
Genetic Load of Mutations Causing Inherited Diseases and Its Classification in Dairy Cattle Bred in the Russian Federation
 , and
, and 
Abstract
:1. Introduction
2. What Does Genetic Load Mean?
3. Mutations Leading to Disorders in Cattle Breeds
4. Mutation Occurrence in the World and Russia
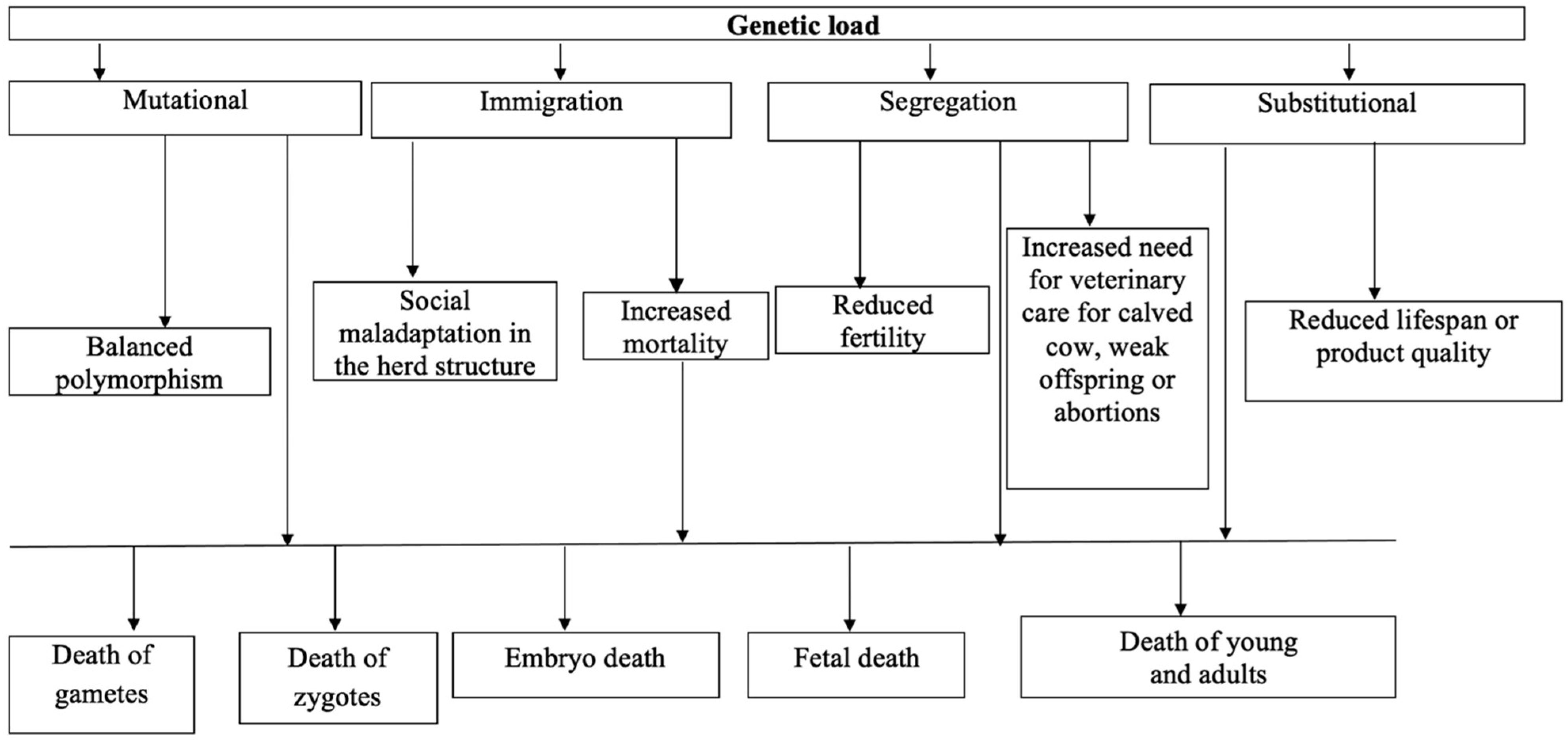
5. Classification of Genetic Load
5.1. Mutational Genetic-Load
5.2. Immigration Genetic-Load
5.3. Segregation Genetic-Load
5.4. Substitutional Genetic-Load
6. Integrating Genomic Approaches to Improve Dairy Cattle
7. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Haldane, J.B.S. The cost of natural selection. J. Genet. 1957, 55, 511–524. [Google Scholar] [CrossRef]
- Marzanov, N.S.; Devrishov, D.A.; Marzanova, S.N. Genetic Burden of Mutations in Cattle Breeds. In Materials of the International Scientific and Practical Conference on Molecular Genetic Technologies for Analysis of Gene Expression Related to Animal Productivity and Disease Resistance, Moscow, Russia, 21–22 November 2019; Kochish, I.I., Romanov, M.N., Surai, P.F., Nikonov, I.N., Selina, M.V., Eds.; Sel’skokhozyaistvennye tekhnologii: Moscow, Russia, 2019; pp. 124–130. Available online: https://elibrary.ru/item.asp?id=41444945 (accessed on 19 January 2022).
- Marzanov, N.S.; Devrishov, D.A.; Abylkasymov, D.A.; Marzanova, S.N.; Konovalova, N.V.; Libet, I.S.; Novikova, L.F.; Popov, A.N.; Turbina, I.S.; Marzanova, L.K. Occurrence of β-CNA2 and β-CNA1 Alleles at the Beta-casein Locus in Cattle. In Increasing the Competitiveness of Animal Husbandry and Staffing Tasks, Proceedings of the 25th International Scientific and Practical Conference, Podolsk, Bykovo, Moscow Oblast, Russia, 24–25 June 2019; Pyzhov, A.P., Zhukov, V.F., Ponomarev, N.V., Pyzhova, E.A., Eds.; Russian Academy of Livestock Management: Podolsk, Moscow Oblast, Russia, 2019; pp. 134–143. Available online: https://elibrary.ru/item.asp?id=38589459 (accessed on 19 January 2022).
- Marzanova, S.N.; Nagorniy, V.A.; Devrishov, D.A.; Alekseev, Y.I.; Konovalova, N.V.; Tokhov, M.K.; Eskin, G.V.; Turbina, I.S.; Lukashina, A.A.; Marzanov, N.S. Founder effect and genogeography of CV and BL mutations in Black and White cattle. Russ. Agric. Sci. 2016, 42, 90–93. [Google Scholar] [CrossRef]
- Marzanova, S.N.; Devrishov, D.A.; Alekseev, J.I.; Konovalova, N.V.; Marzanov, N.S. Set of Sequence of Primers and Allele-specific Probes for Simultaneous Gene Diagnostics of Two Mutant Alleles Causing CVM and BLAD in Cattle. Proprietor: LLC “Agrovet”. Patent for Invention RU 2 577 990 C1. Federal Service for Intellectual Property, Moscow, Russia. Applied 11 February 2015. Published 20.03.2016b. Bull. No. 8. Available online: https://patents.google.com/patent/RU2577990C1/en (accessed on 19 January 2022).
