Induction of salt stress tolerance in cherry tomatoes under different salicylic acid application methods. Semin. Cienc. Agrar. 2022, 43, 1145–1166. [Google Scholar] [CrossRef]
- Bezerra, R.R.; Júnior, J.A.S.; Pessoa, U.C.; Silva, Ê.F.d.F.e.; de Oliveira, T.F.; Nogueira, K.F.; de Souza, E.R. Water efficiency of coriander under flows of application of nutritive solutions prepared in brackish waters. Water 2022, 14, 4005. [Google Scholar] [CrossRef]
- Yildirim, E.; Turan, M.; Guvenc, I. Effect of foliar salicylic acid applications on growth, chlorophyll, and mineral content of cucumber grown under salt stress. J. Plant Nutr. 2008, 31, 593–612. [Google Scholar] [CrossRef]
- Das, A.; Singh, S.; Islam, Z.; Munshi, A.D.; Behera, T.K.; Dutta, S.; Dey, S. Current progress in genetic and genomics-aided breeding for stress resistance in cucumber (Cucumis sativus L.). Sci. Hortic. 2022, 300, e111059. [Google Scholar] [CrossRef]
- Maas, E.V.; Hoffman, G.J. Crop Salt tolerance—Current assessment. J. Irrig. Drain. Div. ASCE. 1977, 103, 115–134. [Google Scholar]
- Medeiros, P.R.; Duarte, S.N.; Dias, C.T. Tolerância da cultura do pepino à salinidade em ambiente protegido. Rev. Bras. Eng. Agricola Ambient. 2009, 13, 406–410. [Google Scholar] [CrossRef]
- Sotiroudis, G.; Melliou, E.; Sotiroudis, T.G.; Chinou, I. Chemical analysis, antioxidant and antimicrobial activity of three Greek cucumber (Cucumis sativus) cultivars. J. Food Biochem. 2010, 34, 61–78. [Google Scholar]
- Uthpala, T.G.G.; Marapana, R.A.U.J. Study on nutritional composition on firmness of two gherkin (Cucumis sativus L.) varieties (Ajax and Vlasset) on brine fermentation. Am. J. Food Sci. Technol. 2017, 5, 61–63. [Google Scholar] [CrossRef]
- Agatemor, U.M.; Nwodo, O.F.; Anosike, C.A. Phytochemical and proximate composition of cucumber (Cucumis sativus) fruit from Nsukka, Nigeria Afr. J. Biotechnol. 2018, 17, 1215–1219. [Google Scholar] [CrossRef]
- Brengi, S.H.; Abd Allah, E.M.; Abouelsaad, I.A. Effect of melatonin or cobalt on growth, yield and physiological responses of cucumber (Cucumis sativus L.) plants under salt stress. J. Saudi Soc. Agric. Sci. 2022, 21, 51–60. [Google Scholar] [CrossRef]
- Chen, T.W.; Gomez Pineda, I.M.; Brand, A.M.; Stützel, H. Determining ion toxicity in cucumber under salinity stress. Agronomy 2020, 10, 677. [Google Scholar] [CrossRef]
- Isla. Available online: https://www.isla.com.br/arquivos-para-download/catalogos (accessed on 5 November 2022).
