
Different Response of Soil Microbial Carbon Use Efficiency in Compound of Feldspathic Sandstone and Sand
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sites and Experimental Design
2.2. Soil Sampling
2.3. Soil Physicochemical Properties, Microbial Biomass, and Extracellular Enzyme Activity Analysis
2.4. Statistical Analyses
3. Results
3.1. Physicochemical Properties and Stoichiometry of Soil in Different Compound Ratios
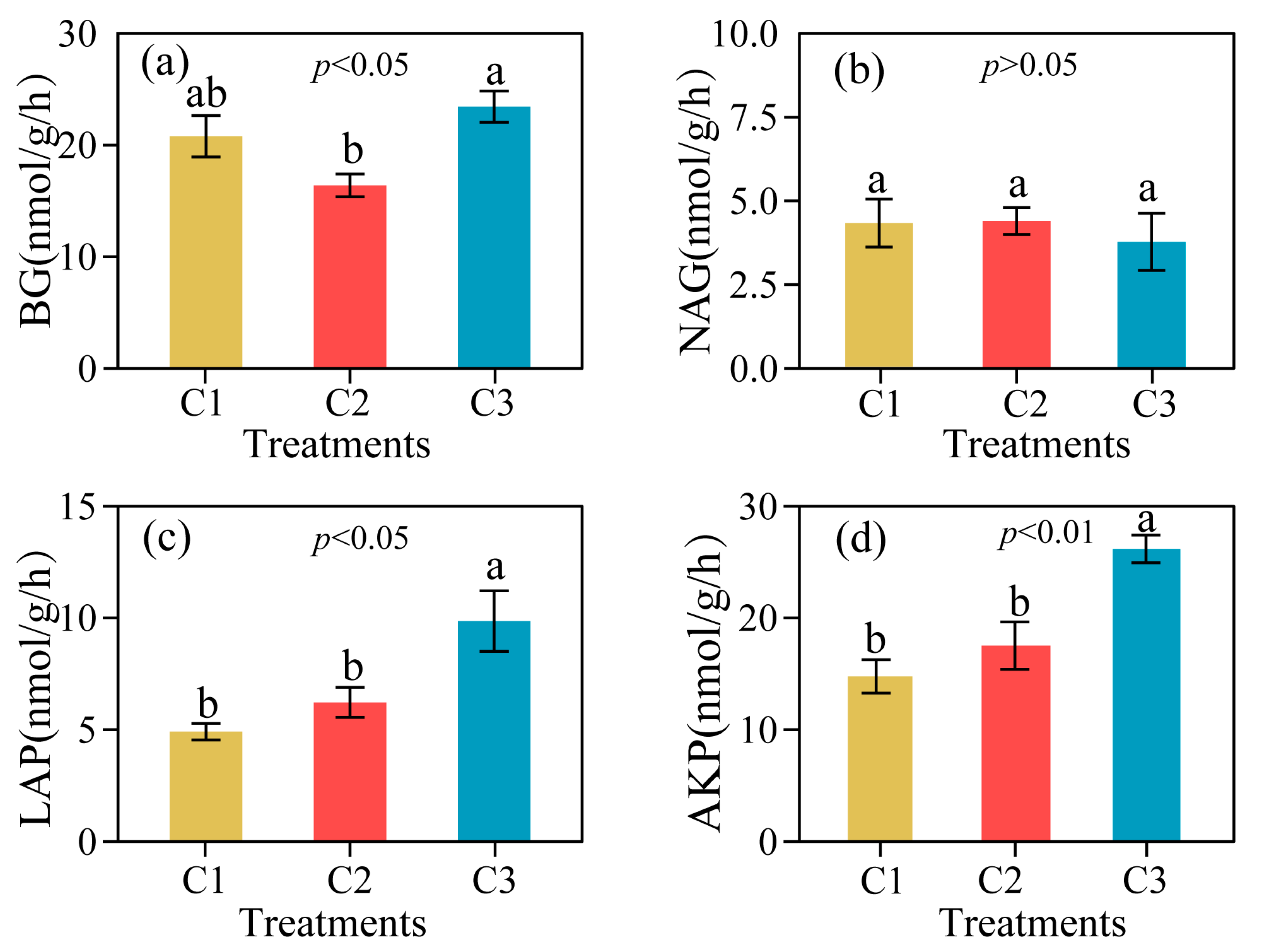
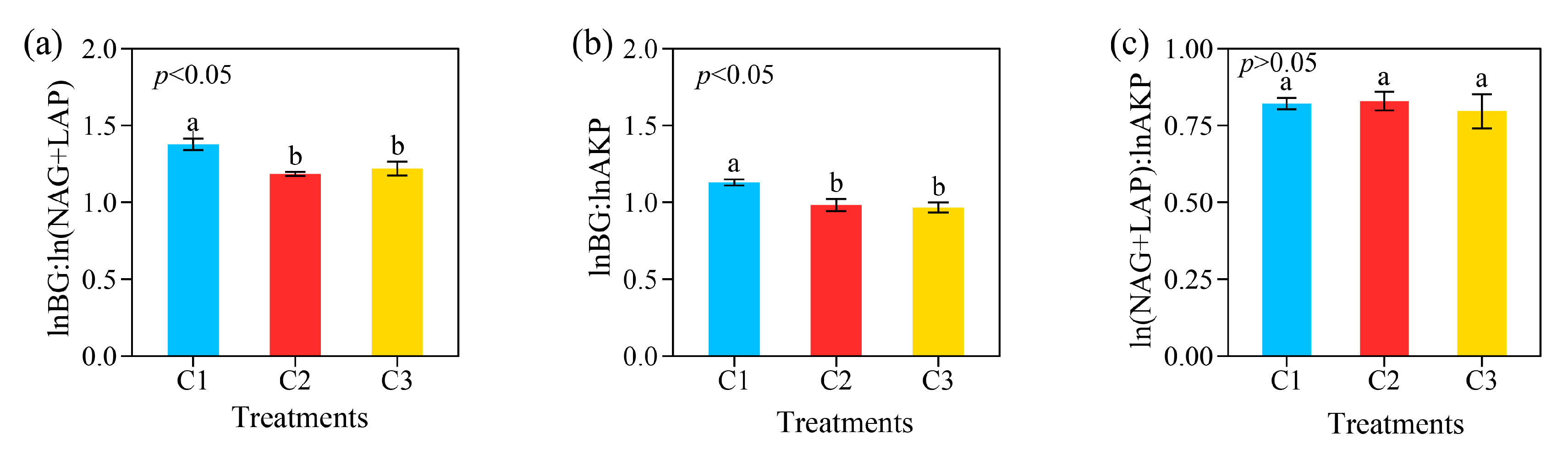
3.2. Extracellular Enzyme Activity and Its Soil Stoichiometry in Different Compound Ratios
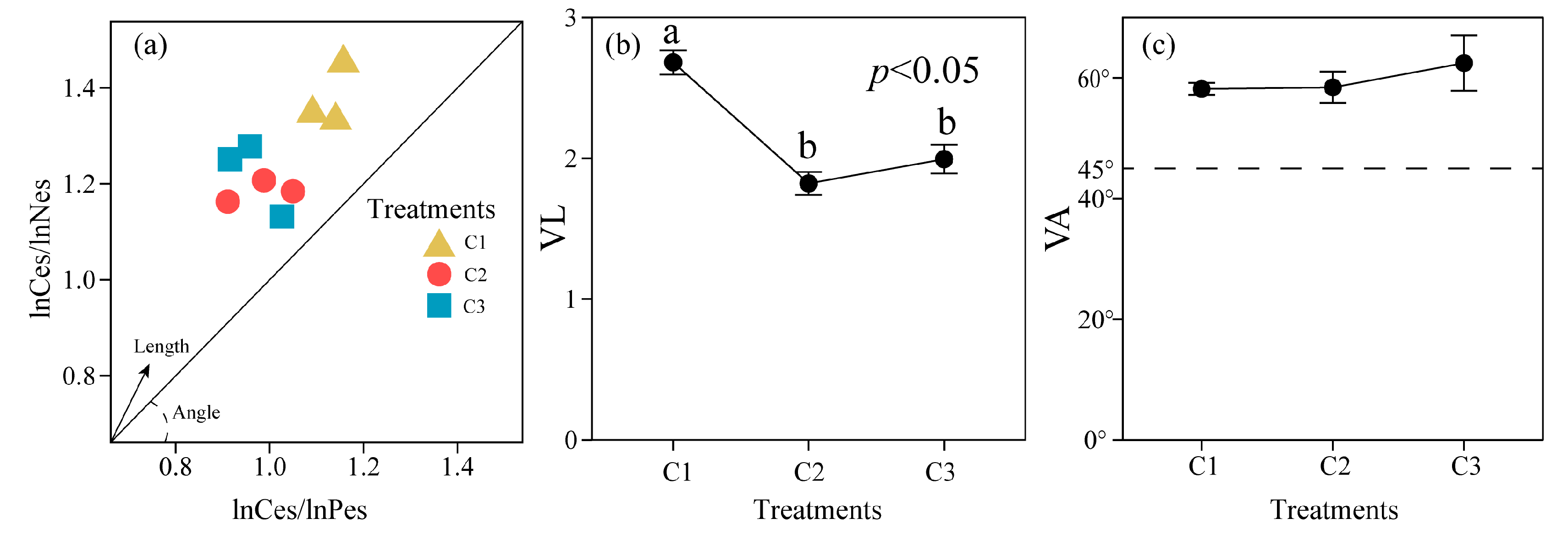
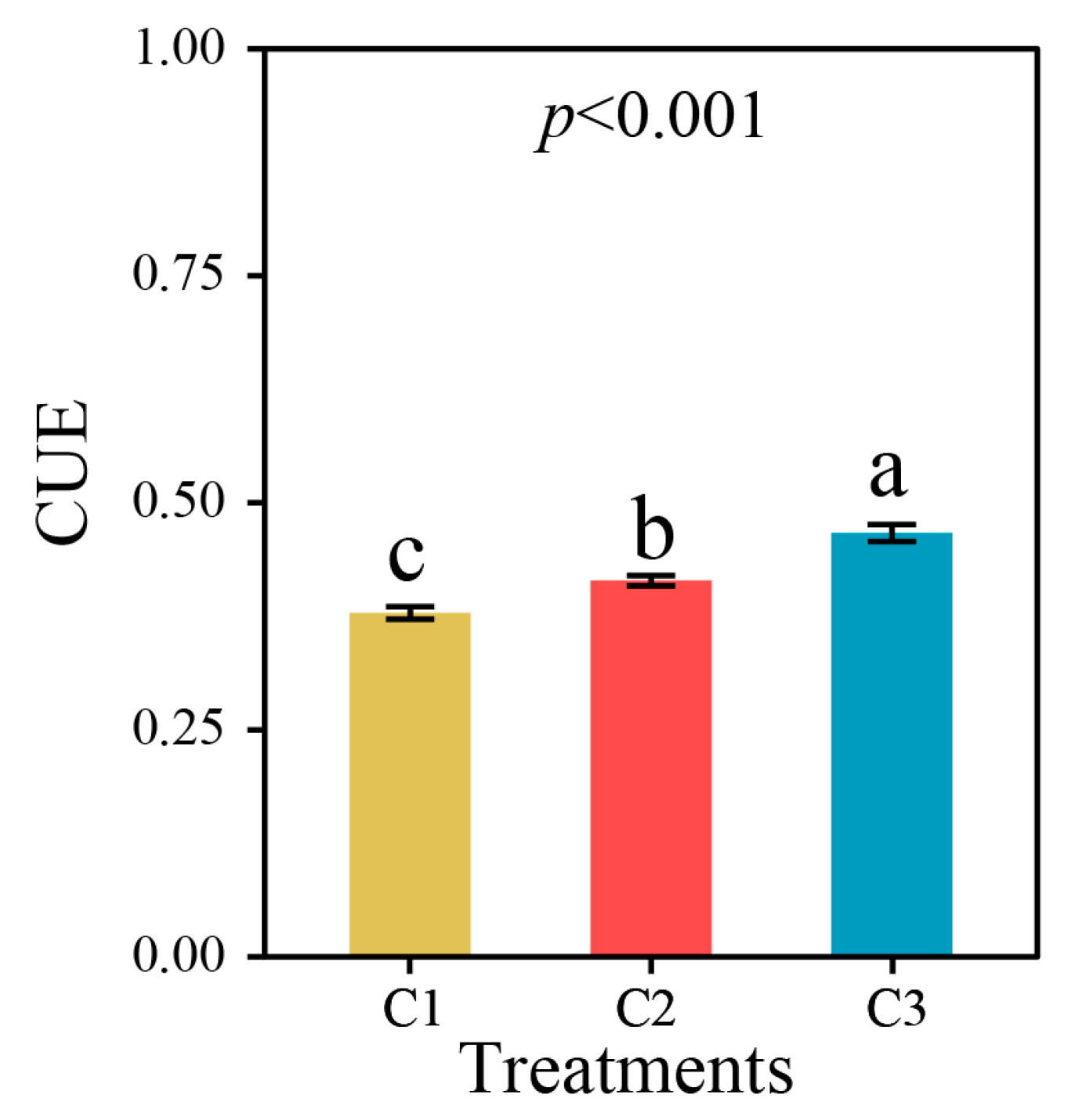
3.3. Microbial Carbon Use Efficiency of Soil in Different Compound Ratios
4. Discussion
4.1. Variation of the Stoichiometric Ratio in Different Compound Ratio Soils
4.2. Explanation of the Change in Microbe Biomass and Extracellular Enzyme Activity
4.3. Variation of Microbial Carbon Use Efficiency in Different Compound Ratio Soils
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, X.; Zhang, L.; Zhao, F.; Bai, H.; Doughty, R. Effects of Mixing Feldspathic Sandstone and Sand on Soil Microbial Biomass and Extracellular Enzyme Activities-A Case Study in Mu Us Sandy Land in China. Appl. Sci. 2019, 9, 3963. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.Q.; Sun, Z.H.; Li, G.; Wang, H.Y.; Cheng, J.; Hao, M.D. Influences of water chemical property on infiltration into mixed soil consisting of feldspathic sandstone and aeolian sandy soil. Sci. Rep. 2020, 10, 19497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Xu, M.Z.; Wang, Z.Y.; Duan, X.H.; Bi, C.F. Ecological Impacts of Seabuckthorn in the Pisha Sandstone Area. In Advances in Water Resources and Hydraulic Engineering; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1102–1107. [Google Scholar]
