
Soil Microbial Community Responses to Cyanobacteria versus Traditional Organic Fertilizers
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Soil Sampling
2.2. Ester-Linked Fatty Acid Methyl Ester (EL-FAME) Analysis
2.3. Statistical Analysis
3. Results
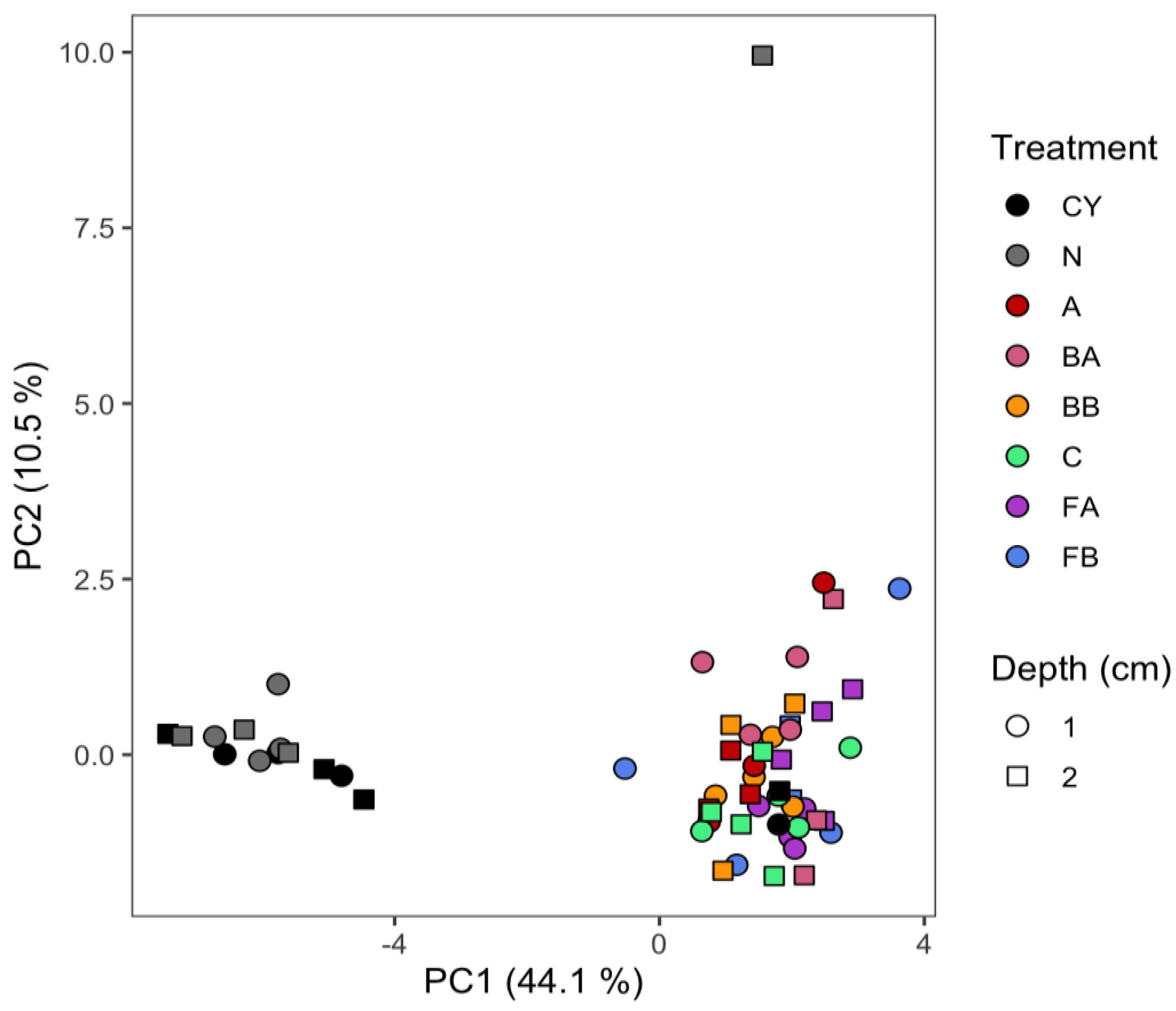
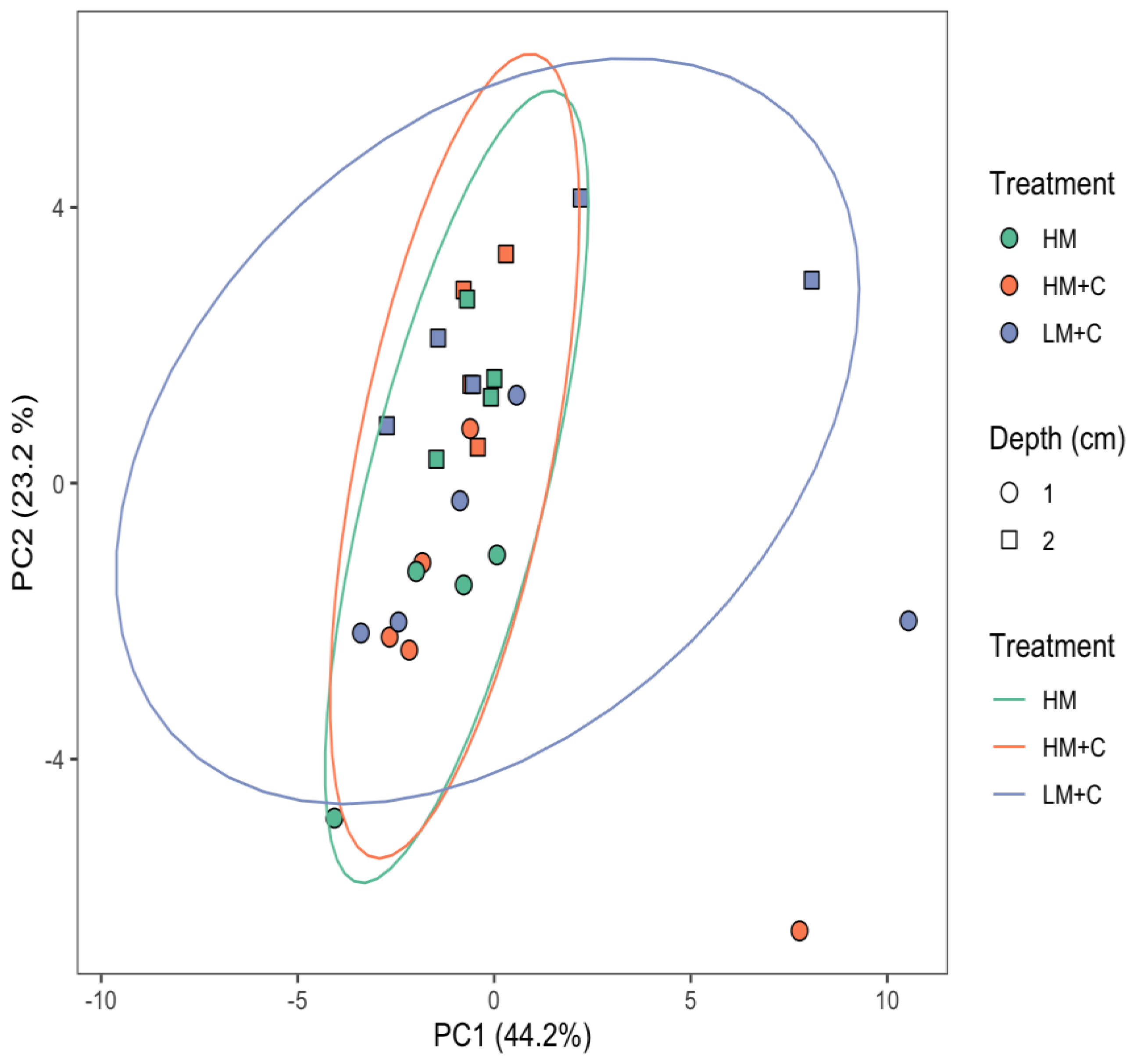
3.1. Microbial Community Structure
3.2. Microbial Community Biomass
4. Discussion
4.1. Microbial Community Responses to Fertilizers
4.2. Microbial Community Groups under Two Different Depths in Cucumber and Peach Soils
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dixon, M.; Rohrbaugh, C.; Afkairin, A.; Vivanco, J. Impacts of the green revolution on rhizosphere microbiology related to nutrient acquisition. Appl. Microbiol. 2022, 2, 992–1003. [Google Scholar] [CrossRef]
- Abed, R.M.M.; Dobretsov, S.; Sudesh, K. Applications of cyanobacteria in biotechnology. J. Appl. Microbiol. 2009, 106, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Crecchio, C.; Curci, M.; Mininni, R.; Ricciuti, P.; Ruggiero, P. Short-term effects of municipal solid waste compost amendments on soil carbon and nitrogen content, some enzyme activities and genetic diversity. Biol. Fertil. Soils 2001, 34, 311–318. [Google Scholar] [CrossRef]
- Ebhin Masto, R.; Chhonkar, P.K.; Singh, D.; Patra, A.K. Changes in soil biological and biochemical characteristics in a long-term field trial on a sub-tropical inceptisol. Soil Biol. Biochem. 2006, 38, 1577–1582. [Google Scholar] [CrossRef]
- Kidd, J.; Manning, P.; Simkin, J.; Peacock, S.; Stockdale, E. Impacts of 120 years of fertilizer addition on a temperate grassland ecosystem. PLoS ONE 2017, 12, e0174632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hao, X.; Alexander, T.W.; Thomas, B.W.; Shi, X.; Lupwayi, N.Z. Long-term and legacy effects of manure application on soil microbial community composition. Biol. Fertil. Soils 2018, 54, 269–283. [Google Scholar] [CrossRef]