- Medeiros, P.R.; Duarte, S.N.; Dias, C.T.; Silva, M.F. Tolerância do pepino à salinidade em ambiente protegido: Efeitos sobre propriedades físico-químicas dos frutos. Irriga 2010, 15, 301–311. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Medeiros, J.F.; Lisboa, R.A.; de Oliveira, M.; de Silva Júnior, M.J.; Alves, L.P. Caracterização das águas subterrâneas usadas para irrigação na área produtora de melão da Chapada do Apodi. Rev. Bras. Eng. Agricola Ambient. 2003, 7, 469–472. [Google Scholar] [CrossRef]
- Weatherley, P.E. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Scotti-Campos, P.; Pham-Thi, A.T.; Semedo, J.N.; Pais, I.P.; Ramalho, J.C.; Matos, M.C. Physiological responses and membrane integrity in three Vigna genotypes with contrasting drought tolerance. Emir. J. Food Agric. 2013, 25, 1002–1013. [Google Scholar] [CrossRef] [Green Version]
- Arnon, D.I. Copper enzymes in isolated chloroplasts: Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Carvalho, A.D.F.; Amaro, G.B.; Lopes, J.F.; Vilela, N.J.; Michereff Filho, M.; Andrade, R. A Cultura do Pepino; MAPA: Brasília, Brazil, 2013; Volume 1, 18p. [Google Scholar]
- Instituto Adolfo Lutz—IAL. Normas analíticas do Instituto Adolfo Lutz. In Métodos Químicos e Físicos Para Análise de Alimentos, 3rd ed.; IMESP: São Paulo, Brazil, 2008; p. 1020. [Google Scholar]
- Ferreira, D.F. SISVAR: A computer analysis system to fixed effects split plot type designs. Rev. Bras. Biom. 2019, 37, 529–535. [Google Scholar] [CrossRef]
- Cavalcante, E.S.; Lacerda, C.F.D.; Costa, R.N.T.; Gheyi, H.R.; Pinho, L.L.; Bezerra, F.M.S.; Canjá, J.F. Supplemental irrigation using brackish water on maize in tropical semi-arid regions of Brazil: Yield and economic analysis. Sci. Agric. 2021, 78, e20200151. [Google Scholar] [CrossRef]
- Dantas, M.V.; de Lima, G.S.; Gheyi, H.R.; Pinheiro, F.W.A.; Silva, P.C.C.; Soares, L.A.A. Gas exchange and hydroponic production of zucchini under salt stress and H2O2 application. Rev. Caatinga. 2022, 35, 436–449. [Google Scholar] [CrossRef]
- Yadav, M.; Gupta, P.; Seth, C.S. Foliar application of α-lipoic acid attenuates cadmium toxicity on photosynthetic pigments and nitrogen metabolism in Solanum lycopersicum L. Acta Physiol. Plant. 2022, 44, 112. [Google Scholar] [CrossRef]
- Roque, I.A.; Soares, L.A.A.; de Lima, G.S.; Lopes, I.A.P.; Silva, L.A.; Fernandes, P.D. Biomass, gas exchange and production of cherry tomato cultivated under saline water and nitrogen fertilization. Rev. Caatinga 2022, 35, 686–696. [Google Scholar] [CrossRef]
- Ali, M.; Kamran, M.; Abbasi, G.H.; Saleem, M.H.; Ahmad, S.; Parveen, A.; Fahad, S. Melatonin-induced salinity tolerance by ameliorating osmotic and oxidative stress in the seedlings of two tomato (Solanum lycopersicum L.) cultivars. J. Plant Growth Regul. 2021, 40, 2236–2248. [Google Scholar] [CrossRef]