- Wang, N.; Xie, J.C.; Han, J.C. A Sand Control and Development Model in Sandy Land Based on Mixed Experiments of Arsenic Sandstone and Sand: A Case Study in Mu Us Sandy Land in China. Chin. Geogr. Sci. 2013, 23, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Han, J.; Wang, H.; Du, Y.; Tong, W. The improvement effects of softrock on sandy soil in Mu Us sandy land. J. Arid. Land Resour. Environ. 2015, 29, 122–127. [Google Scholar]
- Zhang, L.; Ban, J. Analyzing the Sand-fixing Effect of Feldspathic Sandstone from the Texture Characteristics. IOP Conf. Ser. Earth Environ. 2018, 108, 032039. [Google Scholar] [CrossRef]
- Li, J.; Tong, X.G.; Awasthi, M.K.; Wu, F.Y.; Ha, S.; Ma, J.Y.; Sun, X.H.; He, C. Dynamics of soil microbial biomass and enzyme activities along a chronosequence of desertified land revegetation. Ecol. Eng. 2018, 111, 22–30. [Google Scholar] [CrossRef]
- Xu, M.P.; Jian, J.N.; Wang, J.Y.; Zhang, Z.J.; Yang, G.H.; Han, X.H.; Ren, C.J. Response of root nutrient resorption strategies to rhizosphere soil microbial nutrient utilization along Robinia pseudoacacia plantation chronosequence. For. Ecol. 2021, 489, 119053. [Google Scholar] [CrossRef]
- Adingo, S.; Yu, J.R.; Liu, X.L.; Li, X.D.; Jing, S.; Xiaong, Z. Variation of soil microbial carbon use efficiency (CUE) and its Influence mechanism in the context of global environmental change: A review. PeerJ 2021, 9, e12131. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil Microorganisms Mediating Phosphorus Availability. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Manzoni, S.; Taylor, P.; Richter, A.; Porporato, A.; Agren, G.I. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol. 2012, 196, 79–91. [Google Scholar] [CrossRef]
- Siebielec, S.; Siebielec, G.; Klimkowicz-Pawlas, A.; Galazka, A.; Grzadziel, J.; Stuczynski, T. Impact of Water Stress on Microbial Community and Activity in Sandy and Loamy Soils. Agronomy 2020, 10, 1429. [Google Scholar] [CrossRef]
