1. Introduction
Due to their relatively low toxicity to humans and high insecticidal efficiency, pyrethroids are among the most widely used insecticides [
1]. Beta-cypermethrin is one of the most commonly used synthetic pyrethroids to protect crops by reducing pest infestations, thereby minimizing yield losses. However, with the extensive use of pyrethroids, many pests have developed varying degrees of resistance to these insecticides [
2,
3]: for example,
Spodoptera litura [
4],
Musca domestica [
5], the maize fall armyworm [
6] and the picture-winged flies [
7]. Pyrethroids have also been used for a long time to control aphids such as
Aphis glycines [
8],
Aphis gossypii [
9] and wheat aphids [
10]. However, Gong et al. monitored the resistance of wheat aphids to pyrethroids in 15 provinces of China in the period from 2018 to 2019, finding numerous wheat aphids’ moderate or higher resistance to pyrethroids; the most notable among these aphids include
Sitobion avenae and
Rhopalosiphum padi [
10]. In light of such previous studies, novel research on the resistance mechanism of beta-cypermethrin to pest is highly significant when such a study ensures the rational use of pyrethroids as well as suggesting ways of delaying pests’ early resistance to pyrethroids [
11].
Resistance to insecticides is a common adaptive strategy. The increased expression of metabolic resistance-related protein family genes and the decreased penetration rate of insect epidermis are two very important resistance mechanisms [
12].
Penetration resistance refers to modifications in the cuticle that eventually slows down the penetration of insecticide molecules into insects’ body. So far, two mechanisms of penetration resistance have been described: cuticle thickening and altering cuticle composition. Cuticular modifications occur due to the overexpression of diversified genes or exoskeleton proteins such as cuticular proteins, enzymes-catalyzed enzymatic reactions (
CYP4G16 and laccase 2) or ABC transporters that promote cuticular translocation [
13]. Many experiments have demonstrated that epidermal penetration is integral to insects’ resistance to pyrethroids. For example,
Blattella Germanica, a beta-cypermethrin-resistant insect, has lower epidermal penetration than that of sensitive strains [
14]. According to additional research, cuticular penetration is a significant factor in the development of fenvalerate resistance in common house mosquitoes such as
Culex pipiens pallens [
15].
Insects exhibit pyrethroid-resistance as a result of an increase in the expression levels of multiple metabolic detoxification enzymes. Pyrethroid insecticides are mainly metabolized by the oxidation of methyl, methylene, alkyl and aryl substituents, and the resulting metabolites are hydrolyzed by the corresponding alcohols, phenols, acids and glycine, sulfate, glucuronic acid or glucoside and ester bonds, which can be excreted by insects [
11]. Oxidation requires phase I detoxifying enzymes of Cytochromes P450 (P450s) [
16], phase II detoxifying enzymes of UDP–glucose–transferase (UGTs) [
17] and glutathione–s–transferase (GSTs) [
18], and ATP binding cassette transporters (ABCs) [
19], while ester bond breaking requires esterases such as carboxyl/cholinesterase (CCEs) [
20]. Among them, P450s are the superfamily of detoxifying enzymes, and they play a crucial role in the detoxification of many exogenous substances found in insects [
21]. Many related studies have linked high expression levels of the cytochrome P450 gene with pyrethroid resistance in the aphid. For example, P450 plays an important role in pyrethroid resistance in
Aphis gossypii,
Myzus persicae and
Aphis glycines [
22,
23,
24]. Further studies on the resistance of UGT-related genes to insects are also in progress. For instance, the major phase II detoxification enzymes—Uridine diphosphate (UDP) and -glycosyltransferases (UGTs)—have been implicated in the glycosylation of lipophilic endobiotics and xenobiotics and thus, potentially lead to the evolution of insecticide resistance [
25]. Moreover, some UGTs have also been found to be associated with insecticide resistance [
26,
27,
28].