- Rady, M.M.; Belal, H.E.E.; Gadallah, F.M.; Semida, W.M. Selenium application in two methods elevates drought tolerance in Solanum lycopersicum by increasing yield, quality, and antioxidant defense system and suppressing oxidative stress biomarkers. Sci. Hortic. 2020, 266, e109290. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [PubMed]
- Sullivan, C.Y. Mechanisms of heat drought resistence in grain sorghum and methods of measurement. In Sorghum in Seventies; Rao, N.G.P., House, L.R., Eds.; Oxford and IBH Publication: New Delhi, India, 1971; Volume 1, 247p. [Google Scholar]
- Oliveira, V.K.N.; de Lima, G.S.; Soares, M.D.M.; Soares, L.A.A.; Gheyi, H.R.; Silva, A.A.R.; Fernandes, P.D. Salicylic acid does not mitigate salt stress on the morphophysiology and production of hydroponic melon. Braz. J. Biol. 2022, 82, e262664. [Google Scholar] [CrossRef] [PubMed]
- Xavier, A.V.; de Lima, G.S.; Gheyi, H.R.; da Silva, A.A.R.; de Lacerda, C.N.; Soares, L.A.A.; Fernandes, P.D. Salicylic acid alleviates salt stress on guava plant physiology during rootstock formation. Rev. Bras. Eng. Agrícola e Ambient. 2022, 26, 855–862. [Google Scholar] [CrossRef]
- Gharbi, E.; Martínez, J.P.; Benahmed, H.; Fauconnier, M.L.; Lutts, S.; Quinet, M. Salicylic acid differently impacts ethylene and polyamine synthesis in the glycophyte Solanum lycopersicum and the wild-related halophyte Solanum chilense exposed to mild salt stress. Physiol. Plant. 2016, 158, 152–167. [Google Scholar] [CrossRef]
- Jini, D.; Joseph, B. Physiological mechanism of salicylic acid for alleviation of salt stress in rice. Rice Sci. 2017, 24, 97–108. [Google Scholar] [CrossRef]
- Roumani, A.; Biabani, A.; Karizaki, A.R.; Alamdari, E.G. Foliar salicylic acid application to mitigate the effect of drought stress on isabgol (Plantago ovata forssk). Biochem. Syst. Ecol. 2022, 104, e104453. [Google Scholar] [CrossRef]
- Dias, A.S.; de Lima, G.S.; Sá, F.V.S.; Gheyi, H.R.; Soares, L.A.A.; Fernandes, P.D. Gas exchanges and photochemical efficiency of West Indian cherry cultivated with saline water and potassium fertilization. Rev. Bras. Eng. Agricola Ambient. 2018, 22, 628–633. [Google Scholar] [CrossRef]
- Gurmani, A.R.; Khan, S.U.; Ali, A.; Rubab, T.; Schwinghamer, T.; Jilani, G.; Zhang, J. Salicylic acid and kinetin mediated stimulation of salt tolerance in cucumber (Cucumis sativus L.) genotypes varying in salinity tolerance. HEB 2018, 59, 461–471. [Google Scholar] [CrossRef]
- Turan, M.; Ekinci, M.; Kul, R.; Boynueyri, F.G.; Yildirim, E. Mitigation of salinity stress in cucumber seedlings by exogenous hydrogen sulfide. J. Plant Res. 2022, 135, 517–529. [Google Scholar] [CrossRef]
- Lee, S.Y.; Damodaran, P.N.; Roh, K.S. Influence of salicylic acid on rubisco and rubisco activase in tobacco plant grown under sodium chloride in vitro. Saudi J. Biol. Sci. 2014, 21, 417–426. [Google Scholar] [CrossRef]