- Tapia-Torres, Y.; Elser, J.J.; Souza, V.; Garcia-Oliva, F. Ecoenzymatic stoichiometry at the extremes: How microbes cope in an ultra-oligotrophic desert soil. Soil Biol. Biochem. 2015, 87, 34–42. [Google Scholar] [CrossRef]
- Widdig, M.; Schleuss, P.M.; Biederman, L.A.; Borer, E.T.; Crawley, M.J.; Kirkman, K.P.; Seabloom, E.W.; Wragg, P.D.; Spohn, M. Microbial carbon use efficiency in grassland soils subjected to nitrogen and phosphorus additions. Soil Biol. Biochem. 2020, 146, 107815. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Shah, J.J.F. Ecoenzymatic Stoichiometry and Ecological Theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 313–343. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Hill, B.H.; Shah, J.J.F. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795-U117. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Shi, C. Prediction of Bacterial Community Structure and Function in Different Compound Ratio Soils. Environ. Sci. Technol. 2021, 44, 69–76. [Google Scholar]
- Neal, A.L.; Rossmann, M.; Brearley, C.; Akkari, E.; Guyomar, C.; Clark, I.M.; Allen, E.; Hirsch, P.R. Land-use influences phosphatase gene microdiversity in soils. Environ. Microbiol. 2017, 19, 2740–2753. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.Z.; Bai, L.; Wang, J.Y.; Deng, J.; Ren, C.J.; Han, X.H.; Yang, G.H.; Wang, J. Change in soil bacterial community during secondary succession depend on plant and soil characteristics. Catena 2019, 173, 246–252. [Google Scholar] [CrossRef]
- Ren, C.; Zhao, F.; Kang, D.; Yang, G.; Han, X.; Tong, X.; Feng, Y.; Ren, G. Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland. Forest Ecol. Manag. 2016, 376, 59–66. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem. 1982, 14, 319–329. