
Temperature Requirements for the Colony Growth and Conidial Germination of Selected Isolates of Entomopathogenic Fungi of the Cordyceps and Paecilomyces Genera

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. Effect of Temperature on Colony Growth
2.3. Effect of Temperature on Conidial Germination
2.4. Statistical Analyses
3. Results
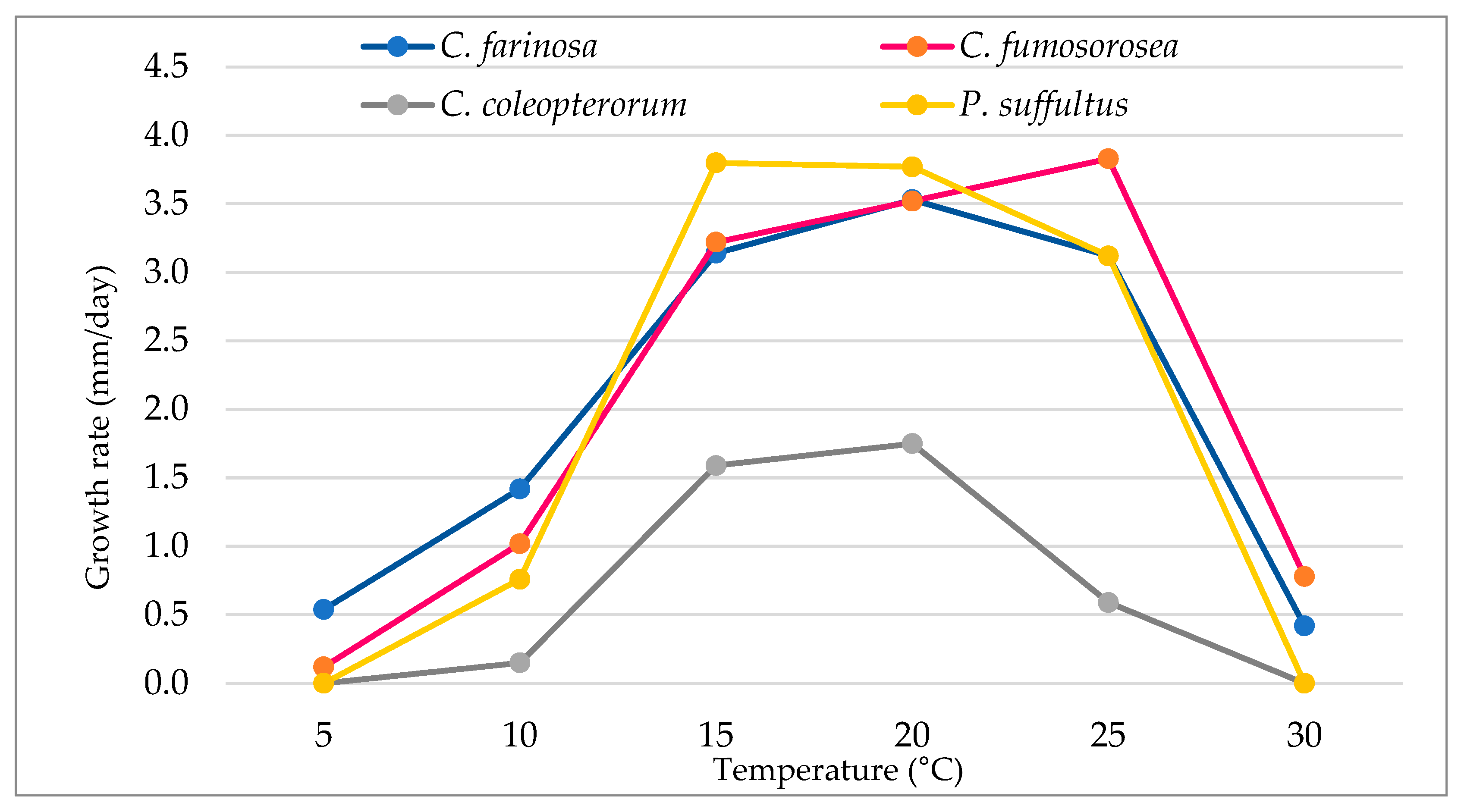
3.1. Effects of Temperature on Mycelial Growth Rate
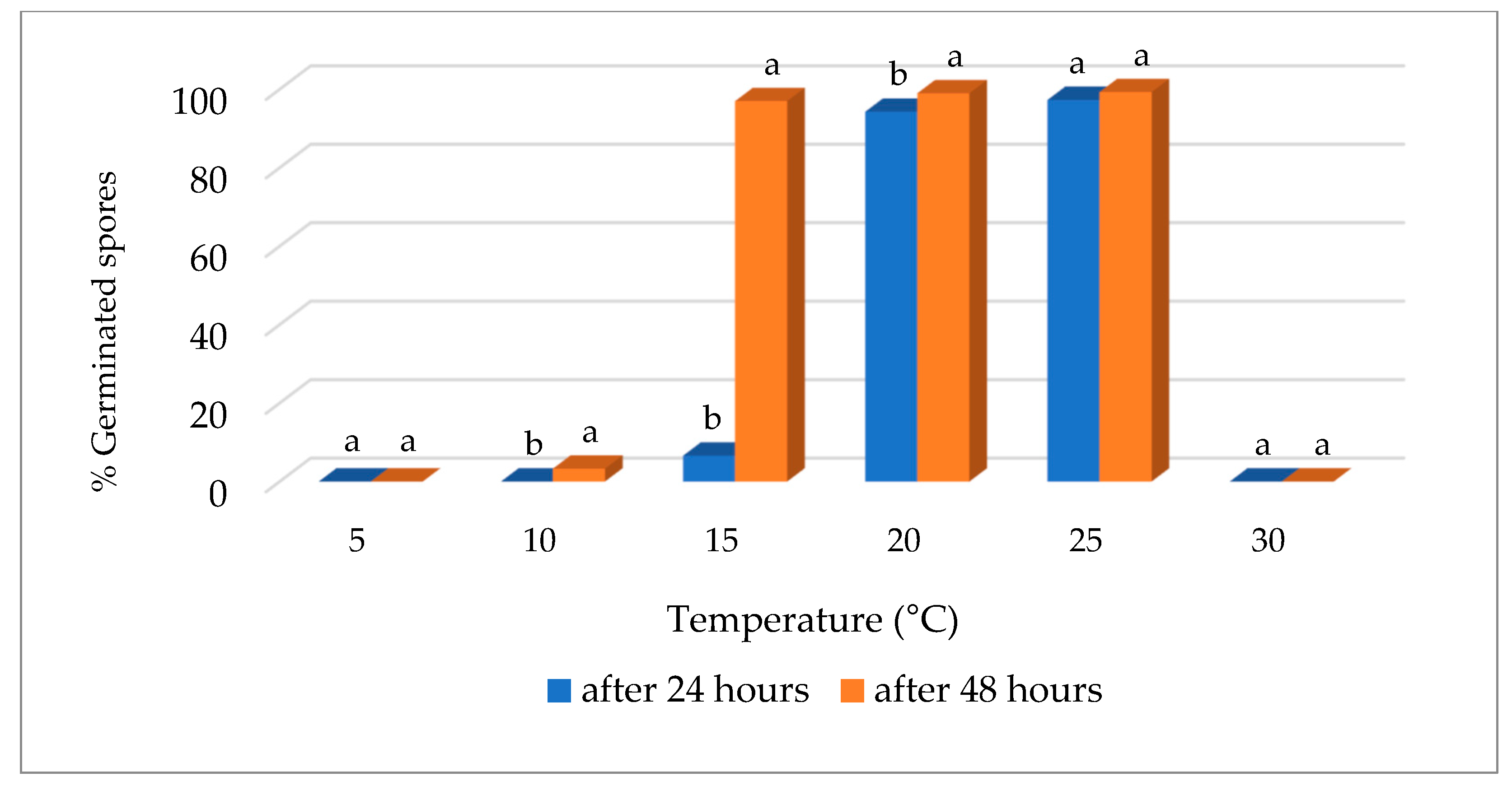
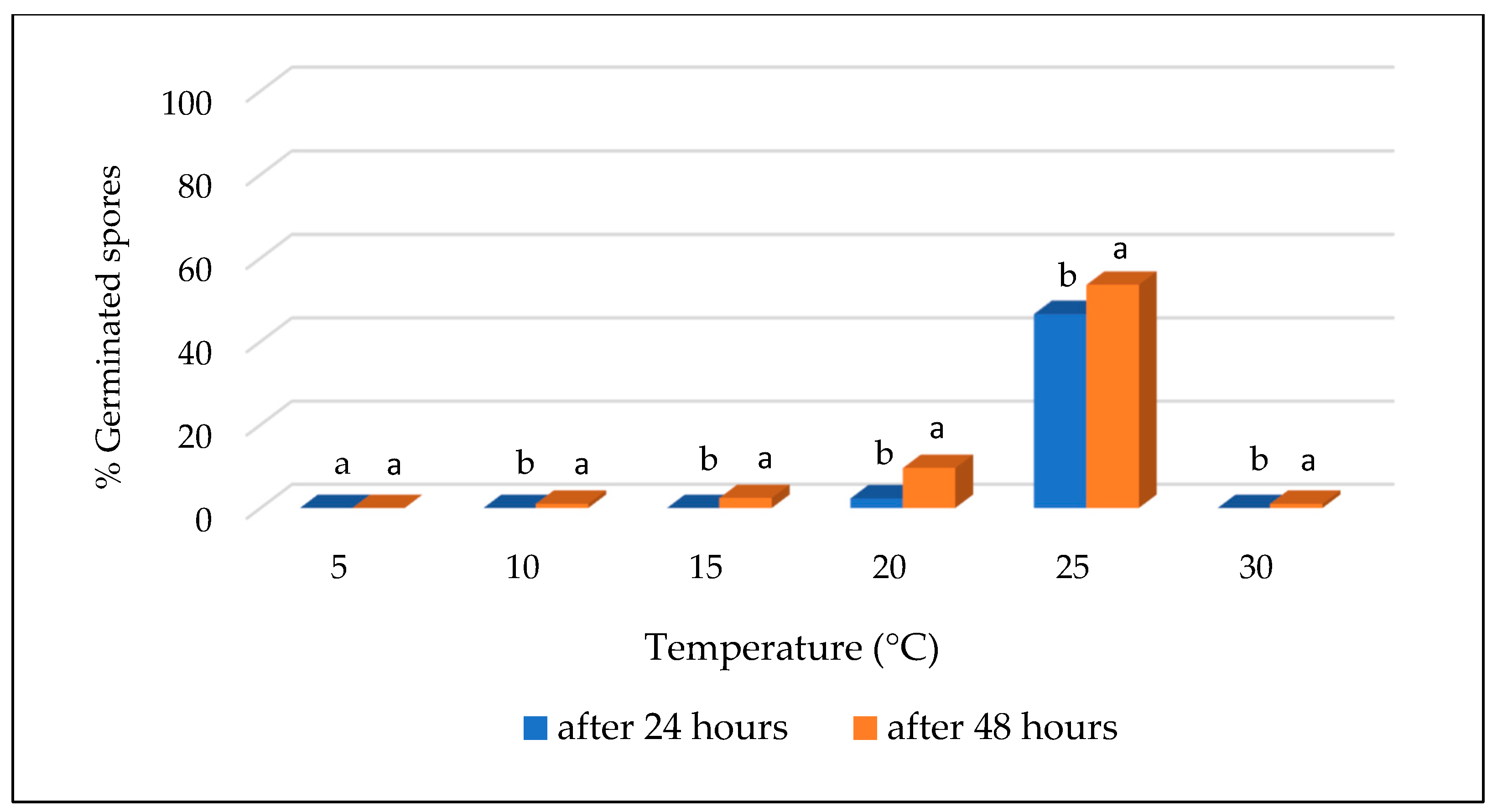
3.2. Effect of Temperature on Conidial Germination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hajek, A.E.; St. Leger, R.J. Interactions between fungal pathogens and insect host. Annu. Rev. Entomol. 1994, 39, 39–293. [Google Scholar] [CrossRef]
- Chandler, D.; Sunderland, K.D.; Ball, B.V.; Davidson, G. Prospective biological control agents of Varroa destructor n. sp., an important pest of the European honeybee, Apis mellifera. Biocontrol Sci. Technol. 2001, 11, 429–448. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Navas-Cortés, J.A.; Maranhao, E.A.; Ortiz-Urquiza, A.; Santiago-Alvarez, C. Factors affecting the occurrence and distribution of entomopathogenic fungi in natural and cultivated soils. Mycol. Res. 2007, 111, 947–966. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A.; Gotz, P. Parasitic fungi and their interactions with the insect immune system. Adv. Parasitol. 1999, 43, 267–313. [Google Scholar] [CrossRef]
- Xiong, Q.; Xiea, Y.; Zhua, Y.; Xuea, J.; Li, J.; Fan, R. Morphological and ultrastructural characterization of Carposina sasakii larvae (Lepidoptera: Carposinidae) infected by Beauveria bassiana (Ascomycota: Hypocreales: Clavicipitaceae). Micron 2013, 44, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, S. Insect pathogenic fungi: Genomics, molecular interactions, and genetic improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Lai, Y.; Wang, G.; Chen, H.; Li, F.; Wang, S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef]
- Gonzalez, F.; Tkaczuk, C.; Dinu, M.M.; Fiedler, Ż.; Vidal, S.; Zchori-Fein, E.; Messelink, G.J. New opportunities for the integration of microorganisms into biological pest control systems in greenhouse crops. J. Pest Sci. 2016, 89, 89–295. [Google Scholar] [CrossRef]