- Marzanova, S.N.; Devrishov, D.A.; Alekseev, Y.I.; Konovalova, N.V.; Marzanov, N.S. Set of Primers Sequence and Allele-Specific Probes for Simultaneous Four Mutant Kappa-Casein Alleles Gene Diagnostics in Bovine Cattle. Proprietor: LLC “Agrovet”. Patent for Invention RU 2 646 140 C1. Federal Service for Intellectual Property, Moscow, Russia. Applied 24 March 2017. Published 01.03.2018a. Bull. No. 7. Available online: https://patents.google.com/patent/RU2646140C1/en (accessed on 19 January 2022).
- Marzanova, S.N.; Devrishov, D.A.; Alekseev, Y.I.; Konovalova, N.V.; Marzanov, N.S. Method for Genetic Diagnosis of Mutant Allele That Causes Short Spine Syndrome or Brachyspine Syndrome in Cattle and a Test System for Implementation Thereof. Proprietor: LLC “Agrovet”. Patent for Invention RU 2 664 454 C2. Federal Service for Intellectual Property, Moscow, Russia. Applied 4 April 2016. Published 17.08.2018b. Bull. No. 23. Available online: https://patents.google.com/patent/RU2664454C2/en (accessed on 19 January 2022).
- Marzanova, S.N.; Devrishov, D.A.; Alekseev, Y.I.; Konovalova, N.V.; Marzanov, N.S. Method for Simultaneous Genodiagnostic of Four Mutant Alleles of Kappa-Casein in Cattle and Test System for Implementation Thereof. Proprietor: LLC “Agrovet”. Patent for Invention RU 2 691 995 C2. Federal Service for Intellectual Property, Moscow, Russia. Applied 2 March 2017. Published 19.06.2019. Bull. No. 17. Available online: https://patents.google.com/patent/RU2691995C2/en (accessed on 19 January 2022).
- Lien, S.; Aleström, P.; Klungland, H.; Rogne, S. Detection of multiple β-casein (CASB) alleles by amplification created restriction sites (ACRS). Anim. Genet. 1992, 23, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Dinç, H. Genotyping of Beta-casein, Kappa-casein and Beta-lactoglobulin Genes in Turkish Native Cattle Breeds and Efforts to Delineate BCM-7 on Human PBMC. Ph.D. Thesis, Graduate School of Natural and Applied Sciences, Middle East Technical University, Ankara, Turkey. Available online: https://open.metu.edu.tr/handle/11511/18963 (accessed on 19 January 2022).
- Bertorelle, G.; Raffini, F.; Bosse, M.; Bortoluzzi, C.; Iannucci, A.; Trucchi, E.; Morales, H.E.; van Oosterhout, C. Genetic load: Genomic estimates and applications in non-model animals. Nat. Rev. Genet. 2022, 23, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Altukhov, Y.P. Genetic Processes in Populations, 3rd ed.; Akademkniga: Moscow, Russia, 2003; ISBN 5-94628-083-X. Available online: https://search.rsl.ru/en/search#q=ISBN%205-94628-083-X (accessed on 19 January 2022).
- Mutovin, G.R. Clinical Genetics. Genomics and Proteomics of Hereditary Pathology: Textbook, 3rd ed.; GEOTAR-Media: Moscow, Russia, 2010; ISBN 978-5-9704-1152-0. Available online: https://search.rsl.ru/en/search#q=ISBN%20978-5-9704-1152-0 (accessed on 19 January 2022).
- Farrell, H.M., Jr.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the proteins of cows’ milk—Sixth revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Online Mendelian Inheritance in Animals—OMIA. Sydney School of Veterinary Science. Available online: https://omia.org/ (accessed on 19 January 2022).
- Zhigachev, A.I.; Ernst, L.K.; Bogachev, A.S. About accumulation of mutation weight in the cattle breeds as result of intensive reproduction technology and improvement on target determinants. Sel’skokhozyaistvennaya Biol. (Agr. Biol.) 2008, 43, 25–32. Available online: https://www.elibrary.ru/item.asp?id=11709790 (accessed on 19 January 2022).
- Schütz, E.; Scharfenstein, M.; Brenig, B. Implication of complex vertebral malformation and bovine leukocyte adhesion deficiency DNA-based testing on disease frequency in the Holstein population. J. Dairy Sci. 2008, 91, 4854–4859. [Google Scholar] [CrossRef]
- Marzanova, S.N. Development of Gene Diagnostics of the Complex of Spinal Anomalies [CVM] and Immunodeficiency [BLAD] in Black-and-White Holstein Cattle. Ph.D. Thesis, K.I. Skryabin Moscow State Academy of Veterinary Medicine and Biotechnology, Moscow, Russia, 2012. Available online: https://search.rsl.ru/ru/record/01005529649 (accessed on 19 January 2022).
- Marzanova, S.N.; Devrishov, D.A.; Turbina, I.S.; Alekseev, Y.I.; Konovalova, N.V.; Marzanov, N.S. Genetic diagnostics of FANCIBY mutation in representatives of the Holstein breed and its crosses. Biotekhnologiya 2021, 37, 16–25. [Google Scholar] [CrossRef]
- Turbina, I.S. Characterization of Sires by Various Genetic Markers. Ph.D. Thesis, Russian Academy of Livestock Management, Moscow, Russia, 2006. [Google Scholar]
- Marzanova, S.N.; Devrishov, D.A.; Turbina, I.S.; Marzanov, N.S. DNA Technology and estimation of drift of mutant alleles in populations of the Holstein breed and its crosses. Russ. J. Genet. 2022, 58, 876–879. [Google Scholar] [CrossRef]
- Chun, H.C.; Yi, M.C.; Kuo, H.L. Genotype screening of bovine brachyspina in Taiwan Holstein cows. Am. J. Anim. Vet. Sci. 2020, 15, 206–210. [Google Scholar] [CrossRef]
- Boichard, D.; Amigue, Y. Investigation on possible linkage between the BLAD gene and a QTL for production or type traits in Holstein cattle. In Book of Abstracts, Proceedings of the 46th Annual Meeting of the European Association for Animal Production (EAAP), Prague, Czech Republic, 4–7 September 1995; van Arendonk, J.A.M., Ed.; Wageningen Pers.: Wageningen, The Netherlands, 1995; p. 27. Available online: https://hal.inrae.fr/hal-02772606 (accessed on 19 January 2022).
- Casas, E.; Kehrli, M.E., Jr. A review of selected genes with known effects on performance and health of cattle. Front. Vet. Sci. 2016, 3, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epishko, O.A.; Pestis, B.K.; Tanana, L.A.; Kuzmina, T.I.; Cheburanova, E.S.; Shevchenko, M.Y.; Petrova, A.P.; Glinskaya, H.A.; Trahimchik, R.V. Definition of recessive mutations of BLAD, CVM and BS in population of cattle of the lactic direction of Republic of Belarus. In Agriculture—Problems and Prospects: Collection of Scientific Papers—Zootechnics; Pestis, V.K., Ed.; Grodno State Agrarian University: Grodno, Belarus, 2017; Volume 37, pp. 44–51. Available online: https://www.elibrary.ru/item.asp?id=32443408 (accessed on 19 January 2022)ISBN 978-985-537-108-4.