- Franco-Otero, V.G.; Soler-Rovira, P.; Hernández, D.; López-de-Sá, E.G.; Plaza, C. Short-term effects of organic municipal wastes on wheat yield, microbial biomass, microbial activity, and chemical properties of soil. Biol. Fertil. Soils 2011, 48, 205–216. [Google Scholar] [CrossRef]
- Marschner, P.; Kandeler, E.; Marschner, B. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem. 2003, 35, 453–461. [Google Scholar] [CrossRef]
- Yoder, N.; Davis, J.G. Organic fertilizer comparison on growth and nutrient content of three kale cultivars. HortTechnology 2020, 30, 176–184. [Google Scholar] [CrossRef]
- Barminski, R.; Storteboom, H.; Davis, J.G. Development and evaluation of an organically certifiable growth medium for cultivation of cyanobacteria. J. Appl. Phycol. 2016, 28, 2623–2630. [Google Scholar] [CrossRef]
- Wolde, G.; Asmamaw, M.; Sido, M.Y.; Yigrem, S.; Wolde-meskel, E.; Chala, A.; Storteboom, H.; Davis, J.G. Optimizing a cyanobacterial biofertilizer manufacturing system for village-level production in Ethiopia. J. Appl. Phycol. 2020, 32, 3983–3994. [Google Scholar] [CrossRef]
- Afkairin, A.; Ippolito, J.A.; Stromberger, M.; Davis, J.G. Solubilization of organic phosphorus sources by cyanobacteria and a commercially available bacterial consortium. Appl. Soil Ecol. 2021, 162, 103900. [Google Scholar] [CrossRef]
- Asmamaw, M.; Wolde, G.; Yohannes, M.; Yigrem, S.; Woldemeskel, E.; Chala, A.; Davis, J.G. Comparison of cyanobacterial bio-fertilizer with urea on three crops and two soils of Ethiopia. Afr. J. Agric. Res. 2019, 14, 588–596. [Google Scholar] [CrossRef]