- Wu, S.; Toews, M.D.; Oliveira-Hofman, C.; Behle, R.W.; Simmons, A.M.; Shapiro-Ilan, D.I. Environmental tolerance of entomopathogenic fungi: A new strain of Cordyceps javanica isolated from a whitefly epizootic versus commercial fungal strains. Insects 2020, 11, 711. [Google Scholar] [CrossRef]
- Fargues, J.; Luz, C. Effects of fluctuating moisture and temperature regimes on the infection potential of Beauveria bassiana for Rhodinus prolixus. J. Invertebr. Pathol. 2000, 75, 202–211. [Google Scholar] [CrossRef]
- Wu, S.; Youngman, R.R.; Kok, L.T.; Laub, C.A.; Pfeiffer, D.G. Interaction between entomopathogenic nematodes and entomopathogenic fungi applied to third instar southern masked chafer white grubs, Cyclocephala lurida (Coleoptera: Scarabaeidae), under laboratory and greenhouse conditions. Biol. Control 2014, 76, 65–73. [Google Scholar] [CrossRef]
- Hajek, A.E.; Meyling, N.V. Fungi. In Ecology of Invertebrate Diseases; Hajek, A.E., Shapiro-Ilan, D.I., Eds.; Wiley: Oxford, UK, 2017; pp. 327–377. [Google Scholar]
- Devi, K.U.; Sridevi, V.; Mohan, C.M.; Padmavathi, J. Effect of high temperature and water stress on in vitro germination and growth in isolates of the entomopathogenic fungus Beauveria bassiana (Bals. ) Vuillemin. J. Invertebr. Pathol. 2005, 88, 181–189. [Google Scholar] [CrossRef]
- Alexandre, T.M.; Alves, L.F.A.; Neves, P.M.O.J.; Alves, S.B. Effect of temperature and poultry litter on Beauveria bassiana (Bals.) Vuill. and Metarhizium anisopliae (Metsch) virulence against the lesser mealworm Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae. Neotrop. Entomol. 2006, 35, 75–82. [Google Scholar] [CrossRef]
- Polar, P.; Aquino de Muro, M.; Kairo, M.T.K.; Moore, D.; Pegram, R.; John, S.A.; Roach-Benn, C. Thermal characteristics of Metarhizium anisopliae isolates important for the development of biological pesticides for the control of cattle ticks. Vet. Parasitol. 2005, 134, 159–167. [Google Scholar] [CrossRef]