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Turner, B.L.; Talbot, J.M.; Waring, B.G.; Powers, J.S.; Kuske, C.R.; Moorhead, D.L.; Shah, J.J.F. Stoichiometry of microbial carbon use efficiency in soils. Ecol. Monogr. 2016, 86, 172–189. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Li, W.; Lu, X.; Gu, Y.; Wu, S.; Shen, Z.; Han, X.; Yang, G.; Ren, C. Adaptive pathways of soil microorganisms to stoichiometric imbalances regulate microbial respiration following afforestation in the Loess Plateau, China. Soil Biol. Biochem. 2020, 151, 108048. [Google Scholar] [CrossRef]
- Guo, Z.; Han, J.C.; Li, J. Response of organic carbon mineralization and bacterial communities to soft rock additions in sandy soils. PeerJ 2020, 8, e8948. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Li, H.; Zhu, J.; Wei, Y.; Zhang, F.; Yang, Y.; Li, Y. The asymptotic response of soil water holding capacity along restorationduration of artificial grasslands from degraded alpine meadowsin the Three River Sources, Qinghai–Tibetan Plateau, China. Ecol. Res. 2018, 33, 1001–1010. [Google Scholar] [CrossRef]
- Xu, X.; Thornton, P.E.; Post, W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- Brucker, E.; Kernchen, S.; Spohn, M. Release of phosphorus and silicon from minerals by soil microorganisms depends on the availability of organic carbon. Soil Biol. Biochem. 2020, 143, 107737. [Google Scholar] [CrossRef]
- Sariyildiz, T.; Anderson, J.M. Interactions between litter quality, decomposition and soil fertility: A laboratory study. Soil Biol. Biochem. 2003, 35, 391–399. [Google Scholar] [CrossRef]
- Sun, F.; Song, C.; Wang, M.; Lai, D.Y.F.; Tariq, A.; Zeng, F.; Zhong, Q.; Wang, F.; Li, Z.; Peng, C. Long-term increase in rainfall decreases soil organic phosphorus decomposition in tropical forests. Soil Biol. Biochem. 2020, 151, 108056. [Google Scholar] [CrossRef]
- Jiang, Y.; Guo, X. Stoichiometric patterns of soil carbon, nitrogen, and phosphorus in farmland of the Poyang Lake region in Southern China. J. Soils Sediments 2019, 19, 3476–3488. [Google Scholar] [CrossRef]