- Ivanov, V.A.; Marzanov, N.S.; Eliseeva, L.I.; Tadzhiyev, K.P.; Marzanova, S.N. Genotypes of cattle breeds and quality of milk. Problemy Biologii Productivnykh Zhivotnykh (Probl. Product. Anim. Biol.) 2017, 3, 48–65. Available online: https://www.elibrary.ru/item.asp?id=30058078 (accessed on 19 January 2022).
- Ignatiev, V.M. Screening of the Leukocyte Adhesion Gene (BLAD-Syndrome) in Black-and-White Bovine Animals. Ph.D. Thesis, All-Russia Research Institute for Animal Husbandry, Dubrovitsy, Moscow Oblast, Russia, 1998. Available online: https://elibrary.ru/item.asp?id=19145898 (accessed on 19 January 2022).
- Hagemoser, W.A.; Roth, J.A.; Lofstedt, J.; Fagerland, J.A. Granulocytopathy in a Holstein heifer. J. Am. Vet. Med. Assoc. 1983, 183, 1093–1094. Available online: https://pubmed.ncbi.nlm.nih.gov/6643217/ (accessed on 19 January 2022). [PubMed]
- Kehrli, M.E., Jr.; Schmalstieg, F.C.; Anderson, D.C.; Van der Maaten, M.J.; Hughes, B.J.; Ackermann, M.R.; Wilhelmsen, C.L.; Brown, G.B.; Stevens, M.G.; Whetstone, C.A. Molecular definition of the bovine granulocytopathy syndrome: Identification of deficiency of the Mac-1 (CD11b/CD18) glycoprotein. Am. J. Vet. Res. 1990, 51, 1826–1836. Available online: https://pubmed.ncbi.nlm.nih.gov/1978618/ (accessed on 19 January 2022). [PubMed]
- Kehrli, M.E., Jr.; Shuster, D.E.; Ackermann, M.R. Leukocyte adhesion deficiency among Holstein cattle. Cornell Vet. 1992, 82, 103–109. Available online: https://pubmed.ncbi.nlm.nih.gov/1623723/ (accessed on 19 January 2022).
- Ackermann, M.R.; Kehrli, M.E., Jr.; Morfitt, D.C. Ventral dermatitis and vasculitis in a calf with bovine leukocyte adhesion deficiency. J. Am. Vet. Med. Assoc. 1993, 202, 413–415. Available online: https://pubmed.ncbi.nlm.nih.gov/8440633/ (accessed on 19 January 2022).
- Gilbert, R.O.; Rebhun, W.C.; Kim, C.A.; Kehrli, M.E., Jr.; Shuster, D.E.; Ackermann, M.R. Clinical manifestations of leukocyte adhesion deficiency in cattle: 14 cases (1977–1991). J. Am. Vet. Med. Assoc. 1993, 202, 445–449. Available online: https://pubmed.ncbi.nlm.nih.gov/8095042/ (accessed on 19 January 2022).
- Duncan, R.B., Jr.; Carrig, C.B.; Agerholm, J.S.; Bendixen, C. Complex vertebral malformation in a Holstein calf: Report of a case in the USA. J. Vet. Diagn. Investig. 2001, 13, 333–336. [Google Scholar] [CrossRef] [Green Version]
- VanRaden, P.M.; Olson, K.M.; Null, D.J.; Hutchison, J.L. Harmful recessive effects on fertility detected by absence of homozygous haplotypes. J. Dairy Sci. 2011, 94, 6153–6161. [Google Scholar] [CrossRef] [Green Version]
- Holstein Association USA, Inc. Brattleboro, VT, USA. Available online: https://www.holsteinusa.com/ (accessed on 19 January 2022).
- Select Sires Inc. Plain City, OH, USA. Available online: https://www.selectsires.com/ (accessed on 19 January 2022).
- Anon. VDS-Beschluß zu BLAD. Ostfr. Landvolk. 1992, 107, 28. [Google Scholar]
- Agerholm, J.S.; DeLay, J.; Hicks, B.; Fredholm, M. First confirmed case of the bovine brachyspina syndrome in Canada. Can. Vet. J. 2010, 51, 1349–1350. Available online: https://pubmed.ncbi.nlm.nih.gov/21358926/ (accessed on 19 January 2022).
- Poli, M.A.; Dewey, R.; Semorile, L.; Lozano, M.E.; Albariño, C.G.; Romanowski, V.; Grau, O. PCR screening for carriers of bovine leukocyte adhesion deficiency (BLAD) and uridine monophosphate synthase (DUMPS) in Argentine Holstein cattle. J. Vet. Med. Ser. A 1996, 43, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.A.; Baron, E.E.; Martinez, M.L.; Coutinho, L.L. PCR screening and allele frequency estimation of bovine leukocyte adhesion deficiency in Holstein and Gir cattle in Brazil. Genet. Mol. Biol. 2000, 23, 831–834. [Google Scholar] [CrossRef] [Green Version]
- Schilcher, F.; Hotter, H.; Tammen, I.; Schuh, M. Occurrence of bovine leukocyte adhesion deficiency (BLAD) of Holstein Friesian in Austria. Wien. Tierarztl. Monatsschr. 1995, 82, 207–212. Available online: http://a.xueshu.baidu.com/usercenter/paper/show?paperid=52ca18b04dbff9c033612a462f966bfd (accessed on 19 January 2022).
- Cox, E.; Kuczka, A.; Tammen, I.; Schwenger, B. Leukocyten adhesien deficientie bij rund en hond: Een erfelijke stoornis van het immuunstelsel. Vlaams Diergeneeskd. Tijdschr. 1993, 62, 71–79. Available online: https://library.wur.nl/WebQuery/titel/830081 (accessed on 19 January 2022).
- Georges, M.; Coppieters, W.; Charlier, C.; Agerholm, J.S.; Fredholm, M. A Genetic Marker Test for Brachyspina and Fertility in Cattle. Patent for Invention EP2310528B1. International Patent Publ. No. WO2010/012690, 20 November 2013. Available online: https://patents.google.com/patent/EP2310528B1/en (accessed on 19 January 2022).