Rhopalosiphum padi (Hemiptera: Aphididae) (
R. padi) is among the dominant wheat aphid populations found in wheat [
29]. At present, the use of pyrethroids insecticides is the major technique to control the growth and spread of
R. padi [
30]. Thus, it is very necessary to study the resistance of aphids to pyrethroids. We assumed that insect resistance to insecticides might be the result of the co-regulation of multiple genes, and a significant number of genes can be searched more comprehensively through the transcriptome. Therefore, we used multiple transcriptome comparisons to compare the differentially expressed genes in multiple dimensions to expose key insecticide-related resistance genes more easily. For example, Feng et al. used short-term induction of agents to select resistance-related genes in their insecticide resistance study on
Tetranychus cinnabarinus [
31], and Wang et al. also used short-term induction of agents to obtain one of the transcripts in their insecticide resistance study on
R. padi [
32].
In this study, a highly beta-cypermethrin-resistant R. padi population was found in the field while high-throughput RNA-seq was used to determine the transcriptome profile of beta-cypermethrin-resistant (Beta-R) and beta-cypermethrin-sensitive (SS) R. padi. We treated the two R. padi populations (SS and Beta-R) with a spray solution of lethal beta-cypermethrin concentration (i.e., LC50) to produce different transcripts; then, we compared the transcripts of beta-cypermethrin-sensitive and beta-cypermethrin-resistant strains of R. padi, and we also compared the transcripts of beta-cypermethrin-susceptible and -resistant strains of R. padi before and after their exposure to beta-cypermethrin. Through this process, effective resistant genes were screened out. In consequence, the results of the study are extremely useful for understanding the beta-cypermethrin resistance mechanism of R. padi and developing more effective management strategies.
2. Materials and Methods
2.1. Aphid Strain
For the current study, the R. padi population used in the experiments was collected from Shizuishan in Ningxia (NXS) in 2018 (N39°05′57″, E106°44′51″), and the sensitive strain (SS) and beta-cypermethrin resistant strain (Beta-R) were obtained from different single-female populations. After rearing the population for a specific period of time, the lethal beta-cypermethrin concentration (i.e LC50) of the single female population to beta-cypermethrin (95% purity, Beijing Green Agricultural Science and Technology Group Co., Ltd., Beijing, China) was evaluated; this measurement showed the lowest LC50 as the sensitive strain (SS), and the sensitive strain was not exposed to any chemical agent during the rearing period. The population with the highest LC50 was selected as the primary resistant strain, and selection for the beta-cypermethrin resistance strain was performed via spraying the aqueous solution of beta-cypermethrin once a week with concentrations their current lethal concertation (i.e., LC50) for the pyrethroid insecticide. Meanwhile, the lethal concertation (i.e., LC50) of R. padi resistance strain to beta-cypermethrin was evaluated every third generation. The high resistance strain (Beta-R) was established after continuous culture for multiple generations.
2.2. Bioassay
The LC
50 of the pyrethroids (beta-cypermethrin) was tested with leaf-dip bioassays [
33]. Beta-cypermethrin was dissolved in acetone and used to prepare five serial concentrations in water containing 0.1% Tween 80, respectively. Wheat leaves with about 60 apterous adult
R. padi were dipped into insecticide solutions for 3–5 s; the residual droplets of the solution on the leaves were absorbed by dry filter paper and subsequently placed in a 90 mm diameter plastic Petri dish. The control was treated with water containing 0.1% Tween 80 and 0.1% acetone alone. Three replicates of approximately 60 individuals were used for each insecticide concentration, and 5 serial concentrations were used for each insecticide and a control. The mortality was checked after 24 h. During and after treatment, the aphids were maintained at 22–24 °C with a photoperiod of 16:8 (L:D) h and a relative humidity of 60 ± 10%.