- Ibarra-Corteś, K.H.; Guzmán-Franco, A.W.; González-Fernández, H.; Suarez-Espinosa, J.; Baverstock, J. Selection of a fungal isolate for the control of the pink hibiscus mealybug Maconellicoccus hirsutus. Pest. Manag. Sci. 2013, 69, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Ferron, P.; Fargues, J.; Riba, G. Fungi as microbial insecticides against pests. In Handbook of Applied Mycology. Volume 2: Humans, Animals, and Insects; Arora, D.K., Ajello, L., Mukerji, K.G., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1991; pp. 665–706. [Google Scholar]
- Fargues, J.; Goettel, M.S.; Smits, N.; Ouedraog, A. Effect of temperature on vegetative growth of Beauveria bassiana isolates from different origins. Mycologia 1997, 89, 383. [Google Scholar] [CrossRef]
- Vidal, C.; Fargues, J.; Lacey, L.A. Intraspecific variability of Paecilomyces fumosoroseus: Effect of temperature on vegetative growth. J. Invertebr. Pathol. 1997, 70, 18–26. [Google Scholar] [CrossRef]
- Jackson, M.A.; Jaronski, S.T. Production of microsclerotia of the fungal entomopathogen Metarhizium anisopliae and their potential for use as a biocontrol agent for soil-inhabiting Insects. Mycol. Res. 2009, 113, 842–850. [Google Scholar] [CrossRef]
- Jaronski, S.T. Ecological factors in the inundative use of fungal entomopathogens. BioControl 2010, 55, 159–185. [Google Scholar] [CrossRef]
- Zimmermann, G. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol. 2008, 18, 865–901. [Google Scholar] [CrossRef]
- Ropek, D.; Krysa, A.; Rola, A.; Frączek, K. Antagonistic effect of Trichoderma viride on entomopathogenic fungi Beauveria bassiana, Isaria fumosorosea and Metarhizium anisopliae in vitro. Pol. J. Agron. 2014, 16, 57–63. [Google Scholar]
- Luangsa-ard, J.J.; Hywel-Jones, N.L.; Samson, R.A. The order level polyphyletic nature of Paecilomyces sensu lato as revealed through 18S-generated rRNA phylogeny. Mycologia 2004, 96, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Luangsa-ard, J.J.; Hywel-Jones, N.L.; Manoch, L.; Samson, R.A. On the relationships of Paecilomyces sect. Isarioidea species. Mycol. Res. 2005, 109, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Gams, W.; Hodge, K.T.; Samson, R.A.; Korf, R.P.; Seifert, K.A. Proposal to conserve the name Isaria (anamorphic fungi) with a conserved type. Taxon 2005, 54, 537. [Google Scholar] [CrossRef]