- Andrews, A.H.; Fishwick, J.; Waters, R.J. Bovine leukocyte adhesion deficiency (BLAD) in a one year old Holstein Friesian bull—The first report in the United Kingdom. Br. Vet. J. 1996, 152, 347–351. [Google Scholar] [CrossRef]
- Zsolnai, A.; Fésüs, L. Simultaneous analysis of bovine κ-casein and BLAD alleles by multiplex PCR followed by parallel digestion with two restriction enzymes. Anim. Genet. 1996, 27, 207–209. [Google Scholar] [CrossRef]
- Fésüs, L.; Zsolnai, A.; Anton, I.; Bárány, I.; Bozó, S. BLAD genotypes and cow production traits in Hungarian Holsteins. J. Anim. Breed. Genet. 1999, 116, 169–174. [Google Scholar] [CrossRef]
- Agerholm, J.S.; Houe, H.; Jørgensen, C.B.; Basse, A. Bovine leukocyte adhesion deficiency in Danish Holstein-Friesian cattle. II. Patho-anatomical description of affected calves. Acta Vet. Scand. 1993, 34, 237–243. [Google Scholar] [CrossRef]
- Agerholm, J.S.; Basse, A.; Christensen, K. Investigations on the occurrence of hereditary diseases in the Danish cattle population 1989–1991. Acta Vet. Scand. 1993, 34, 245–253. [Google Scholar] [CrossRef]
- Jørgensen, C.B.; Agerholm, J.S.; Pedersen, J.; Thomsen, P.D. Bovine leukocyte adhesion deficiency in Danish Holstein-Friesian cattle. I. PCR screening and allele frequency estimation. Acta Vet. Scand. 1993, 34, 231–236. [Google Scholar] [CrossRef]
- Agerholm, J.S.; Bendixen, C.; Andersen, O.; Arnbjerg, J. Complex vertebral malformation in Holstein calves. J. Vet. Diagn. Investig. 2001, 13, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Agerholm, J.S.; Andersen, O.; Almskou, M.B.; Bendixen, C.; Arnbjerg, J.; Aamand, G.P.; Nielsen, U.S.; Panitz, F.; Petersen, A.H. Evaluation of the inheritance of the complex vertebral malformation syndrome by breeding studies. Acta Vet. Scand. 2004, 45, 133–137. [Google Scholar] [CrossRef]
- Agerholm, J.S.; McEvoy, F.; Arnbjerg, J. Brachyspina syndrome in a Holstein calf. J. Vet. Diagn. Investig. 2006, 18, 418–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agerholm, J.S.; Peperkamp, K. Familial occurrence of Danish and Dutch cases of the bovine brachyspina syndrome. BMC Vet. Res. 2007, 3, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlier, C.; Agerholm, J.S.; Coppieters, W.; Karlskov-Mortensen, P.; Li, W.; de Jong, G.; Fasquelle, C.; Karim, L.; Cirera, S.; Cambisano, N.; et al. A deletion in the bovine FANCI gene compromises fertility by causing fetal death and brachyspina. PLoS ONE 2012, 7, e43085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stöber, M.; Pohlenz, J.; Leibold, W.; Simon, D.; Kuczka, A.; Schwenger, B.; Kuhlmann, E.; Tammen, L.; Piturru, P. Deficienza di adesione dei leucociti nei bovini (BLAD) un difetto ereditario da porre sotto vigilanza. Prax. Vet. 1992, 8, 5–7. [Google Scholar]
- Testoni, S.; Diana, A.; Olzi, E.; Gentile, A. Brachyspina syndrome in two Holstein calves. Vet. J. 2008, 177, 144–146. [Google Scholar] [CrossRef]
- Viana, J.L.; Fernández, A.; Iglesias, A.; Pernas, G.S. Diagnóstico y control de las principales enfermedades genéticas (citrulinemia, DUMPS y BLAD) descritas en ganado Holstein-Frisón. Med. Vet. 1998, 15, 538–544. Available online: https://dialnet.unirioja.es/servlet/articulo?codigo=4410498 (accessed on 19 January 2022).
- Adamov, N.; Mitrov, D.; Esmerov, I.; Dovc, P. Detection of recessive mutations (BLAD and CVM) in Holstein-Friesian cattle population in Republic of Macedonia. Maced. Vet. Rev. 2014, 37, 61–68. [Google Scholar] [CrossRef]
- Müller, K.; Bernadina, W.E.; Kalsbeek, H.C.; Wensing, T.; Elving, L.; Verbeek, B.; Wentink, G.H. BLAD: Bovine leucocyte adhesion deficiency. Tijdschr. Diergeneeskd. 1993, 118, 183–184. Available online: https://pubmed.ncbi.nlm.nih.gov/8096659/ (accessed on 19 January 2022).
- Mirck, M.H.; Von Bannisseht-Wijsmuller, T.; Timmermans-Besselink, W.J.; Van Luijk, J.H.; Buntjer, J.B.; Lenstra, J.A. Optimization of the PCR test for the mutation causing bovine leukocyte adhesion deficiency. Cell. Mol. Biol. 1995, 41, 695–698. Available online: https://pubmed.ncbi.nlm.nih.gov/7580848/ (accessed on 19 January 2022). [PubMed]
- Arrayet, J.L.; Oberbauer, A.M.; Famula, T.R.; Garnett, I.; Oltjen, J.W.; Imhoof, J.; Kehrli, M.E., Jr.; Graham, T.W. Growth of Holstein calves from birth to 90 days: The influence of dietary zinc and BLAD status. J. Anim. Sci. 2002, 80, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Lubieniecki, K.; Grzybowski, G. Diagnostyka molekularna wrodzonego niedoboru leukocytarnych czasteczek adhezyjnych [BLAD] u bydla. Med. Weter. 1997, 53, 214–217. Available online: http://agro.icm.edu.pl/agro/element/bwmeta1.element.agro-article-b700eed0-54f0-4e01-9630-f8a5be251758 (accessed on 19 January 2022).
- Ruść, A.; Kamiński, S. Prevalence of complex vertebral malformation carriers among Polish Holstein-Friesian bulls. J. Appl. Genet. 2007, 48, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Ruść, A.; Kamiński, S. Detection of Brachyspina carriers within Polish Holstein-Friesian bulls. Pol. J. Vet. Sci. 2015, 18, 453–454. [Google Scholar] [CrossRef] [Green Version]
- Marzanov, N.S.; Popov, A.N.; Zinovieva, N.A.; Polezhaeva, V.A.; Ignatiev, V.M.; Brem, G. Screening of the BLAD syndrome gene in Black-and-white root animals. Vestnik Rossiyskoy Akademii Sel’skokhozyaystvennykh Nauk (Her. Russ. Acad. Sci.) 1997, 4, 59–61. Available online: https://elibrary.ru/item.asp?id=21635211 (accessed on 19 January 2022).
- Marzanov, N.S.; Eskin, G.V.; Turbina, I.S.; Devrishov, D.A.; Tokhov, M.K.; Marzanova, S.N. Gene Diagnostics and Distribution of the Immunodeficiency Allele, or BLAD Syndrome, in Black-and-White Cattle; Rosinformagrotech: Moscow, Russia, 2013; Available online: https://elibrary.ru/item.asp?id=21055161 (accessed on 19 January 2022).
- Kalashnikova, L.A.; Dunin, I.M.; Glazko, V.I.; Glazko, G.V.; Ryzhova, N.V.; Golubina, E.P. DNA Technologies in the Evaluation of Farm Animals; All-Russian Research Institute of Animal Breeding: Lesnye Polyany, Moscow Oblast, Russia, 1999; Available online: https://www.elibrary.ru/item.asp?id=25478053 (accessed on 19 January 2022).