2.3. RNA Isolation
We diluted beta-cypermethrin to the LC50 of the sensitive and resistant strains. The sensitive strain (SS) was divided into two groups; one group was treated with SS without any external treatment, while the other group was treated for short-term spray treatment with SS strain LC50 (3.22 mg/L) and it was named SS-T. The resistant strain Beta-R was also divided into two groups. One group was treated with Beta-R alone, while the other received a short-term spray treatment with Beta-R strain LC50 (14,774.89 mg/L) and was designated Beta-R-T. Later, the samples (10 wingless adults from each treatment along with four replicates per treatment) were collected after 24 h. Altogether, 16 samples were collected for RNA-seq.
Total RNA was isolated by using RNA Easy Fast Tissue/Cell Kit (Tiangen, Beijing, China). The RNA concentration and purity were measured using a Nano Photometer N50 Touch (IMPLEN, München, Germany). The integrity of RNA was confirmed using Agarose gel electrophoresis.
2.4. Library Preparation and Sequencing
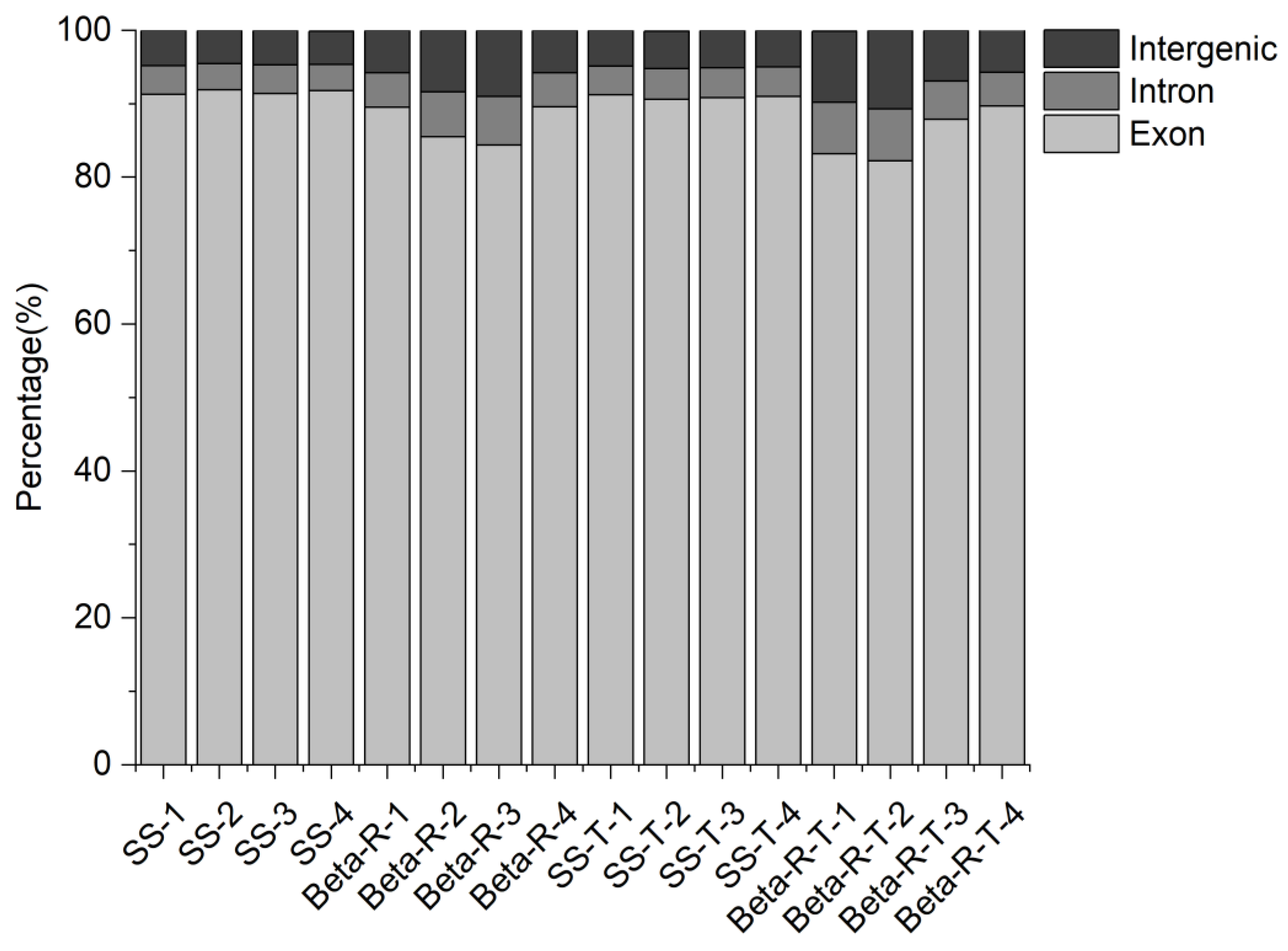
Illumina-based RNA-seq was performed on 4 treatments of