- Hodge, K.T.; Gams, W.; Samson, R.A.; Korf, R.P.; Seifert, K.A. Lectotypification and status of Isaria Pers. Taxon 2005, 54, 485–489. [Google Scholar] [CrossRef]
- Kepler, R.M.; Luangsa-Ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R. A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef]
- Vänninen, I. Distribution and occurrence of four entomopathogenic fungi in Finland: Effect of geographical location, habitat type and soil type. Mycol. Res. 1996, 100, 93–101. [Google Scholar] [CrossRef]
- Chandler, D.; Hay, D.; Reid, A.P. Sampling and occurrence of entomopathogenic fungi and nematodes in UK soils. Appl. Soil. Ecol. 1997, 5, 133–141. [Google Scholar] [CrossRef]
- Lacey, L.A.; Wraight, S.P.; Kirk, A.A. Entomopathogenic fungi for control of Bemisia spp. In Classical Biological Control of Bemisia tabaci in the USA: A Review of Interagency Research and Implementation; Progress in Biological Control; Gould, J.K., Hoelmer, K., Goolsby, J., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 4, p. 3369. [Google Scholar]
- Kim, W.G.; Seok, S.J.; Weon, H.Y.; Lee, H.H.; Lee, C.J.; Kim, Y.S. Isolation and identification of entomopathogenic fungi collected from mountains and Islands in Korea. Kor. J. Mycol. 2010, 38, 99–104. [Google Scholar] [CrossRef]
- Samson, R.A. Paecilomyces and some allied Hyphomycetes. Stud. Mycol. 1974, 6, 32–41. [Google Scholar]
- Humber, R.A. Manual of Techniques in Invertebrate Pathology. In Identification of Entomopathogenic Fung; Lacey, L.A., Ed.; Academic Press: London, UK, 2012; pp. 151–187. [Google Scholar]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Kovač, M.; Gorczak, M.; Wrzosek, M.; Tkaczuk, C.; Pernek, M. Identification of entomopathogenic fungi as naturally occurring enemies of the invasive oak lace bug, Corythucha arcuata (Say) (Hemiptera: Tingidae). Insects 2020, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Yeo, H.; Pell, J.K.; Alderson, P.G.; Clark, S.J.; Pye, B.J. Laboratory evaluation of temperature effects on the germination and growth of entomopathogenic fungi and on their pathogenicity to two aphid species. Pest. Manag. Sci. 2003, 59, 56165. [Google Scholar] [CrossRef]