- Turbina, I.S.; Fedorova, E.V.; Kertieva, N.M.; Yeskin, G.V.; Turbina, G.S.; Marzanov, N.S. Genealogy and some biological features in BLAD carrier and non-carrier bulls. In Selektsiya, kormleniye, soderzhaniye sel’skokhozyaystvennykh zhivotnykh i tekhnologiya proizvodstva produktov zhivotnovodstva: Trudy VNIIPlem (Breeding, Feeding, Keeping Farm Animals and Technology for the Production of Livestock Products: Proceedings of VNIIPlem); All-Russian Research Institute of Animal Breeding: Lesnye Polyany, Moscow Oblast, Russia, 2004; pp. 3–7. [Google Scholar]
- Kalashnikova, L.A.; Khabibrakhmanova, Y.A.; Prozherin, V.P.; Yaluga, V.L. Genotyping of brachyspina anomaly in Holsteinized cattle of the Kholmogory breed. In Molecular Diagnostics 2021—Section 21: Technologies for the Detection of Infectious and Hereditary Animal Diseases, Proceedings of the 10th Anniversary International Scientific and Practical Conference, Moscow, Russia, 9–11 November 2021; Center for Strategic Planning and Management of Biomedical Health Risks: Moscow, Russia, 2021; Volume 2, pp. 186–188. [Google Scholar]
- Vătăşescu-Balcan, R.A.; Manea, M.A.; Georgescu, S.E.; Dinischiotu, A.; Tesio, C.D.; Costache, M. Evidence of single point mutation inducing BLAD disease in Romanian Holstein-derived cattle breed. Biotechnol. Anim. Husb. 2007, 23, 375–381. [Google Scholar] [CrossRef]
- Grobet, L.; Charlier, C.; Hanset, R. Diagnosis of bovine leucocyte adhesion deficiency (BLAD) at the DNA level. Ann. Med. Vet. 1993, 137, 27–31. Available online: https://agris.fao.org/agris-search/search.do?recordID=BE9300687 (accessed on 19 January 2022).
- Eggen, A.; Duchesne, A.; Laurent, P.; Grohs, C.; Denis, C.; Gautier, M.; Boichard, D.; Ducos, A. Controlling genetic disorders in the French dairy cattle population. In Proceedings of the 29th International Conference on Animal Genetics, Tokyo, Japan, 11–16 September 2004; International Society for Animal Genetics (ISAG), INT: Tokyo, Japan, 2004; p. 35. Available online: https://hal.inrae.fr/hal-02762175/file/ISAG_Proceedings_2004_1.pdf (accessed on 19 January 2022).
- Valour, D.; Michot, P.; Eozenou, C.; Lefebvre, R.; Bonnet, A.; Capitan, A.; Uzbekova, S.; Sellem, E.; Ponsart, C.; Schibler, L. Dairy cattle reproduction is a tightly regulated genetic process: Highlights on genes, pathways, and biological processes. Anim. Front. 2015, 5, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Anon. Erbfehler—Was ist Blad? Hann. Land. Forstwirtsch. Ztg. 1991, 11, 91. [Google Scholar]
- Kuczka, A.; Schwenger, B. BLAD im Griff [The right feeling for BLAD [bovine leukocyte adhesion deficiency]]. Tierzüchter 1993, 45, 32–35. Available online: https://agris.fao.org/agris-search/search.do?recordID=DE19930058362 (accessed on 19 January 2022).
- Lienau, A.; Stöber, M.; Kehrli, M.E.; Tammen, I.; Schwenger, B.; Kuczka, A.; Pohlenz, J. Bovine Leukozyten-Adhäsions-Defizienz: Klinisches Bild und Differential-Diagnostik (Bovine leukocyte adhesion deficiency: Clinical picture and differential diagnosis). Dtsch. Tierarztl. Wochenschr. 1994, 101, 405–406. Available online: https://pubmed.ncbi.nlm.nih.gov/7851303/ (accessed on 19 January 2022).
- Tammen, I. Weiterentwicklung des DNA-Tests auf BLAD (Bovine Leukozyten-Adhäsions-Defizienz) für den Einsatz in Rinderzucht und Klinischer Diagnostik. Ph.D. Thesis, Tierärztliche Hochschule Hannover, Hannover, Germany, 1994. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201300291400 (accessed on 19 January 2022).
- Duesmann, K.; Schmidt, W.; Görlach, A. Die PCR als Methode zur Gendiagnostik im Routinebetrieb einer Besamungsstation am Beispiel der Bovinen Leukozyten-Adhäsions-Defizienz (BLAD). Reprod. Domest. Anim. 1994, 29, 193. [Google Scholar]
- Engelhardt, I. Inzucht, Bedeutende Ahnen und Wahrscheinlichkeit für BLAD-Merkmalsträger in der Deutschen Schwarzbuntzucht. Ph.D. Thesis, Tierärztliche Hochschule Hannover, Hannover, Germany, 1996. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201300016803 (accessed on 19 January 2022).
- Distl, O.T. The use of molecular genetics in eliminating of inherited anomalies in cattle. Arch. Anim. Breed. 2005, 48, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Buck, B.C.; Ulrich, R.; Wöhlke, A.; Kuiper, H.; Baumgaertner, W.; Distl, O. Missbildungen an der Wirbelsaule mit multiplen Organanomalien bei einem Kalb der Rasse Deutsche Holsteins (Vertebral and multiple organ malformations in a black and white German Holstein calf). Berl. Munch. Tierarztl. Wochenschr. 2010, 123, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Segelke, D.; Täubert, H.; Reinhardt, F.; Thaller, G. Considering genetic characteristics in German Holstein breeding programs. J. Dairy Sci. 2016, 99, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Glazko, V.I.; Lavrovsky, V.V.; Filenko, A.N.; Mariutsa, A.E. Intrabreed genetic differentiation and the presence of the BLAD mutation in Holstein . Sel’skokhozyaistvennaya Biol. (Agr. Biol.) 2000, 4, 45–47. Available online: https://scholar.google.com/scholar?cluster=6636562904833334721&hl=en&oi=scholarr (accessed on 19 January 2022).
- Mariutsa, A.E. Population-Genetic Mechanisms of Adaptation and Spread of Semi-Lethal Recessive Mutations on the Example of BLAD in Cattle. Ph.D. Thesis, Institute of Agroecology and Environmental Management UAAS, Kyiv, Ukraine, 2005. Available online: http://www.disslib.org/populjatsyonno-henetycheskye-mekhanyzmy-adaptatsyy-y-rasprostranenye-poluletalnykh.html (accessed on 19 January 2022).
- Birukova, O.D. The Population-Genetic Monitoring of Gene Pool Forming of the Ukrainian Black-and-White Dairy Breed. Ph.D. Thesis, Institute of Animal Breeding and Genetics UAAS, Chubinske, Kyiv Region, Ukraine, 2005. Available online: http://www.irbis-nbuv.gov.ua/cgi-bin/irbis_nbuv/cgiirbis_64.exe?C21COM=2&I21DBN=ARD&P21DBN=ARD&Z21ID=&Image_file_name=DOC/2005/05bodrmp.zip&IMAGE_FILE_DOWNLOAD=1 (accessed on 19 January 2022).