R. padi and its 4 replicas for each treatment. Based on synthesis and sequencing technology, cDNA libraries were sequenced, and numerous reads were produced. These ‘raw reads’ are usually provided in FASTQ (FQ) format. To obtain clean reads, the adapter, ploy-N, and low-quality sequences were removed from raw reads. Q score, GC content, and sequence duplication level were calculated to obtain clean, high-quality reads, which were then utilized for all subsequent analyses. The clean reads were mapped to the
R. padi genome (GenBank no. GCA_020882245.1), and the expression levels of the genes were calculated by using fragments per kilobase per million reads (FPKM) values [
34].
2.5. Functional Annotation
KOBAS 2.0 software, and topGO R packages were used to compare selected transcript sequences against two databases: Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG).
2.6. Differentially Expressed Genes (DEGs) Analysis and Annotation
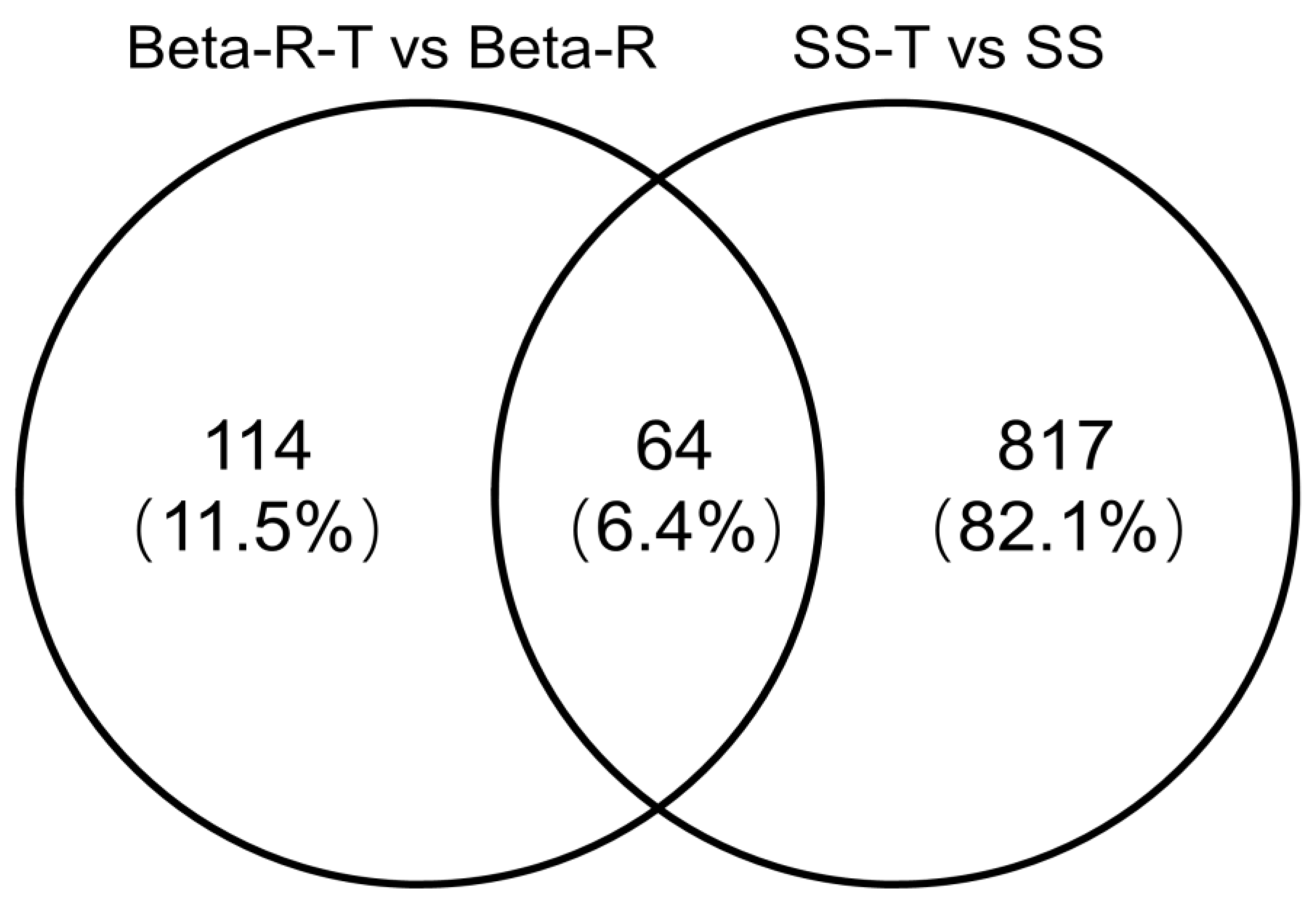
We evaluated four transcription profiles: “Beta-R-T compared with Beta-R,” “SS-T compared with SS,” “Beta-R compared with SS-T,” and “Beta-R compared with SS” in order to identify the mechanism of R. padi resistance to beta-cypermethrin. Then, these four transcription profiles were divided into two groups for comparative analysis of DEGs. The first group (“Beta-R-T compared with Beta-R”, “SS-T compared with SS”) was the resistant group and sensitive group; when the aphids were briefly induced by beta-cypermethrin, the changes were observed in the various functions of resistant and sensitive strains. The second group (“Beta-R-T compared with SS-T” and “Beta-R compared with SS”) was the beta-cypermethrin-induced group and the non-beta-cypermethrin-induced control group, to locate and screen a large number of drug-resistance genes.
Differential expression analysis was performed, using edgeR [
35], to calculate the
p-value of differential expression. The greater the significance of the gene expression difference, the smaller the
p-value; hence, genes were identified as differentially expressed genes (DEGs). In order to control the false detection rate (FDR), it is necessary to filter genetic variations using a combination of
p-value and FoldChange, with the screening condition being
p-value 0.05 and log2 |FoldChange| > 1. The functional annotation and classification of the genes were performed through the GO database, while biological pathway annotations were obtained by using the KEGG database.