- Piątkowski, J.; Krzyżewska, A. Influence of some physical factors on the growth and sporulation of entomopathogenic fungi. Acta Myc. 2007, 42, 255–265. [Google Scholar] [CrossRef]
- Sapna, M.; Peeyush, K.; Anushree, M. Effect of temperature and humidity on pathogenicity of native Beauveria bassiana isolate against Musca domestica L. J. Parasit. Dis. 2015, 39, 697–704. [Google Scholar] [CrossRef]
- Christos, G.; Athanassiou, N.G.; Kavallieratos, C.I.R.; Demetrius, C.K. Influence of temperature and relative humidity on the insecticidal efficacy of Metarhizium anisopliae against larvae of Ephestia kuehniella (Lepidoptera: Pyralidae) on wheat. J. Insect Sci. 2017, 17, 22. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Jaronski, S.T. Mass production of entomopathogenic fungi—State of the art. In Mass Production of Beneficial Organisms; Academic Press: London, UK, 2023; pp. 317–357. [Google Scholar] [CrossRef]
- Fargues, J.; Goettel, M.S.; Smits, N.; Ouedraogo, A.; Vidal, C.; Lacey, L.A.; Lomer, C.J.; Rougier, M. Variability in susceptibility to simulated sunlight of conidia among isolates of entomopathogenic Hyphomycetes. Mycopathologia 1997, 135, 171–181. [Google Scholar] [CrossRef]
- Doberski, J.W. Comparative laboratory studies on three fungal pathogens of the elm bark beetle Scolytus scolytus: Effect of temperature and humidity on infection by Beauveria bassiana, Metarhizium anisopliae and Paecilomyces farinosus. J. Invertebr. Pathol. 1981, 37, 195–200. [Google Scholar] [CrossRef]
- Machowicz-Stefaniak, Z. Studies on the effect of temperature on the growth and development of the fungus Paecilomyces farinosus (Dicks, ex Fr.) Brown et Smith (Moniliales, Deuteromycetes). Rocz. Nauk. Roln. 1988, 18, 121–130. [Google Scholar]
- Mietkiewski, R.; Tkaczuk, C.; Żurek, M.; Van der Geest, L.P.S. Temperature requirements of four entomopathogenic fungi. Acta Myc. 1994, 29, 109–120. [Google Scholar] [CrossRef]
- Hallsworth, J.E.; Magan, N. 1999. Water and temperature relations of growth of the entomogenous fungi Beauveria bassiana, Metarhizium anisopliae, and Paecilomyces farinosus. J. Invertebr. Pathol. 1999, 74, 261–266. [Google Scholar] [CrossRef]