- Meylan, M.; Abegg, R.; Sager, H.; Jungi, T.W.; Martig, J. Fallvorstellung: Bovine Leukozyten-Adhäsions-Defizienz (BLAD) in der Schweiz (Case presentation: Bovine leukocyte adhesion deficiency (BLAD) in Switzerland). Schweiz. Arch. Tierheilkd. 1997, 139, 277–281. Available online: https://pubmed.ncbi.nlm.nih.gov/9411734/ (accessed on 19 January 2022). [PubMed]
- Citek, J.; Rehout, V.; Hajkova, J.; Pavkova, J. Monitoring of the genetic health of cattle in the Czech Republic. Vet. Med. 2006, 51, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Han, G.; Zhang, S.; Li, N.; Sun, F. Detection of bovine leukocyte adhesion deficiency (BLAD) and haplotype analysis. Acta Vet. Zootech. Sin. 2008, 39, 1285–1288. Available online: http://www.xmsyxb.com/CN/Y2008/V39/I9/1285 (accessed on 19 January 2022).
- Chu, Q.; Sun, D.; Yu, Y.; Zhang, Y.; Zhang, Y. Identification of complex vertebral malformation carriers in Chinese Holstein. J. Vet. Diagn. Investig. 2008, 20, 228–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Fan, X.; Sun, D.; Wang, Y.; Yu, Y.; Xie, Y.; Zhang, S.; Zhang, Y. A novel method for rapid and reliable detection of complex vertebral malformation and bovine leukocyte adhesion deficiency in Holstein cattle. J. Anim. Sci. Biotechnol. 2012, 3, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, L.; Li, Y.; Zhang, Y.; Sun, D.; Liu, L.; Zhang, Y.; Zhang, S. Identification of brachyspina syndrome carriers in Chinese Holstein cattle. J. Vet. Diagn. Investig. 2013, 25, 508–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhai, L.; Fang, L.; Zhang, S.; Liu, L.; Zhu, Y.; Xue, J.; Xiaoqing, L.; Qiao, L.; Sun, D. A novel multiplex polymerase chain reaction method for the identification of brachyspina syndrome carriers in Chinese Holstein cattle. J. Vet. Sci. Med. Diagn. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Miyagawa, K.; Abe, S.; Kurosawa, T.; Sonoda, M.; Nakade, T.; Nagahata, H.; Noda, H.; Chihaya, Y.; Isogai, E. Bovine granulocytopathy syndrome of Holstein-Friesian calves and heifers. Nihon Juigaku Zasshi 1987, 49, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Tajima, M.; Irie, M.; Kirisawa, R.; Hagiwara, K.; Kurosawa, T.; Takahashi, K. The detection of a mutation of CD18 gene in bovine leukocyte adhesion deficiency (BLAD). J. Vet. Med. Sci. 1993, 55, 145–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahata, H.; Miura, T.; Tagaki, K.; Ohtake, M.; Noda, H.; Yasuda, T.; Nioka, K. Prevalence and allele frequency estimation of bovine leukocyte adhesion deficiency (BLAD) in Holstein-Friesian cattle in Japan. J. Vet. Med. Sci. 1997, 59, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahata, H. Bovine leukocyte adhesion deficiency (BLAD): A review. J. Vet. Med. Sci. 2004, 66, 1475–1482. [Google Scholar] [CrossRef] [Green Version]
- Healy, P.J. Bovine leukocyte adhesion deficiency (BLAD)—Another genetic defect of Holstein/Friesians. Aust. Vet. J. 1992, 69, 190. [Google Scholar] [CrossRef]
- Norouzy, A.; Nassiry, M.R.; Eftekhari Shahrody, F.; Javadmanesh, A.; Mohammad Abadi, M.R.; Sulimova, G.E. Identification of bovine leukocyte adhesion deficiency (BLAD) carriers in Holstein and Brown Swiss AI bulls in Iran. Russ. J. Genet. 2005, 41, 1409–1413. [Google Scholar] [CrossRef]
- Rezaee, A.R.; Nassiry, M.R.; Valizadeh, R.; Tahmoorespour, M.; Javadmanesh, A.; Zarei, A.; Janati, H. Study of complex vertebral malformation disorder in Iranian Holstein bulls. World J. Zool. 2008, 3, 36–39. Available online: https://profdoc.um.ac.ir/paper-abstract-1014702.html (accessed on 19 January 2022).
- Hemati, B.; Saberi, J.; Noshary, A.R. The study of complex vertebral malformation genetic defect in a population of Sistani cows. J. Plant Anim. Environ. Sci. 2014, 4, 485–487. Available online: https://www.cabdirect.org/cabdirect/abstract/20143288422 (accessed on 19 January 2022).
- Öner, Y.; Keskin, A.; Elmaci, C. Identification of BLAD, DUMPS, citrullinaemia and factor XI deficiency in Holstein cattle in Turkey. Asian J. Anim. Vet. Adv. 2010, 5, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.K.; Singh, K.M.; Soni, K.J.; Chauhan, J.B.; Sambasiva Rao, K.R. Low incidence of bovine leukocyte adhesion deficiency (BLAD) carriers in Indian cattle and buffalo breeds. J. Appl. Genet. 2007, 48, 153–155. [Google Scholar] [CrossRef]
- Gholap, P.N.; Kale, D.S.; Sirothia, A.R. Genetic diseases in cattle: A review. Res. J. Anim. Vet. Fish. Sci. 2014, 2, 24–33. Available online: http://www.isca.in/AVFS/Archive/v2/i2/5.ISCA-RJAVFS-2014-005.php (accessed on 19 January 2022).
- Nasreen, F.; Altaf Malik, N.; Naeem Riaz, M.; Anver Qureshi, J. Detection and screening of bovine leukocyte adhesion deficiency in Pakistan using molecular methods. Hereditas 2009, 146, 74–78. [Google Scholar] [CrossRef]
- Stadler, P.; Van Amstel, S.R.; Van Rensburg, I.B.; Williams, M.C. Verdagte oorerflike granulositopatie in vier Holstein frieskalwers (Suspected inherited granulocytopathy in four Holstein Friesian calves). J. S. Afr. Vet. Assoc. 1993, 64, 172–177. Available online: https://pubmed.ncbi.nlm.nih.gov/8176699/ (accessed on 19 January 2022).
- FAO. Gateway to Dairy Production and Products. Available online: https://www.fao.org/dairy-production-products/en/ (accessed on 19 January 2022).
- Kosyachenko, N.M.; Abramova, M.V.; Ilyina, A.V.; Zyryanova, S.V.; Konovalov, A.V.; Kosourova, T.N. Holstein Breed in the Creation of Improved Genotypes and Intrabreed Types of Cattle; Kantsler: Yaroslavl, Russia, 2020; ISBN 978-5-907417-06-9. Available online: https://search.rsl.ru/ru/search#q=ISBN%20978-5-907417-06-9 (accessed on 19 January 2022).