- Quan, X.U.; Wenyi, R.U.I.; Jialong, L.I.U.; Zhi, L.I.U.; Ling, Y.; Yujing, Y.I.N.; Weiian, Z. Spatial Variation of Coupling Characteristics of Soil Carbon and Nitrogen in Farmland of China. J. Ecol. Rural. Environ. 2006, 22, 57–60. [Google Scholar]
- Jiang, X.; Wright, A.L.; Wang, X.; Liang, F. Tillage-induced changes in fungal and bacterial biomass associated with soil aggregates: A long-term field study in a subtropical rice soil in China. Appl. Soil Ecol. 2011, 48, 168–173. [Google Scholar] [CrossRef]
- Wang, R.Z.; Lu, L.Y.; Creamer, C.A.; Dijkstra, F.A.; Liu, H.Y.; Feng, X.; Yu, G.Q.; Han, X.G.; Jiang, Y. Alteration of soil carbon and nitrogen pools and enzyme activities as affected by increased soil coarseness. Biogeosciences 2017, 14, 2155–2166. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.K.; Chen, L.C.; Yang, Q.P.; Sun, T.; Li, C.M. Different effects of single versus repeated additions of glucose on the soil organic carbon turnover in a temperate forest receiving long-term N addition. Geoderma 2019, 341, 59–67. [Google Scholar] [CrossRef]
- Gallardo, A.S.W. Carbon and nitrogen limitati on of soil microbial biomass in desert ecosystems. Biogeochemistry 1992, 18, 1–17. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Responses of extracellular enzymes to simple and complex nutrient inputs. Soil Biol. Biochem. 2005, 37, 937–944. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Niu, D.; Gherardi, L.A.; Liu, Y.; Wang, Y.; Elser, J.J.; Fu, H. Linkages of stoichiometric imbalances to soil microbial respiration with increasing nitrogen addition: Evidence from a long-term grassland experiment. Soil Biol. Biochem. 2019, 138, 107580. [Google Scholar] [CrossRef]