- Toonsiri, P.; Del Grosso, S.J.; Sukor, A.; Davis, J.G. Greenhouse Gas Emissions from solid and liquid organic fertilizers applied to lettuce. J. Environ. Qual. 2016, 45, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Erwiha, G.M.; Ham, J.; Sukor, A.; Wickham, A.; Davis, J.G. Organic fertilizer source and application method impact ammonia volatilization. Commun. Soil Sci. Plant Anal. 2020, 51, 1469–1482. [Google Scholar] [CrossRef]
- Dhuldhaj, U.; Pandya, U. Implementation of biofortification technology by using PGPR for sustainable agricultural production. Agric. Important Microbes Sustain. Agric. 2017, 2, 63–79. [Google Scholar] [CrossRef]
- Kumar, M.; Singh, D.P.; Prabha, R.; Sharma, A.K. Role of Cyanobacteria in nutrient cycle and use efficiency in the soil. In Nutrient Use Efficiency: From Basics to Advances; Springer: New Delhi, India, 2015; pp. 163–171. [Google Scholar] [CrossRef]
- Wenz, J.; Davis, J.G.; Storteboom, H. Influence of light on endogenous phytohormone concentrations of a nitrogen-fixing anabaena Sp. cyanobacterium culture in open raceways for use as fertilizer for horticultural crops. J. Appl. Phycol. 2019, 31, 3371–3384. [Google Scholar] [CrossRef]
- Sukor, A.; Amer, F.S.M.; Vanamala, J.; Davis, J.G. Phytohormones in organic fertilizers influence β-carotene concentration and marketable yield of lettuce (Lactuca sativa). Acta Hortic. 2022, 1348, 15–22. [Google Scholar] [CrossRef]
- Schutter, M.E.; Dick, R.P. Comparison of fatty acid methyl ester (FAME) methods for characterizing microbial communities. Soil Sci. Soc. Am. J. 2000, 64, 1659–1668. [Google Scholar] [CrossRef]
- Li, C.; Cano, A.; Acosta-Martinez, V.; Veum, K.S.; Moore-Kucera, J.A. Comparison between fatty acid methyl ester profiling methods (PLFA and EL-FAME) as soil health indicators. Soil Sci. Soc. Am. J. 2020, 84, 1153–1169. [Google Scholar] [CrossRef]
- Stromberger, M.; Shah, Z.; Westfall, D. Soil microbial communities of no-till dryland agroecosystems across an evapotranspiration Gradient. Appl. Soil Ecol. 2007, 35, 94–106. [Google Scholar] [CrossRef]