- Costa, A.A.; Paiva, E.P.; Torres, S.B.; Souza Neta, M.L.; Pereira, K.T.O.; Leite, M.S.; Benedito, C.P. Osmoprotection in Salvia hispanica L. seeds under water stress attenuators. Braz. J. Biol. 2021, 82, e233547. [Google Scholar] [CrossRef] [PubMed]
- Nigam, B.; Dubey, R.S.; Rathore, D. Protective role of exogenously supplied salicylic acid and PGPB (Stenotrophomonas sp.) on spinach and soybean cultivars grown under salt stress. Sci. Hortic. 2022, 293, e110654. [Google Scholar] [CrossRef]
- Kang, G.; Li, G.; Guo, T. Molecular mechanism of salicylic acid-induced abiotic stress tolerance in higher plants. Acta Physiol. Plant. 2014, 36, 2287–2297. [Google Scholar] [CrossRef]
- Hundare, A.; Joshi, V.; Joshi, N. Salicylic acid attenuates salinity-induced growth inhibition in in vitro raised ginger (Zingiber officinale Roscoe) plantlets by regulating ionic balance and antioxidative system. Plant Stress. 2022, 4, e100070. [Google Scholar] [CrossRef]
- Lamnai, K.; Anaya, F.; Fghire, R.; Zine, H.; Janah, I.; Wahbi, S.; Loutfi, K. Combined effect of salicylic acid and calcium application on salt-stressed strawberry plants. Russ. J. Plant Physiol. 2022, 69, 12. [Google Scholar] [CrossRef]
- Da Silva, A.A.R.; de Lima, G.S.; de Azevedo, C.A.V.; Gheyi, H.R.; Soares, L.A.A.; Veloso, L.L.S.A. Salicylic acid improves physiological indicators of soursop irrigated with saline water. Rev. Bras. Eng. Agricola Ambient. 2022, 26, 412–419. [Google Scholar] [CrossRef]
- Veloso, L.L.S.A.; de Azevedo, C.A.V.; Nobre, R.G.; de Lima, G.S.; de Capitulino, J.D.; Silva, F.A. H2O2 alleviates salt stress effects on photochemical efficiency and photosynthetic pigments of cotton genotypes. Rev. Bras. Eng. Agricola Ambient. 2023, 27, 34–41. [Google Scholar] [CrossRef]
- Silva, M.M.P.; Vasquez, H.M.; Bressan-Smith, R.; da Silva, J.F.C.; Erbesdobler, E.D.A.; Andrade Junior, P.S.C. Eficiência fotoquímica de gramíneas forrageiras tropicais submetidas à deficiência hídrica. Rev. Bras. Zootec. 2006, 35, 67–74. [Google Scholar] [CrossRef]
- Tatagiba, S.D.; Moraes, G.A.B.K.; Nascimento, K.J.T.; Peloso, A.F. Limitações fotossintéticas em folhas de plantas de tomateiro submetidas a crescentes concentrações salinas. Eng. Agri. 2014, 22, 138–149. [Google Scholar]
- Sá, F.V.S.; Gheyi, H.R.; de Lima, G.S.; de Paiva, E.P.; Silva, L.A.; Moreira, R.C.L.; Dias, A.S. Ecophysiology of West Indian cherry irrigated with saline water under phosphorus and nitrogen doses. Biosci. J. 2019, 35, 211–221. [Google Scholar] [CrossRef]
- Larbi, A.; Baccar, R.; Boulal, H. Response of olive tree to ammonium nitrate fertilization under saline conditions. J. Plant Nutr. 2020, 44, 1432–1445. [Google Scholar] [CrossRef]
- da Silva, J.S.; Sá, F.V.D.S.; Dias, N.D.S.; Neto, M.F.; Jales, G.D.; Fernandes, P.D. Morphophysiology of mini watermelon in hydroponic cultivation using reject brine and substrates. Rev. Bras. Eng. Agrícola Ambient. 2021, 25, 402–408. [Google Scholar] [CrossRef]