- Huang, B.; Fan, M.Z.; Li, Z.Z.; Zhou, Q.; Wang, Z.X. Biological characteristics of different strains of Paecilomyces farinosus. J. Anhui Agric. 2005, 32, 8–11. [Google Scholar]
- Brown, H.S.; Smith, G. The genus Paecilomyces Bainier and its perfect stage Byssochlamys Westling. Trans. Br. Mycol. 1957, 40, 17–89. [Google Scholar] [CrossRef]
- Ayyasamy, R.; Baskaran, P. Effect of temperature and relative humidity on radial growth and sporulation of Paecilomyces farinosus. J. Food Agric. Environ. 2005, 3, 137–138. [Google Scholar] [CrossRef]
- Tian, J.; Hao, C.; Liang, L.; Ma, R.Y. Effects of temperature and relative humidity on conidial germination of Isaria fumosorosea (Hypocreales: Cordycipitaceae) IF-1106 and pathogenicity of the fungus against Bemisia tabaci (Homoptera: Aleyrodidae). Mycosystema 2014, 33, 668–679. [Google Scholar] [CrossRef]
- Rojas, V.M.A.; Iwanicki, N.S.A.; D’Alessandro, C.P.; Fatoretto, M.B.; Demétrio, C.G.B.; Delalibera, I., Jr. Characterization of Brazilian Cordyceps fumosorosea isolates: Conidial production, tolerance to ultraviolet-B radiation, and elevated temperature. J. Invertebr. Pathol. 2023, 197, 107888. [Google Scholar] [CrossRef]
- Fargues, J.; Maniania, N.K.; Delmas, J.C.; Smits, N. Influence de la temperature sur la croissance in vitro dhyphomycetes entomopathogenes. Agronomie 1992, 12, 557–564. [Google Scholar] [CrossRef]
- Sosnowska, D.; Ratajkiewicz, H. Assessment of the potential of local entomopathogenic fungi isolates as biocontrol agents. Prog. Plant Prot. 2021, 61, 121–127. [Google Scholar] [CrossRef]
- Bouamama, N.; Vidal, C.; Fargues, J. Effects of fluctuating moisture and temperature regimes on the persistence of quiescent conidia of Isaria fumosorosea. J. Invertebr. Pathol. 2010, 105, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Tigano-Milani, M.S.; Carneiro, R.G.; De Faria, M.R.; Frazao, H.S.; McCoy, C.W. Isozyme characterization and pathogenicity of Paecilomyces fumosoroseus and P. lilacinus to Diabrotica speciosa (Coleoptera: Chrysomelidae) and Meloidogyne javanica (Nematoda: Tylenchidae). Biol. Control. 1995, 5, 378382. [Google Scholar] [CrossRef]