- Peacock, A.; Mullen, M.; Ringelberg, D.B.; Tyler, D.D.; Hedrick, D.B.; Gale, P.M.; White, D.C. Soil microbial community responses to dairy manure or ammonium nitrate applications. Soil Biol. Biochem. 2001, 33, 1011–1019. [Google Scholar] [CrossRef]
- Balkwill, D.L.; Leach, F.R.; Wilson, J.T.; Mcnabb, J.F.; White, D.C. Equivalence of Microbial biomass measures based on membrane lipid and cell wall components, adenosine triphosphate, and direct counts in subsurface aquifer sediments. Ecotoxicol. Environ. Saf. 1988, 16, 73–84. [Google Scholar] [CrossRef]
- Guckert, J.B.; Ringelberg, D.B.; White, D.C.; Hanson, R.S.; Bratina, B.J. Membrane fatty acids as phenotypic markers in the polyphasic taxonomy of methylotrophs within the proteobacteria. Microbiology 1991, 137, 2631–2641. [Google Scholar] [CrossRef] [PubMed]
- Vestal, J.R.; White, D.C. Lipid Analysis in Microbial Ecology. BioScience 1989, 39, 535–541. [Google Scholar] [CrossRef]
- Steger, K.; Jarvis, Å.; Smårs, S.; Sundh, I. Comparison of signature lipid methods to determine microbial community structure in compost. J. Microbiol. Methods 2003, 55, 371–382. [Google Scholar] [CrossRef]
- Schutter, M.; Sandeno, J.; Dick, R. Seasonal, Soil type, and alternative management influences on microbial communities of vegetable cropping systems. Biol. Fertil. Soils 2001, 34, 397–410. [Google Scholar] [CrossRef]
- Hsu, S.-F.; Buckley, D.H. Evidence for the functional significance of diazotroph community structure in soil. ISME J. 2008, 3, 124–136. [Google Scholar] [CrossRef]
- Bending, G.D.; Turner, M.K.; Jones, J.E. Interactions between crop residue and soil organic matter quality and the functional diversity of soil microbial communities. Soil Biol. Biochem. 2002, 34, 1073–1082. [Google Scholar] [CrossRef]