- Agnihotri, A.; Gupta, P.; Dwivedi, A.; Seth, C.S. Counteractive mechanism (s) of salicylic acid in response to lead toxicity in Brassica juncea (L.) Czern. cv. Varuna. Planta 2018, 248, 49–68. [Google Scholar] [CrossRef]
- Gupta, S.; Seth, C.S. Salicylic acid alleviates chromium (VI) toxicity by restricting its uptake, improving photosynthesis and augmenting antioxidant defense in Solanum lycopersicum L. Physiol. Mol. Biol. Plants 2021, 27, 2651–2664. [Google Scholar] [CrossRef] [PubMed]
- Ó, L.M.G.D.; Cova, A.M.W.; Neto, A.D.D.A.; da Silva, N.D.; Silva, P.C.C.; Santos, A.L.; Gheyi, H.R.; da Silva, L.L. Osmotic adjustment, production, and post-harvest quality of mini watermelon genotypes differing in salt tolerance. Sci. Hort. 2022, 306, e111463. [Google Scholar] [CrossRef]
- Modesto, F.J.N.; dos Santos, M.Â.C.M.; Soares, T.M.; Santos, E.P.M. Crescimento, produção e consumo hídrico do quiabeiro submetido à salinidade em condições hidropônicas. Irriga 2019, 24, 2486–2497. [Google Scholar] [CrossRef]
- Abbasi, F.; Khaleghi, A.; Khadivi, A. The effect of salicylic acid on physiological and morphological traits of cucumber (Cucumis sativus L. cv. Dream). Gesunde Pflanz. 2020, 72, 155–162. [Google Scholar] [CrossRef]
- Bione, M.A.A.; Soares, T.M.; Cova, A.M.W.; Silva Paz, V.P.; Gheyi, H.R.; Rafael, M.R.S.; Neves, B.S.L. Hydroponic production of ‘Biquinho’ pepper with brackish water. Agric. Water Manag. 2021, 245, e106607. [Google Scholar] [CrossRef]
- Aires, E.S.; Ferraz, A.K.L.; Carvalho, B.L.; Teixeira, F.P.; Rodrigues, J.D.; Ono, E.O. Foliar application of salicylic acid intensifies antioxidant system and photosynthetic efficiency in tomato plants. Bragantia 2022, 81, e1522. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [PubMed]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How can salicylic acid and jasmonic acid mitigate salt toxicity in soybean plants? Ecotoxicol. Environ. Saf. 2018, 147, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Seymen, M.; Yavuz, D.; Ercan, M.; Akbulut, M.; Çoklar, H.; Kurtar, E.S.; Türkmen, Ö. Effect of wild watermelon rootstocks and water stress on chemical properties of watermelon fruit. HEB 2021, 62, 411–422. [Google Scholar] [CrossRef]
- De Lima, G.S.; Pinheiro, F.W.A.; Gheyi, H.R.; Soares, L.A.D.A.; Silva, S.S. Growth and post-harvest fruit quality of West Indian cherry under saline water irrigation and potassium fertilization. Rev. Caatinga 2020, 33, 775–784. [Google Scholar] [CrossRef]
- Blankenship, S.M.; Dole, J.M. 1-Methylcyclopropene: A review. Postharvest Biol. Technol. 2003, 28, 1–25. [Google Scholar] [CrossRef]
- Brasil, A.S.; Sigarini, K.D.S.; Pardinho, F.C.; Faria, R.; Siqueira, N.F.M.P. Avaliação da qualidade físico-química de polpas de fruta congeladas comercializadas na cidade de Cuiabá MT. Rev. Bras. Fruticultura 2016, 38, 167–175. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ECns (dS m−1) | NaCl | CaCl2.2H2O | MgCl2.6H2O |