- Cole, J.B.; Null, D.J.; VanRaden, P.M. Phenotypic and genetic effects of recessive haplotypes on yield, longevity, and fertility. J. Dairy Sci. 2016, 99, 7274–7288. [Google Scholar] [CrossRef] [Green Version]
- Marzanov, N.S.; Turbina, I.S.; Eskin, G.V.; Turbina, G.S.; Ignat’ev, V.M.; Popov, A.N.; Harlizius, B. Screening of the gene of the leukocytal adhesion deficiency in Black-and-white Holsteinized cattle. Sel’skokhozyaistvennaya Biol. (Agr. Biol.) 2003, 38, 23–30. Available online: https://www.elibrary.ru/item.asp?id=18100805 (accessed on 19 January 2022).
- Grant, V. The Evolutionary Process: A Critical Study of Evolutionary Theory; Columbia University Press: New York, NY, USA, 1991; ISBN 0-231-07324-0. Available online: https://www.google.com/books/edition/The_Evolutionary_Process/qAt_QgAACAAJ?hl=en (accessed on 19 January 2022).
- Kaminski, S.; Cieslinska, A.; Kostyr, E. Polymorphism of bovine beta-casein and its potential effect on human health. J. Appl. Genet. 2007, 48, 189–198. [Google Scholar] [CrossRef]
- Woodford, K.B. Devil in the Milk: Illness, Health and Politics of A1 and A2 Milk; Chelsea Green Publishing: White River Junction, VT, USA, 2009; Available online: https://www.google.com/books/edition/_/QqD6btySu7AC?hl=en (accessed on 19 January 2022)ISBN 978-1-60358-102-8.
- Marzanov, N.S.; Devrishov, D.A.; Abylkasymov, D.A.; Marzanova, S.N.; Konovalova, N.V.; Libet, I.S. Characteristics of Russian dairy cattle by the occurrence of genotypes and alleles in the beta casein locus. Veterenariya Zootekhniya i Biotehnologiya (Vet. Med. Zootech. Biotechnol.), 2020; 1, 47–52. [Google Scholar] [CrossRef]
- Marzanov, N.S.; Abylkasymov, D.A.; Devrishov, D.A.; Marzanova, S.N.; Libet, I.S. The characteristic of allelotype in cows of Black and multicolored breed of β- and k-casein locus and qualitative indicators of milk. In Aktual’nyye Voprosy Molochnoy Promyshlennosti, Mezhotraslevyye Tekhnologii i Sistemy Upravleniya Kachestvom (Actual Issues of the Dairy Industry, Intersectoral Technologies and Quality Management Systems); All-Russian Dairy Research Institute: Moscow, Russia, 2020; pp. 368–376. [Google Scholar] [CrossRef]
- Marzanova, S.N.; Alekseev, Y.I.; Konovalova, N.V.; Turbina, I.S.; Devrishov, D.A.; Sochivko, D.G.; Marzanov, N.S. Developing a method of diagnostics of complex vertebral malformation (CVM) by Real-Time PCR method in Black-and-white cattle. Problemy Biologii Produktivnyh Životnyh (Probl. Product. Anim. Biol.) 2011, 79–82. Available online: https://elibrary.ru/item.asp?id=17644709 (accessed on 19 January 2022).
- Marzanov, N.S.; Devrishov, D.A.; Marzanova, S.N.; Getokov, О.О.; Abylkasymov, D.A.; Libet, I.S. DNA diagnostics of populations of the Blackand-white breed by the beta-casein locus. Veterenariya Zootekhniya i Biotehnologiya (Vet. Med. Zootech. Biotechnol.) 2021, 3, 78–84. [Google Scholar] [CrossRef]
- Wiggans, G.R.; Cole, J.B.; Hubbard, S.M.; Sonstegard, T.S. Genomic Selection in Dairy Cattle: The USDA Experience. Annu. Rev. Anim. Biosci. 2017, 5, 309–327. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.; Kearney, J.F.; Cromie, A.R.; Hely, F.S.; Amer, P.R. Genetic benefits of genomic selection breeding programmes considering foreign sire contributions. Genet. Sel. Evol. 2019, 51, 40. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Reinoso, M.A.; Aponte, P.M.; Garcia-Herreros, M. Genomic Analysis, Progress and Future Perspectives in Dairy Cattle Selection: A Review. Animals 2021, 11, 599. [Google Scholar] [CrossRef]
- Bouquet, A.; Juga, J. Integrating genomic selection into dairy cattle breeding programmes: A review. Animal 2013, 7, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Brito, L.F.; Bedere, N.; Douhard, F.; Oliveira, H.R.; Arnal, M.; Peñagaricano, F.; Schinckel, A.P.; Baes, C.F.; Miglior, F. Review: Genetic selection of high-yielding dairy cattle toward sustainable farming systems in a rapidly changing world. Animal 2021, 15 (Suppl. S1), 100292. [Google Scholar] [CrossRef]
- Smaragdov, M.G. Genomic selection of milk cattle. The practical application over five years. Russ. J. Genet. 2013, 49, 1089–1097. [Google Scholar] [CrossRef]
- Plemyashov, К.V.; Smaragdov, M.G.; Romanov, M.N. Molecular Genetic Polymorphism in Animal Populations and Its Application in Intensive Breeding of Dairy Cattle—A Review. In Materials of the 3rd International Scientific and Practical Conference on Molecular Genetic Technologies for Analysis of Gene Expression Related to Animal Productivity and Disease Resistance, Moscow, Russia, 30 September 2021; Pozyabin, S.V., Kochish, I.I., Romanov, M.N., Eds.; Sel’skokhozyaistvennye tekhnologii: Moscow, Russia, 2021; pp. 368–378. Available online: https://elibrary.ru/item.asp?id=46668865 (accessed on 19 January 2022).
- Plemyashov, К.V.; Smaragdov, M.G.; Romanov, M.N. Genomic Assessment of Breeding Bulls. In Materials of the 3rd International Scientific and Practical Conference on Molecular Genetic Technologies for Analysis of Gene Expression Related to Animal Productivity and Disease Resistance, Moscow, Russia, 30 September 2021; Pozyabin, S.V., Kochish, I.I., Romanov, M.N., Eds.; Sel’skokhozyaistvennye tekhnologii: Moscow, Russia, 2021; pp. 363–367. Available online: https://elibrary.ru/item.asp?id=46668864 (accessed on 19 January 2022).