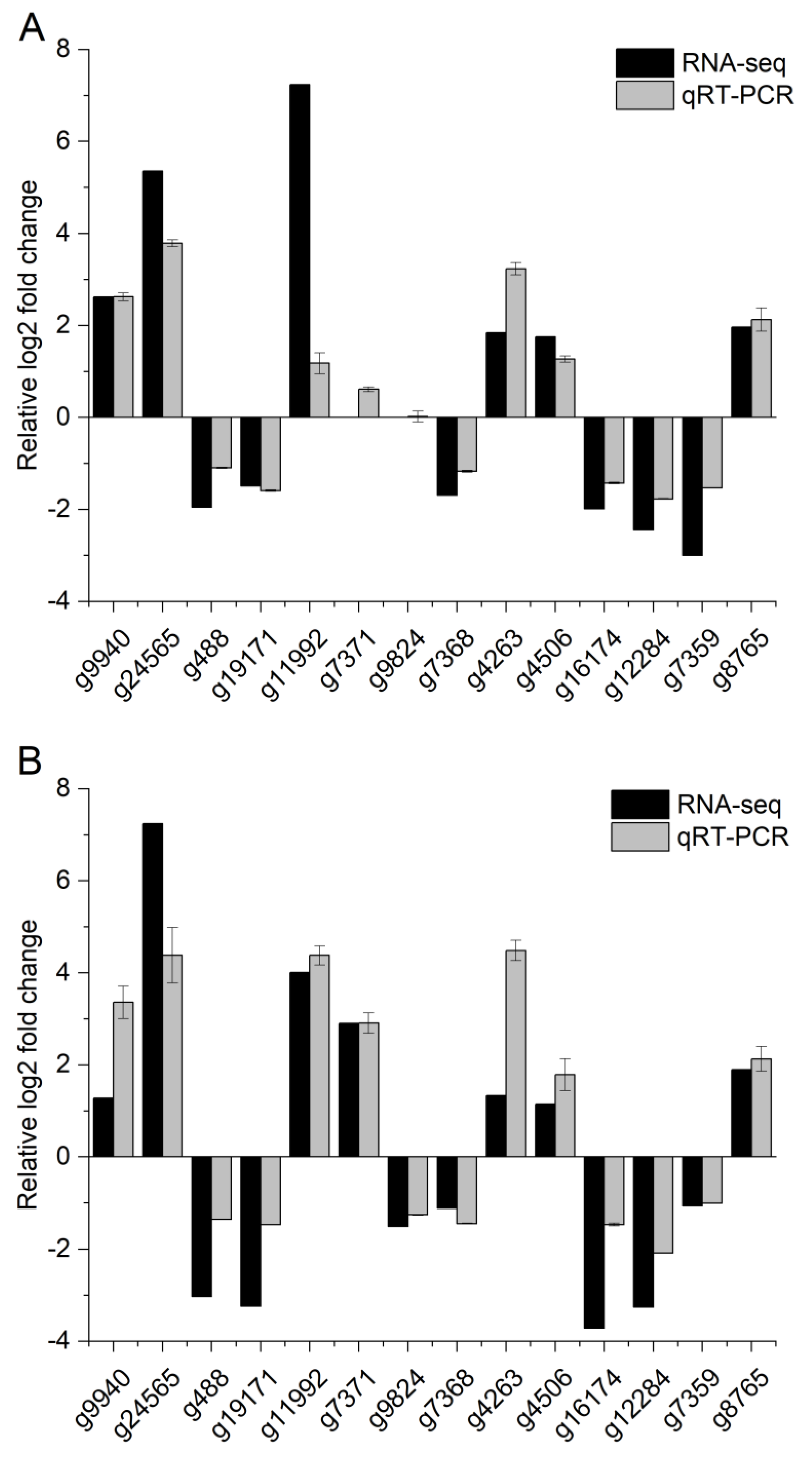
2.7. Quantitative Real-Time PCR Analysis
Quantitative real-time PCR (qRT-PCR) analysis was conducted to validate the expression profiles of 10 randomly selected DEGs and one reference gene, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) genes [
36]. DNAMAN was used to design specific primers to confirm the assembled sequences, followed by the use of a simple technique of reverse transcription PCR (the primers used in qRT-PCR are summarized in
Table 1). Quantitative reverse transcription PCR (qRT-PCR) reactions were performed by using the reagent SuperReal PreMix Plus (SYBP Green) (Tiangen, Beijing, China) on a CFX Connect™ Fluorescent quantitative PCR detection system (Bio-Rad, Hercules, CA, USA). All qRT-PCR experiments were performed in triplicate using independent samples. The expression levels were determined by the 2
−△△Ct method [
37], using the geometric mean of the reference gene for normalization [
36].
4. Discussion
A common approach in studying the physiological adaptations that lead to insect resistance to insecticides is to compare the transcriptome of resistant strains to that of sensitive strains [
38]. In our transcriptome experiment, to screen more comprehensive resistance-related genes, we treated the existing
R. padi’s beta-cypermethrin resistant and sensitive strains with short-term agent treatment, respectively, to obtain drug-induced strains of resistant and sensitive strains.