|---|---|---|---|
| g | |||
| 2.1 | - | - | - |
| 3.6 | 61.43 | 20.55 | 15.23 |
| 5.1 | 122.85 | 41.10 | 30.45 |
| 6.6 | 184.28 | 61.65 | 45.68 |
| Source of Variation | DF | Mean Squares | |||||
|---|---|---|---|---|---|---|---|
| RWC | % EL | Ci | gs | E | A | ||
| Saline nutrient solution (ECns) | 3 | 1607.72 ** | 106.41 ** | 19,058.13 ** | 0.012 ** | 2.93 ** | 237.04 ** |
| Linear regression | 1 | 4218.93 ** | 307.07 ** | 51,606.70 ** | 0.03 ** | 7.64 ** | 685.88 ** |
| Quadratic regression | 1 | 601.59 ns | 8.19 * | 3835.94 * | 0.001 ns | 0.79 ns | 25.11 ns |
| Residual 1 | 9 | 24.42 | 0.06 | 180.69 | 2.0 × 10−6 | 0.01 | 0.05 |
| Salicylic acid (SA) | 3 | 11.26 ns | 136.83 ** | 4900.61 ** | 0.002 ** | 0.38 ** | 130.82 ** |
| Linear regression | 1 | - | 60.31 * | 1070.47 ns | 0.001 * | 0.07 * | 141.21 * |
| Quadratic regression | 1 | - | 223.59 ** | 11,184.12 ** | 0.004 ** | 1.04 ** | 165.86 ** |
| Interaction (ECns × SA) | 9 | 107.06 * | 0.33 ** | 1300.44 * | 6.0 × 10−6 * | 3.52 ** | 10.86 ** |
| Residual 2 | 36 | 29.07 | 3.40 | 415.59 | 5.7 × 10−4 | 0.28 | 0.64 |
| CV 1 (%) | 6.62 | 3.66 | 5.93 | 10.55 | 6.32 | 5.81 | |
| CV 2 (%) | 7.23 | 5.10 | 8.99 | 13.89 | 12.32 | 9.83 | |
| Source of Variation | DF | Mean Squares | |||
|---|---|---|---|---|---|
| Chl a | Chl b | Chl t | Car | ||
| Saline nutrient solution (ECns) | 3 | 54.47 ** | 27.09 ** | 154.76 ** | 1.90 ** |
| Linear regression | 1 | 158.72 ** | 67.54 ** | 433.61 ** | 5.41 ** |
| Quadratic regression | 1 | 4.16 ns | 12.06 ns | 30.36 ns | 0.26 ns |
| Residual 1 | 9 | 3.94 | 0.59 | 6.59 | 0.62 |
| Salicylic acid (SA) | 3 | 0.49 ns | 1.26 ns | 9.73 ns | 1.01 ns |
| Interaction (ECns × SA) | 9 | 5.42 * | 0.76 ns | 18.33 * | 0.29 ns |
| Residual 2 | 36 | 3.24 | 1.20 | 7.52 | 0.38 |
| CV 1 (%) | 10.87 | 12.02 | 10.43 | 13.65 | |
| CV 2 (%) | 9.87 | 17.19 | 11.14 | 10.48 | |
| Source of Variation | DF | Mean Squares | ||||||
|---|---|---|---|---|---|---|---|---|
| F0 | Fm | Fv | Fv/Fm | Fs | YII | ETR | ||
| Saline nutrient solution (ECns) | 3 | 16,234.27 ** | 878,366.35 ** | 889,782.38 ** | 0.021 ** | 174.32 ** | 0.017 ** | 120.39 ns |
| Linear regression | 1 | 47,458.15 ** | 225,506.26 ** | 20,197.31 ** | 0.028 * | 505.99 ** | 0.006 ns | - |
| Quadratic regression | 1 | 669.52 ns | 73,902.41 ns | 178,325 ns | 0.034 ** | 10.51 ns | 0.037 ** | - |
| Residual 1 | 9 | 1368.75 | 46,782.09 | 178.25 | 72.0 × 10−6 | 4.73 | 0.001 | 53.01 |
| Salicylic acid (SA) | 3 | 1906.64 ns | 44,209.09 ns | 695.02 ns | 25.6 × 10−5 ns | 93.97 ns | 0.0001 ns | 19.61 ns |
| Interaction (ECns × SA) | 9 | 2315.81 ns | 113,099.59 ns | 26,025.02 ns | 0.007 ** | 37.07 ns | 0.002 ns | 62.11 ns |
| Residual 2 | 36 | 2395.35 | 64,148.96 | 15,211.43 | 11.8 × 10−5 | 5.87 | 0.001 | 31.74 |
| CV 1 (%) | 6.44 | 9.73 | 9.94 | 10.21 | 5.41 | 5.94 | 12.94 | |
| CV 2 (%) | 8.51 | 11.40 | 8.66 | 11.42 | 6.79 | 5.08 | 10.01 | |