- Cantone, F.A.; Vandenberg, J.D. Intraspecific diversity in Paecilomyces fumosoroseus. Mycol. Res. 1998, 102, 209–215. [Google Scholar] [CrossRef]
- Fargues, J.; Bon, M.C.; Manguin, S.; Couteaudier, I. Genetic ariability amvong Paecilomyces fumosoroseus isolates from various geographical and host insect origins based on the rDNA-ITS regions. Mycol. Res. 2002, 106, 1066–1074. [Google Scholar] [CrossRef]
- Fargues, J.; Bon, M.C. Influence of temperature preferences of two Paecilomyces fumosoroseus lineages on their co-infection pattern. J. Invertebr. Pathol. 2004, 87, 94–104. [Google Scholar] [CrossRef]
- Gauthier, N.; Dalleau-Clouet, C.; Fargues, J.; Bon, M.C. Microsatellite variability in the entomopathogenic fungus Paecilomyces fumosoroseus: Genetic diversity and population structure. Mycologia 2007, 99, 693–704. [Google Scholar] [CrossRef]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; et al. Insect-fungal-interactions: A detailed review on entomopathogenic fungi pathogenicity to combat insect pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef]
- Tkaczuk, C. Occurrence and Infective Potential of Entomopathogenic Fungi in Soils of Agrocenoses and Seminatural Habitats in the Agricultural Landscape; Scientific Dissertation No. 94; Wydawnictwo Akademii Podlaskiej: Siedlce, Poland, 2008; p. 160. (In Polish) [Google Scholar]
- Petch, T. Notes on entomogenous fungi. Trans. Br. mycol. Soc. 1944, 27, 81–93. [Google Scholar] [CrossRef]
- Sosnowska, D.; Balazy, S.; Prishchepa, L.; Mikulskaya, N. Biodiversity of arthropod pathogens in the Bialowieża Forest. J. Plant Prot. Res. 2004, 44, 313–321. [Google Scholar]
- Żurek, M.; Miętkiewski, R.; Tkaczuk, C. The susceptibility of Bibio hortulanus L. larvae to entomopathogenic fungi in soil. In Invertebrate Pathogen in Biological Control: Presen and Future; IOBC: Bari, Italy, 2005; p. 165. [Google Scholar]
- Bałazy, S. Znaczenie obszarów chronionych dla zachowania zasobów grzybów entomopatogenicznych. Kosmos 2004, 53, 5–16. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Species and Strain Number | Isolate Origin |
|---|---|
| Cordyceps farinosa (P04-UPH) | From the pupa of an unmarked butterfly species, found in oak-hornbeam forest litter of the Białowieża National Park, Poland |