- Amador, J.A.; Atoyan, J.A. Structure and composition of leach field bacterial communities: Role of soil texture, depth and septic tank effluent inputs. Water 2012, 4, 707–719. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth Profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Wickham, A.; Davis, G.J. Optimizing organic carrot (Daucus carota var. sativus) yield and quality using fish emulsions, cyanobacterial fertilizer, and seaweed extracts. Agronomy 2023, 13, 1329. [Google Scholar] [CrossRef]
- Soil Science|Natural Resources Conservation Service. Available online: https://www.nrcs.usda.gov/conservation-basics/natural-resource-concerns/soil/soil-science (accessed on 16 August 2023).
- Sterle, D.G.; Stonaker, F.; Ela, S.; Davis, J.G. Cyanobacterial biofertilizer as a supplemental fertilizer for peaches: Yield, trunk growth, leaf nutrients and chlorosis. J. Am. Pomol. Soc. 2021, 75, 165–175. [Google Scholar]
- Frostegard, A.; Tunlid, A.; Baath, E. Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types experimentally exposed to different heavy metals. Appl. Environ. Microbiol. 1993, 59, 3605–3617. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.D.; Denef, K.; Stewart, C.E.; Zheng, J.; Cotrufo, M.F. Biochar addition rate influences soil microbial abundance and activity in temperate soils. Eur. J. Soil Sci. 2014, 65, 28–39. [Google Scholar] [CrossRef]
- Denef, K.; Roobroeck, D.; Manimel Wadu, M.C.W.; Lootens, P.; Boeckx, P. Microbial community composition and rhizodeposit-carbon assimilation in differently managed temperate grassland soils. Soil Biol. Biochem. 2009, 41, 144–153. [Google Scholar] [CrossRef]
- Grogan, D.W.; Cronan, J.E., Jr. Cyclopropane ring formation in membrane lipids of bacteria. Microbiol. Mol. Biol. Rev. 1997, 61, 429–441. [Google Scholar]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, Aka Least-Squares Means (Version 1.3.4). Emmeans Estim Marg Means Aka Least-Sq Means. 2019. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 10 April 2019).