- Islam, M.R.; Singh, B.; Dijkstra, F.A. Microbial carbon use efficiency of glucose varies with soil clay content: A meta-analysis. Appl. Soil. Ecol. 2023, 181, 104636. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C1 | C2 | C3 | F(2,6) | p | |
|---|---|---|---|---|---|
| pH | 8.9 ± 0.04 a | 8.95 ± 0.03 a | 8.97 ± 0.01 a | 1.22 | - |
| SOC (g/kg) | 1.52 ± 0.07 a | 1.53 ± 0.15 a | 1.95 ± 0.18 a | 3.03 | - |
| TN (g/kg) | 0.15 ± 0.01 b | 0.16 ± 0.01 b | 0.21 ± 0.01 a | 7.01 | <0.05 |
| TP (g/kg) | 0.25 ± 0.01 b | 0.3 ± 0.02 a | 0.31 ± 0 a | 9.42 | <0.05 |
| C:N | 10.15 ± 0.14 a | 9.66 ± 0.58 a | 9.41 ± 0.52 a | 0.68 | - |
| C:P | 6.14 ± 0.44 a | 5.19 ± 0.74 a | 6.23 ± 0.53 a | 0.96 | - |
| N:P | 0.61 ± 0.05 a | 0.54 ± 0.07 a | 0.66 ± 0.04 a | 1.35 | - |
| MBC (mg/kg) | 87.82 ± 2.67 a | 86.25 ± 3.39 a | 93.48 ± 6.75 a | 0.68 | - |
| MBN (mg/kg) | 7.05 ± 0.32 a | 5.48 ± 0.07 b | 5.42 ± 0.28 b | 13.76 | <0.01 |
| MBP (mg/kg) | 6.21 ± 0.8 b | 15.33 ± 1.73 a | 3.92 ± 0.36 b | 28.98 | <0.001 |
| MBC:MBN | 12.51 ± 0.69 b | 15.74 ± 0.46 a | 17.23 ± 0.62 a | 16.33 | <0.01 |
| MBC:MBP | 14.81 ± 2.55 b | 5.74 ± 0.54 c | 24.1 ± 2.09 a | 22.6 | <0.01 |
| MBN:MBP | 1.19 ± 0.22 a | 0.36 ± 0.04 b | 1.41 ± 0.14 a | 13.33 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wang, J.; Chen, L.; Zhou, S.; Zhang, L.; Zhao, F. Different Response of Soil Microbial Carbon Use Efficiency in Compound of Feldspathic Sandstone and Sand. Agriculture 2023, 13, 58. https://doi.org/10.3390/agriculture13010058
Zhang Y, Wang J, Chen L, Zhou S, Zhang L, Zhao F. Different Response of Soil Microbial Carbon Use Efficiency in Compound of Feldspathic Sandstone and Sand. Agriculture. 2023; 13(1):58. https://doi.org/10.3390/agriculture13010058
Chicago/Turabian StyleZhang, Yao, Junqi Wang, Lan Chen, Sha Zhou, Lu Zhang, and Fazhu Zhao. 2023. "Different Response of Soil Microbial Carbon Use Efficiency in Compound of Feldspathic Sandstone and Sand" Agriculture 13, no. 1: 58. https://doi.org/10.3390/agriculture13010058