| Cordyceps fumosorosea (P03-UPH) | From Galleria mellonella larvae from the soil of an arable field in Pietrusy (Mazowieckie Voivodeship), Poland |
| Cordyceps coleopterorum (P07-UPH) | From the larvae of a beetle from the firefly family (Lampyridae), found in the litter of an oak-hornbeam forest in the Białowieża National Park, Poland |
| Paecilomyces suffultus (P06-UPH) | From the larvae of the Bibio sp., found in the litter of a typical oak-hornbeam forest in the Białowieża National Park, Poland |
| Fungi Species | Temperature | |||||
|---|---|---|---|---|---|---|
| 5 °C | 10 °C | 15 °C | 20 °C | 25 °C | 30 °C | |
| Cordyceps farinosa | 9.75 ± 0.81 a | 25.7 ± 1.9 a | 56.6 ± 1.8 b | 63.7 ± 3.5 a | 56.3 ± 2.5 b | 7.5 ± 1.3 b |
| Cordyceps fumosorosea | 2.12 ± 0.32 b | 18.4 ± 1.3 b | 58.0 ± 2.1 b | 63.3 ± 1.6 a | 69.0 ± 1.8 a | 14.5 ± 1.1 a |
| Cordyceps coleopterorum | 0.0 ± 0.0 c | 2.75 ± 0.35 d | 28.7 ± 1.4 c | 31.5 ± 0.5 b | 10.7 ± 0.95 c | 0.0 ± 0.0 c |
| Paecilomyces suffultus | 0.0 ± 0.0 c | 13.7 ± 1.33 c | 68.5 ± 0.76 a | 68.0 ± 1.0 a | 56.2 ± 1.4 b | 0.0 ± 0.0 c |
| p-value | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| F-value | 368.04 | 824.68 | 3502.79 | 4226.46 | 1642.69 | 2488.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tkaczuk, C.; Majchrowska-Safaryan, A. Temperature Requirements for the Colony Growth and Conidial Germination of Selected Isolates of Entomopathogenic Fungi of the Cordyceps and Paecilomyces Genera. Agriculture 2023, 13, 1989. https://doi.org/10.3390/agriculture13101989
Tkaczuk C, Majchrowska-Safaryan A. Temperature Requirements for the Colony Growth and Conidial Germination of Selected Isolates of Entomopathogenic Fungi of the Cordyceps and Paecilomyces Genera. Agriculture. 2023; 13(10):1989. https://doi.org/10.3390/agriculture13101989
Chicago/Turabian StyleTkaczuk, Cezary, and Anna Majchrowska-Safaryan. 2023. "Temperature Requirements for the Colony Growth and Conidial Germination of Selected Isolates of Entomopathogenic Fungi of the Cordyceps and Paecilomyces Genera" Agriculture 13, no. 10: 1989. https://doi.org/10.3390/agriculture13101989