- Esaki, K.; Ichinose, Y.; Yamada, S. Statistical analysis of process monitoring data for software process improvement and its application. Am. J. Oper. Res. 2012, 2, 43–50. [Google Scholar] [CrossRef]
- Frey, S.D.; Elliott, E.T.; Paustian, K. Bacterial and fungal abundance and biomass in conventional and no-tillage agroecosystems along two climatic gradients. Soil Biol. Biochem. 1999, 31, 573–585. [Google Scholar] [CrossRef]
- Wienhold, B.; Halvorson, A. Cropping system influences on several soil quality attributes in the northern great Plains. J. Soil Water Conserv. 1998, 53, 254–258. [Google Scholar]
- Yevdokimov, I.; Gattinger, A.; Buegger, F.; Munch, J.C.; Schloter, M. Changes in microbial community structure in soil as a result of different amounts of nitrogen fertilization. Biol. Fertil. Soils 2008, 44, 1103–1106. [Google Scholar] [CrossRef]
- Kimura, M.; Asakawa, S. Comparison of community structures of microbiota at main habitats in rice field ecosystems based on phospholipid fatty acid analysis. Biol. Fertil. Soils 2005, 43, 20–29. [Google Scholar] [CrossRef]
- Lundquist, E.J.; Scow, K.M.; Jackson, L.E.; Uesugi, S.L.; Johnson, C.R. Rapid response of soil microbial communities from conventional, low input, and organic farming systems to a wet/dry cycle. Soil Biol. Biochem. 1999, 31, 1661–1675. [Google Scholar] [CrossRef]
- MacKenzie, M.D.; Quideau, S.A. Microbial Community structure and nutrient availability in oil sands reclaimed boreal soils. Appl. Soil Ecol. 2010, 44, 32–41. [Google Scholar] [CrossRef]
- Lazcano, C.; Gómez-Brandón, M.; Revilla, P.; Domínguez, J. Short-term effects of organic and inorganic fertilizers on soil microbial community structure and function. Biol. Fertil. Soils 2012, 49, 723–733. [Google Scholar] [CrossRef]
- Ranjan, K.; Priya, H.; Ramakrishnan, B.; Prasanna, R.; Venkatachalam, S.; Thapa, S.; Tiwari, R.; Nain, L.; Singh, R.; Shivay, Y.S. Cyanobacterial inoculation modifies the rhizosphere microbiome of rice planted to a tropical alluvial soil. Appl. Soil Ecol. 2016, 108, 195–203. [Google Scholar] [CrossRef]
- Dinesh, R.; Srinivasan, V.; Hamza, S.; Manjusha, A. Short-term incorporation of organic manures and biofertilizers influences biochemical and microbial characteristics of soils under an annual crop [Turmeric (Curcuma longa L.)]. Bioresour. Technol. 2010, 101, 4697–4702. [Google Scholar] [CrossRef]
- Zeng, L.S.; Liao, M.; Chen, C.L.; Huang, C.Y. Effects of lead contamination on soil enzymatic activities, microbial biomass, and rice physiological indices in soil-lead-rice (Oryza sativa L.) system. Ecotoxicol. Environ. Saf. 2007, 67, 67–74. [Google Scholar] [CrossRef]
- Murate, N.; Nishida, I. Lipids of blue-Green algae (Cyanobacteria). Lipids Struct. Funct. 1987, 9, 315–347. [Google Scholar] [CrossRef]
- Gunstone, F.D.; Harood, J.L. The Lipid Handbook, 2nd ed.; Chapman & Hall: London, UK, 2021; pp. 1–1273. Available online: https://books.google.com/books?hl=en&lr=&id=INZa6WmqDA8C&oi=fnd&pg=PP1&dq=Gunstone,+F.D.+and+Harwood,+J.L.,+2007.+The+lipid+handbook+with+CD-ROM.+CRC+press.&ots=Sv3z6c8Ay1&sig=ueCDpgrIFv7RluBvF14sfdumHR0#v=onepage&q=Gunstone%2CF.D.andHarwood%2CJ.L.%2C (accessed on 27 July 2021).