- Sharko, F.S.; Khatib, A.; Prokhortchouk, E.B. Genomic estimated breeding value of milk performance and fertility traits in the Russian Black-and-white cattle population. Acta Nat. 2022, 14, 109–122. [Google Scholar] [CrossRef]
- Zinovieva, N.A.; Dotsev, A.V.; Sermyagin, A.A.; Deniskova, T.E.; Abdelmanova, A.S.; Kharzinova, V.R.; Sölkner, J.; Reyer, H.; Wimmers, K.; Brem, G. Selection signatures in two oldest Russian native cattle breeds revealed using high-density single nucleotide polymorphism analysis. PLoS ONE 2020, 15, e0242200. [Google Scholar] [CrossRef]
- Krivonogova, A.S.; Bruter, A.V.; Makutina, V.A.; Okulova, Y.D.; Ilchuk, L.A.; Kubekina, M.V.; Khamatova, A.Y.; Egorova, T.V.; Mymrin, V.S.; Silaeva, Y.Y.; et al. AAV infection of bovine embryos: Novel, simple and effective tool for genome editing. Theriogenology 2022, 193, 77–86. [Google Scholar] [CrossRef]
- Cziszter, L.-T.; Ilie, D.-E.; Neamt, R.-I.; Neciu, F.-C.; Saplacan, S.-I.; Gavojdian, D. Comparative study on production, reproduction and functional traits between Fleckvieh and Braunvieh cattle. Asian-Australas. J. Anim. Sci. 2017, 30, 666–671. [Google Scholar] [CrossRef]
- Ernst, L.K.; Zhigachev, A.I. Monitoring of Animal Genetic Diseases in the System of Large-Scale Breeding; Russian Agricultural Academy Publishing House: Moscow, Russia, 2006; ISBN 5-85941-250-9. Available online: https://search.rsl.ru/ru/search#q=ISBN%205-85941-250-9 (accessed on 19 January 2022).
- Marzanov, N.S.; Amerkhanov, H.; Yeskin, G.; Turbina, G.; Fedorova, E.; Turbina, I.; Samorukov, Y.; Kiyko, E.; Popov, N.; Shukurova, E.; et al. Genogeography of BLAD in populations of Black-and-white cattle in Russia and abroad. Molochnoye i Myasnoye Skotovodstvo (Dairy Beef Cattle Breed.) 2008, 4, 2–5. Available online: https://www.elibrary.ru/item.asp?id=11149171 (accessed on 19 January 2022).
- Ussenbekov, E.S.; Terletskiy, V.P. On the absence of genetic defects BLAD, CVM, DUMPS and BC in stud bulls of the local Alatau cattle breed. Veterinariya (Vet. Med.) 2016, 6, 49–51. Available online: https://www.elibrary.ru/item.asp?id=26556053 (accessed on 19 January 2022).
- Terletskiy, V.P.; Buralkhiyev, B.A.; Ussenbekov, Y.S.; Yelubayeva, M.; Tyshchenko, V.I.; Beyshova, I.S. Screening for mutations that determine the development of hereditary diseases in breeding cattle. Aktual’nyye Voprosy Veterinarnoy Biologii (Actual Quest. Vet. Biol.) 2016, 3, 3–6. Available online: https://www.elibrary.ru/item.asp?id=26644420 (accessed on 19 January 2022).
- Marron, B.M.; Robinson, J.L.; Gentry, P.A.; Beever, J.E. Identification of a mutation associated with factor XI deficiency in Holstein cattle. Anim. Genet. 2004, 35, 454–456. [Google Scholar] [CrossRef]
- Kuznetsov, V.M. Statistical analysis of pedigrees. Zootekhniya (Zootechnics). 1998, 2, pp. 5–8. Available online: https://vm-kuznetsov.ru/files/book/Pap1998_2_pedig.pdf (accessed on 19 January 2022).
- Zhigachev, A.I.; Suller, I.L. Hereditary anomalies and their control in cattle. Praktik (Practitioner), 2002; 3–4, 46–53. [Google Scholar]
- Zhigachev, A.I. Estimation of breeding male for invisible genetic defects. Zootekhniya (Zootechnics) 2001, 2, 10–12. Available online: https://www.elibrary.ru/item.asp?id=9124606 (accessed on 19 January 2022).



{kind=link}
{kind=link}
| Region | Country | References |
|---|---|---|
| North America | us USA | [24,28,29,30,31,32,33,34,35,36] * |
| ca Canada | [37,38] * | |
| Latin America and Caribbean | ar Argentina | [39] |
| br Brazil | [40] | |
| Europe | at Austria | [41] |
| be Belgium | [37,42,43] * | |
| gb United Kingdom | [44] | |
| hu Hungary | [45,46] | |
| dk Denmark | [47,48,49,50,51,52,53,54] * | |
| it Italy | [55,56] * | |
| es Spain | [57] | |
| mk North Macedonia | [58] | |
| nl The Netherlands | [53,59,60,61] * | |
| pl Poland | [62,63,64] * | |
| ru Russia | [19,20,27,65,66,67,68,69] * | |
| ro Romania | [70] | |
| fr France | [23,71,72,73] * | |
| de Germany | [74,75,76,77,78,79,80,81,82] * | |
| ua Ukraine | [83,84,85] | |
| ch Switzerland | [86] | |
| cz Czech Republic | [87] | |
| Southeast Asia and the Pacific Basin | cn China | [88,89,90,91,92] * |
| tw Taiwan | [22] * | |
| jp Japan | [93,94,95,96] | |
| nz New Zealand au Australia | [97] | |
| Middle East | ir Iran | [98,99,100] |
| tr Turkey | [101] | |
| South Asia | in India | [102,103] |
| pk Pakistan | [104] | |
| Africa | za South Africa | [105] |
| Breed | n | Gt | Genotype Frequency | Allele Frequency | χ2 | df | p | |||
|---|---|---|---|---|---|---|---|---|---|---|
| CSN2A1/A1 | CSN2A1/A2 | CSN2A2/A2 | CSN2A1 | CSN2A2 | ||||||
| Russian Black Pied | 50 | O | 15 | 26 | 9 | 0.56 | 0.44 | 0.145 | 1 | >0.05 |
| E | 15.68 | 24.64 | 9.68 | |||||||
| Holstein Friesian | 30 | O | 4 | 17 | 9 | 0.42 | 0.58 | 0.82 | 1 | >0.05 |
| E | 5.21 | 14.58 | 10.21 | |||||||
| Yaroslavl | 30 | O | 9 | 13 | 8 | 0.52 | 0.48 | 0.53 | 1 | >0.05 |
| E | 8.0093 | 14.9833 | 7.0074 | |||||||
| Bestuzhev | 67 | O | 35 | 24 | 8 | 0.67 | 0.33 | 0.98 | 1 | >0.05 |
| E | 39 | 21 | 7 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marzanova, S.N.; Devrishov, D.A.; Turbina, I.S.; Marzanov, N.S.; Griffin, D.K.; Romanov, M.N. Genetic Load of Mutations Causing Inherited Diseases and Its Classification in Dairy Cattle Bred in the Russian Federation. Agriculture 2023, 13, 299. https://doi.org/10.3390/agriculture13020299
Marzanova SN, Devrishov DA, Turbina IS, Marzanov NS, Griffin DK, Romanov MN. Genetic Load of Mutations Causing Inherited Diseases and Its Classification in Dairy Cattle Bred in the Russian Federation. Agriculture. 2023; 13(2):299. https://doi.org/10.3390/agriculture13020299
Chicago/Turabian StyleMarzanova, Saida N., Davud A. Devrishov, Irina S. Turbina, Nurbiy S. Marzanov, Darren K. Griffin, and Michael N. Romanov. 2023. "Genetic Load of Mutations Causing Inherited Diseases and Its Classification in Dairy Cattle Bred in the Russian Federation" Agriculture 13, no. 2: 299. https://doi.org/10.3390/agriculture13020299