| Source of Variation | DF | Mean Squares | ||||
|---|---|---|---|---|---|---|
| NF | AFW | TPP | AFL | AFD | ||
| Saline nutrient solution (ECns) | 3 | 40.18 ** | 13,910.47 ** | 5,869,190.94 ** | 49.47 ** | 126.64 ** |
| Linear regression | 1 | 117.83 ** | 40,951.25 ** | 16,913,963.71 ** | 143.91 ** | 364.10 ** |
| Quadratic regression | 1 | 2.71 ns | 481.58 ns | 409,158.52 ns | 4.36 * | 11.78 ns |
| Residual 1 | 9 | 0.47 | 753.89 | 184,044.58 | 0.64 | 2.42 |
| Salicylic acid (SA) | 3 | 0.17 ns | 527.18 ns | 66,638.93 ns | 3.88 ns | 2.62 ns |
| Interaction (ECns × SA) | 9 | 1.67 * | 5877.88 ** | 543,499.86 * | 1.01 ns | 1.60 ns |
| Residual 2 | 36 | 0.42 | 3054.67 | 89,665.21 | 0.63 | 2.79 |
| CV 1 (%) | 21.29 | 8.44 | 19.76 | 4.63 | 4.93 | |
| CV 2 (%) | 19.42 | 16.98 | 17.75 | 3.89 | 5.29 | |
| Source of Variation | DF | Mean Squares | |||
|---|---|---|---|---|---|
| pH | SS | AA | TA | ||
| Saline nutrient solution (ECns) | 3 | 0.53 ** | 17.97 ** | 0.73 ** | 0.41 ** |
| Linear regression | 1 | 1.56 ** | 52.91 ** | 1.99 ** | 1.16 ** |
| Quadratic regression | 1 | 0.01 ns | 0.99 ns | 0.16 ns | 0.005 ns |
| Residual 1 | 9 | 0.03 | 0.004 | 0.006 | 0.001 |
| Salicylic acid (SA) | 3 | 0.35 ** | 0.009 ns | 0.35 ** | 0.65 ** |
| Linear regression | 1 | 0.17 ns | - | 0.49 * | 0.69 * |
| Quadratic regression | 1 | 0.55 ** | - | 0.45 ** | 1.21 ** |
| Interaction (ECns × SA) | 9 | 0.13 * | 0.45 ** | 0.006 ** | 0.04 * |
| Residual 2 | 36 | 0.02 | 0.05 | 0.002 | 0.008 |
| CV 1 (%) | 3.24 | 3.42 | 2.27 | 2.74 | |
| CV 2 (%) | 3.00 | 4.62 | 4.86 | 6.75 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, V.K.N.; Silva, A.A.R.d.; Lima, G.S.d.; Soares, L.A.d.A.; Gheyi, H.R.; de Lacerda, C.F.; Vieira de Azevedo, C.A.; Nobre, R.G.; Garófalo Chaves, L.H.; Dantas Fernandes, P.; et al. Foliar Application of Salicylic Acid Mitigates Saline Stress on Physiology, Production, and Post-Harvest Quality of Hydroponic Japanese Cucumber. Agriculture 2023, 13, 395. https://doi.org/10.3390/agriculture13020395
Oliveira VKN, Silva AARd, Lima GSd, Soares LAdA, Gheyi HR, de Lacerda CF, Vieira de Azevedo CA, Nobre RG, Garófalo Chaves LH, Dantas Fernandes P, et al. Foliar Application of Salicylic Acid Mitigates Saline Stress on Physiology, Production, and Post-Harvest Quality of Hydroponic Japanese Cucumber. Agriculture. 2023; 13(2):395. https://doi.org/10.3390/agriculture13020395
Chicago/Turabian StyleOliveira, Valeska Karolini Nunes, André Alisson Rodrigues da Silva, Geovani Soares de Lima, Lauriane Almeida dos Anjos Soares, Hans Raj Gheyi, Claudivan Feitosa de Lacerda, Carlos Alberto Vieira de Azevedo, Reginaldo Gomes Nobre, Lúcia Helena Garófalo Chaves, Pedro Dantas Fernandes, and et al. 2023. "Foliar Application of Salicylic Acid Mitigates Saline Stress on Physiology, Production, and Post-Harvest Quality of Hydroponic Japanese Cucumber" Agriculture 13, no. 2: 395. https://doi.org/10.3390/agriculture13020395