- Willers, C.; Jansen van Rensburg, P.J.; Claassens, S. Phospholipid fatty acid profiling of microbial communities–a review of interpretations and recent applications. J. Appl. Microbiol. 2015, 119, 1207–1218. [Google Scholar] [CrossRef]
- Gugger, M.; Lyra, C.; Suominen, I.; Tsitko, I.; Humbert, J.-F.; Salkinoja-Salonen, M.S.; Sivonen, K. Cellular fatty acids as chemotaxonomic markers of the genera anabaena, aphanizomenon, microcystis, nostoc and planktothrix (cyanobacteria). Int. J. Syst. Evol. Microbiol. 2002, 52, 1007–1015. [Google Scholar] [CrossRef]
- Li, R.; Watanabe, M.M. Fatty Acid profiles and their chemotaxonomy in planktonic species of anabaena (cyanobacteria) with straight trichomes. Phytochemistry 2001, 57, 727–731. [Google Scholar] [CrossRef]
- Girvan, M.S.; Bullimore, J.; Pretty, J.N.; Osborn, A.M.; Ball, A.S. Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soils. Appl. Environ. Microbiol. 2003, 69, 1800–1809. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Leemans, D.K.; Cook, R.; Hobbs, P.J. Seasonality of the soil biota of grazed and ungrazed hill grasslands. Soil Biol. Biochem. 1997, 29, 1285–1294. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Whiteley, A.S.; O’Donnell, A.G.; Bailey, M.J. Influence of depth and sampling time on bacterial community structure in an upland grassland soil. FEMS Microbiol. Ecol. 2003, 43, 35–43. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Eigenvector Coefficient | |||
|---|---|---|---|
| Microbial Group | EL-FAME | PC1 | PC2 |
| Actinomycetes | 10Me16:0 | −0.574 | −0.032 |
| 10Me17:0 | −0.970 | 0.025 | |
| 10Me18:0 | −0.934 | 0.036 | |
| Fungi | 18:1ω9 | 0.653 | 0.570 |
| 18:2ω6 | 0.200 | −0.568 | |
| AM Fungi | 16:1ω5 | −0.169 | −0.665 |
| Gram-negative Bacteria | 16:1ω7 | −0142 | −0.536 |
| 17:1ω7 | −0.824 | 0.068 | |
| 17:0cy | −0.904 | 0.015 | |
| 18:1ω7 | −0.080 | −0.206 | |
| 18:1ω8 | −0.899 | 0.013 | |
| 19:0cy | −0.412 | 0.223 | |
| Gram-positive Bacteria | i14:0 | 0.259 | −0.368 |
| i15:0 | 0.001 | −0.668 | |
| a15:0 | 0.279 | −0.025 | |
| i16:0 | −0.156 | −0.156 | |
| i17:0 | −0.725 | −0.001 | |
| a17:0 | −0.755 | −0.024 | |
| Non-specific | 13:0 | −0.937 | 0.043 |
| 14:0 | −0.835 | 0.414 | |
| 15:0 | −0.965 | 0.021 | |
| 16:0 | 0.836 | 0.107 | |
| 18:0 | −0.245 | 0.273 | |
| Eigenvector Coefficient | |||
|---|---|---|---|
| Microbial Group | EL-FAME | PC1 | PC2 |
| Actinomycetes | 10Me16:0 | −0.103 | 0.697 |
| 10Me17:0 | −0.634 | 0.587 | |
| 10Me18:0 | −0.970 | −0.017 | |
| Fungi | 18:1ω9 | 0.436 | −0.300 |
| 18:2ω6 | −0.235 | −0.706 | |
| AM Fungi | 16:1ω5 | 0.430 | 0.695 |
| Gram-negative Bacteria | 16:1ω7 | −0.459 | −0.607 |
| 17:1ω7 | −0.883 | 0.115 | |
| 17:0cy | −0.101 | −0.482 | |
| 18:1ω7 | 0.123 | −0.230 | |
| 18:1ω8 | −0.678 | 0.437 | |
| 19:0cy | −0.442 | 0.331 | |