Firstly, we conducted expression analysis and functional enrichment of GO and KEGG by comparing the two transcriptomes of DEGs before and after induction of antagonistic strains and sensitive strains. In this way, it is easier to expose the rapid changes in expression and function of resistance genes after encountering agents to identify the functions that lead to aphid resistance to insecticides. We found that the DEGs of the inducible resistant strain and non-inducible resistant strain treatment comparison groups were significantly less in number than those in the inducible sensitive strain treatment and non-inducible sensitive strain treatment comparison group. Furthermore, almost no DEGs, related to resistance, appeared in the inducible resistance strain treatment and non-inducible resistance strain comparison groups. This result contradicts Wang’s obtained results, where the number of differentially expressed genes in resistant strains was higher than that in sensitive strains [
32]. However, our research found that the expression level of common genes in the resistant groups was significantly higher than sensitive groups. This phenomenon requires further study of the specific reasons and real mechanisms. The functional annotation showed that these genes had different functions, most of which were related to the enhanced activity of detoxification enzymes and structural changes of cuticle proteins.
We then conducted two transcriptome comparisons between uninduced resistant and sensitive strains, and between induced resistant and sensitive strains. We found a large number of resistance-related genes. We found that most DEGs appeared in the comparison of two transcriptomes between non-induced resistant strains and sensitive strains and induced resistant strains and sensitive strains, and the expression levels of some genes changed in the multi-group comparison, and the expression levels changed in the same trend. For example, regarding the cuticle protein gene g9944 and UGT gene g16174, this kind of gene should be studied as the key gene of insecticide resistance. We also found a large number of downregulated genes, which may be a balance to compensate for the fitness cost. This phenomenon is very interesting and requires further research.
The findings could indicate that the mechanism of drug resistance in R. padi is complex, resulting from the joint action of multiple resistance mechanisms. From the contact of the drug with the epidermal protein of the insect body to the digestion of trypsin after entering the insect body, to the final function of the detoxification enzyme gene oxidation hydrolysis upon exiting the body, the process is rigorous and scientific.
The cuticle protein exhibits unique properties and noticeable changes during all kinds of comparisons. The insect cuticle is the first line of defense against external chemicals, specifically the proteinaceous and chitinous procuticle [
39]. Cuticular protein (CP) genes play an important role in insect development and defense [
39]. As such, cuticular proteins have been reported to play crucial roles in the insecticide resistance and tolerance of a variety of insect species, including
Myzus persicae [
40],
Aphis gossypii [
41], and
Plutella xylostella [
42]. Our results demonstrated that epidermal protein-related genes were the most numerous and varied among the insecticide-related DEGs, as shown in
Table 4. The downregulation of epidermal protein genes, as in this study, conforms with previous studies that reported the downregulation of several cuticle protein genes in the imidacloprid-resistant
Myzus persicae [
43]. The cuticular changes underlying insecticide resistance involve two main parameters: thickness and composition of the cuticle [
13]. More significantly, cuticular thickening has been more commonly associated with resistance [
44]; but there are recent reports that link cuticular composition to decreased xenobiotic penetration [
45]. Therefore, on the basis of the results, it is suggested that cuticular reorganization could play a role in insecticide resistance for
D. citri, though the specific roles of the CPs involved in alterations of cuticle structure or composition as specifically related to insecticide penetration require further research.