| Gram-positive Bacteria | i14:0 | 0.623 | −0.688 |
| i15:0 | −0.427 | −0.606 | |
| a15:0 | 0.029 | −0.948 | |
| i16:0 | 0.283 | −0.868 | |
| i17:0 | −0.833 | −0.181 | |
| a17:0 | −0.436 | −0.658 | |
| Non-specific | 13:0 | −0.965 | 0.135 |
| 14:0 | −0.976 | 0.029 | |
| 15:0 | −0.959 | −0.021 | |
| 16:0 | 0.889 | 0.108 | |
| 18:0 | −0.737 | −0.403 | |
| Treatment 1 | Depth | Total Microbial Biomass | Gram+ Bacteria | Gram− Bacteria | Fungi | Actinomycetes |
|---|---|---|---|---|---|---|
| nmol g−1 | ||||||
| A | 0–2.5 | 37.3 bc | 5.02 ab | 7.34 bc | 12.7 a | 0.60 bc |
| 2.5–7.5 | 28.6 | 4.21 | 4.85 | 8.71 | 1.46 | |
| BA | 0–2.5 | 24.2 c | 3.34 b | 3.38 c | 8.33 a | 0.74 bc |
| 2.5–7.5 | 18.5 | 1.86 | 2.87 | 4.90 | 0.00 | |
| BB | 0–2.5 | 39.8 bc | 5.73 ab | 6.61 bc | 14.0 a | 0.59 bc |
| 2.5–7.5 | 22.6 | 3.41 | 3.59 | 9.21 | 0.69 | |
| C | 0–2.5 | 27.8 c | 4.50 ab | 4.44 c | 9.80 a | 0.39 bc |
| 2.5–7.5 | 28.5 | 4.58 | 5.21 | 9.03 | 0.95 | |
| CY | 0–2.5 | 68.1 a | 10.5 a | 15.1 ab | 17.7 a | 3.12 ab |
| 2.5–7.5 | 38.2 | 6.41 | 8.81 | 9.98 | 2.35 | |
| FA | 0–2.5 | 31.6 c | 5.29 ab | 6.16 bc | 10.9 a | 0.00 c |
| 2.5–7.5 | 9.03 | 1.23 | 1.52 | 3.19 | 0.00 | |
| FB | 0–2.5 | 26.5 c | 3.85 b | 5.33 bc | 9.74 a | 0.00 c |
| 2.5–7.5 | 16.3 | 2.73 | 2.43 | 5.52 | 0.00 | |
| N | 0–2.5 | 73.0 a | 11.1 a | 17.5 a | 20.2 a | 4.23 a |
| 2.5–7.5 | 38.5 | 6.33 | 8.36 | 10.2 | 2.59 | |
| Treatment 1 | Depth | Total Microbial Biomass | Gram+ Bacteria | Gram− Bacteria | Fungi | AM Fungi | Actinomycetes |
|---|---|---|---|---|---|---|---|
| nmol g−1 | |||||||
| HM+C | 0–2.5 | 531 | 92.0 | 117 | 175.0 | 46.2 | 23.9 |
| 2.5–7.5 | 368 ab | 56.0 a | 78.7ab | 118 ab | 43.3 ab | 27.7 a | |
| LM+C | 0–2.5 | 472 | 92.0 | 106 | 154 | 48.8 | 24.9 |
| 2.5–7.5 | 490 a | 71.8 a | 108a | 162 a | 64.9 a | 33.7 a | |
| HM | 0–2.5 | 499 | 87.6 | 117 | 162 | 45.2 | 22.5 |
| 2.5–7.5 | 321 b | 51.1 a | 71.2b | 94.5 b | 32.5 b | 25.4 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afkairin, A.; Stromberger, M.; Storteboom, H.; Wickham, A.; Sterle, D.G.; Davis, J.G. Soil Microbial Community Responses to Cyanobacteria versus Traditional Organic Fertilizers. Agriculture 2023, 13, 1902. https://doi.org/10.3390/agriculture13101902
Afkairin A, Stromberger M, Storteboom H, Wickham A, Sterle DG, Davis JG. Soil Microbial Community Responses to Cyanobacteria versus Traditional Organic Fertilizers. Agriculture. 2023; 13(10):1902. https://doi.org/10.3390/agriculture13101902
Chicago/Turabian StyleAfkairin, Antisar, Mary Stromberger, Heather Storteboom, Allison Wickham, David G. Sterle, and Jessica G. Davis. 2023. "Soil Microbial Community Responses to Cyanobacteria versus Traditional Organic Fertilizers" Agriculture 13, no. 10: 1902. https://doi.org/10.3390/agriculture13101902