Cytochrome P450 is an enzyme with a variety of metabolic functions, and it is involved in detoxifying foreign chemicals such as pesticides [
46]. The overexpression of the P450 monooxygenase enzyme is the most common mechanism of resistance to beta-cypermethrin in many insects [
47,
48,
49]. In aphids, the P450 monooxygenase enzyme also plays an important role in insecticide detoxification and resistance [
46,
50,
51]. In the present study, different treatments generated different DEGs; among these, the most P450s-related downregulated single genes were generated in the inducible resistant strain treatment and inducible sensitive strain treatment comparison group (17). Most of the P450-related upregulated single genes were generated in non-inducible resistant strain treatment and non-inducible sensitive strain treatment comparison group (6) (most of which were labelled as CYP4 and CYP6 family genes, as presented in
Supplementary File S8). Both genes (CYP4 and CYP6) were more associated with pesticides than any other P450 family, suggesting that these genes play an important role in pesticide metabolism and detoxification [
52,
53]. Therefore, overexpression of these two genes may be related to the resistance and detoxification process of
R. padi to beta-cypermethrin. For example, the CYP4 family gene of
Aedes albopictus plays an important role in pyrethroid detoxification [
54]. In houseflies, the CYP6 family gene was overexpressed in pyrethroid-resistant populations, which proved to be related to the pyrethroid resistance of houseflies [
55]. Similarly, transcriptional overexpression of CYP6 family genes, associated with pyrethroid resistance, was also found in
Helicoverpa zea as collected from the field [
56]. Although our results demonstrated that overexpression of CYP4 and CYP6 family genes is associated with the detoxification of beta-cypermethrin in
R. padi, further research is required to determine whether these P450s can metabolize imidacloprid and chlorpyrifos.
UDP-glucosyltransferases (UGTs) are detoxification enzymes that are involved in insecticide metabolism [
57]. The phase Ⅱ enzymes, UGTs act on the toxic by-products of the phase Ⅰ metabolism; UGT-catalyzed conjugation of small lipophilic compounds with sugars is an important detoxification and homeostatic function in all living organisms [
58]. For example, in thiamethoxam-resistant strains of
A. gossypii, UGT was significantly upregulated relative to the susceptible strains [
59], and UGTs were also implicated in the resistance of
Meteorus pulchricornis to commonly used insecticides [
60]. In our experiment, we used qRT-PCR to verify the expression levels of UGTs genes (g4263 g15391, g13098, g16174) that were differentially expressed in transcriptome data; it suggests the involvement of UGTs in beta-cypermethrin metabolism and transport. In the qRT-PCR assay of the annotated UGT detoxification enzymes, we found that g4263 expression was highly upregulated by the qRT-PCR assay of the UGT detoxification enzyme. UGTs play an important role in a variety of physiological and biochemical processes in insects, including the detoxification of substrates (such as plant allelochemicals and insecticides) [
61,
62]; our results indicated that UGTs may play a role in the tolerance and detoxification of insecticides.
ATP-binding cassette (ABC) transporters are a family of transmembrane proteins that mainly rely on ATP to generate energy for substrates. Here, ABC transporters, as the phase Ⅲ detoxification enzymes, play a vital role in the detoxification and defense of various tissues and organs of the insect [
63,
64]. ABC transporters have been associated with pyrethroids resistance in insects [
65]. In this study, insecticide-resistant DEGs were associated with ABC transporters, and most were downregulated single genes. Thus, the ABC transporter may play an important role in the detoxification process and insecticide tolerance of
R. padi. In addition, trypsin-related genes, which accounted for a large group of DEGs in
S. avenae [
66], were upregulated in
Caligus rogercresseyi, following exposure to deltamethrin [
67]. In our study, trypsin DEGs were also found to be related to pyrethroid resistance, indicating that the trypsin enzyme is vital in the detoxification process and insecticide tolerance of
R. padi.
Through this experiment, we found a large number of genes related to insecticide resistance, and we should screen out the key genes of resistance for further study in the follow-up experiments. We found that metabolites are the direct defense line of insects against the harm of drugs and can directly respond to the attack of drugs, to enhance insect resistance. Thus, metabolome analysis can assist transcriptome analysis to more accurately identify key genes associated with resistance [
68]. Therefore, metabolism-related experiments should be included in subsequent experiments to assist in verifying the results of transcriptome analysis.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}