
Weather Conditions and Biostimulants Influence Nitrogen Acquisition from Different Sources by Soybean Plants


Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Climate of the Study Area
2.2. Description of the Experiment
2.3. Chemical Analyses
- (a)
- the percentage of nitrogen derived from the atmosphere:
- %NDFA—% of nitrogen fixed from the atmosphere,
- at % 15N wzbog. fx—15N isotope excess in soybean,
- at % 15N wzbog. nfx—15N isotope excess in the control plant—maize;
- (b)
- the percentage of nitrogen derived from fertiliser:
- %NDFF—% of nitrogen fixed from fertiliser,
- at % 15N wzbog. fx—15N isotope excess in soybean,
- at % 15N wzbog. nawozu—15N isotope excess in the fertiliser;
- (c)
- the percentage of nitrogen taken up from soil:
- %NDFS—% of nitrogen taken up from the soil,
- %NDFA—% of nitrogen fixed from the atmosphere;
- %NDFF—% of nitrogen acquired from the fertiliser.
2.4. Analysis of Meteorological Conditions
- P—monthly atmospheric precipitation sum,
- Σt—monthly sum of air temperature.
2.5. Statistical Analyses
3. Results and Discussion
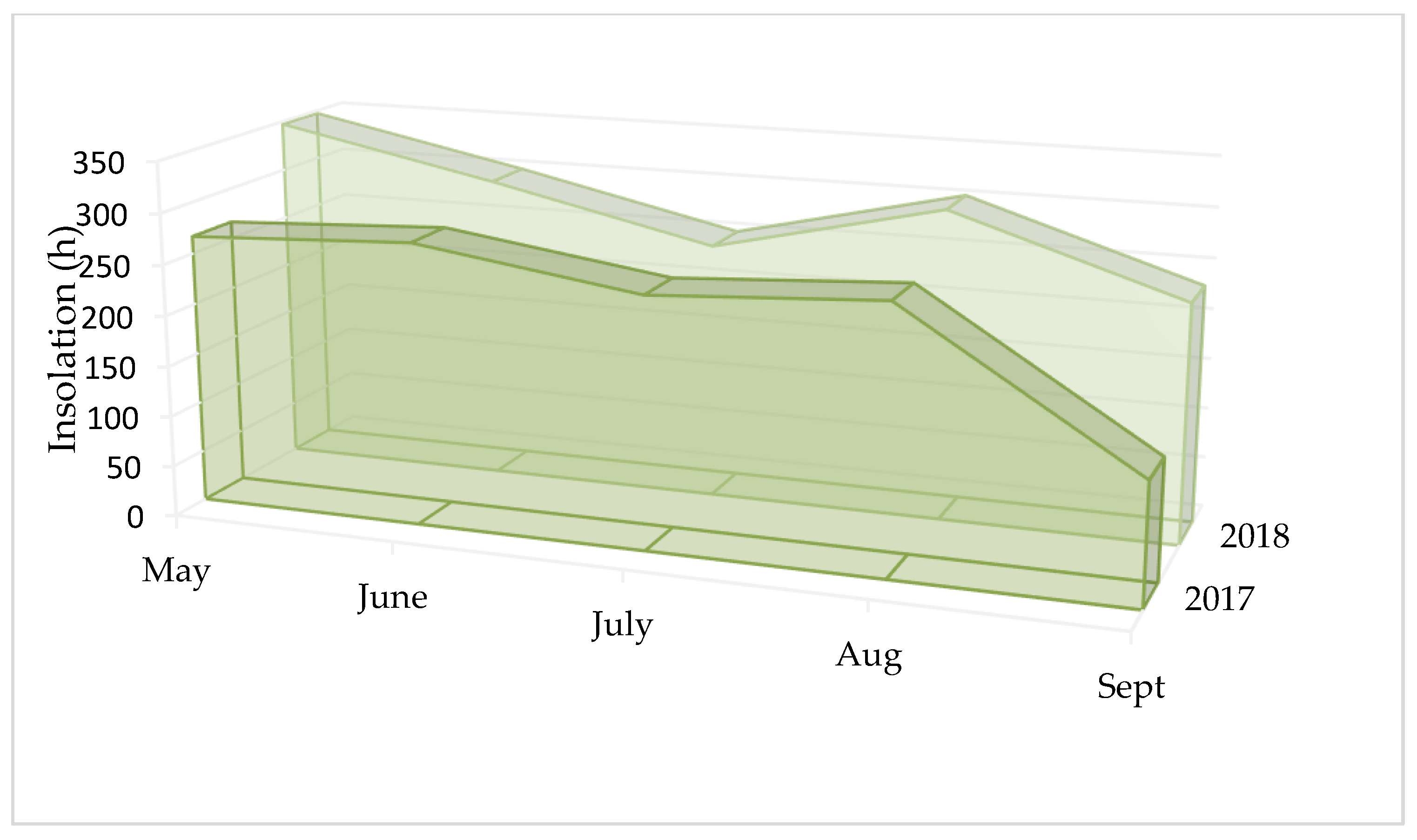
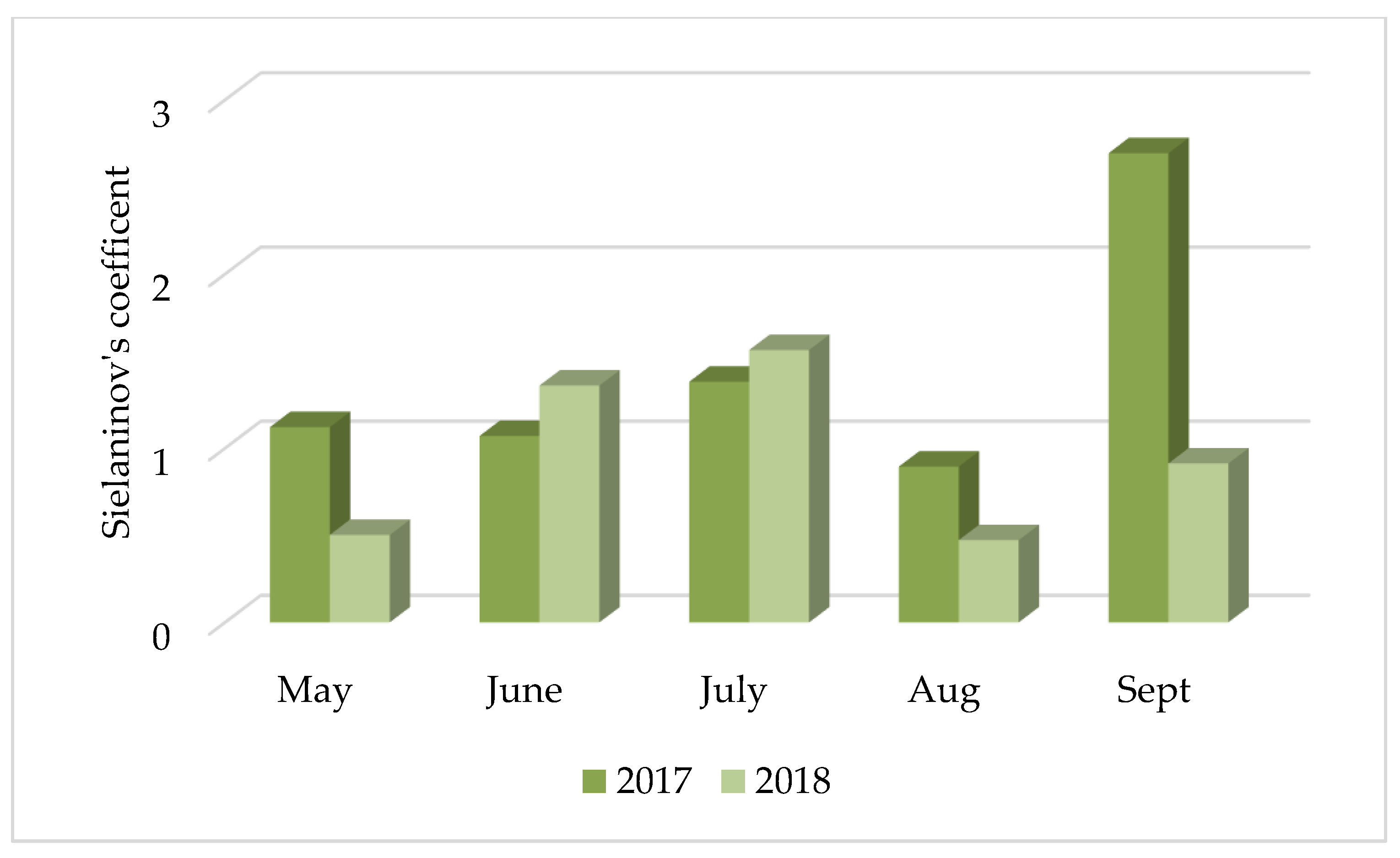
3.1. Analysis of Meteorological Conditions
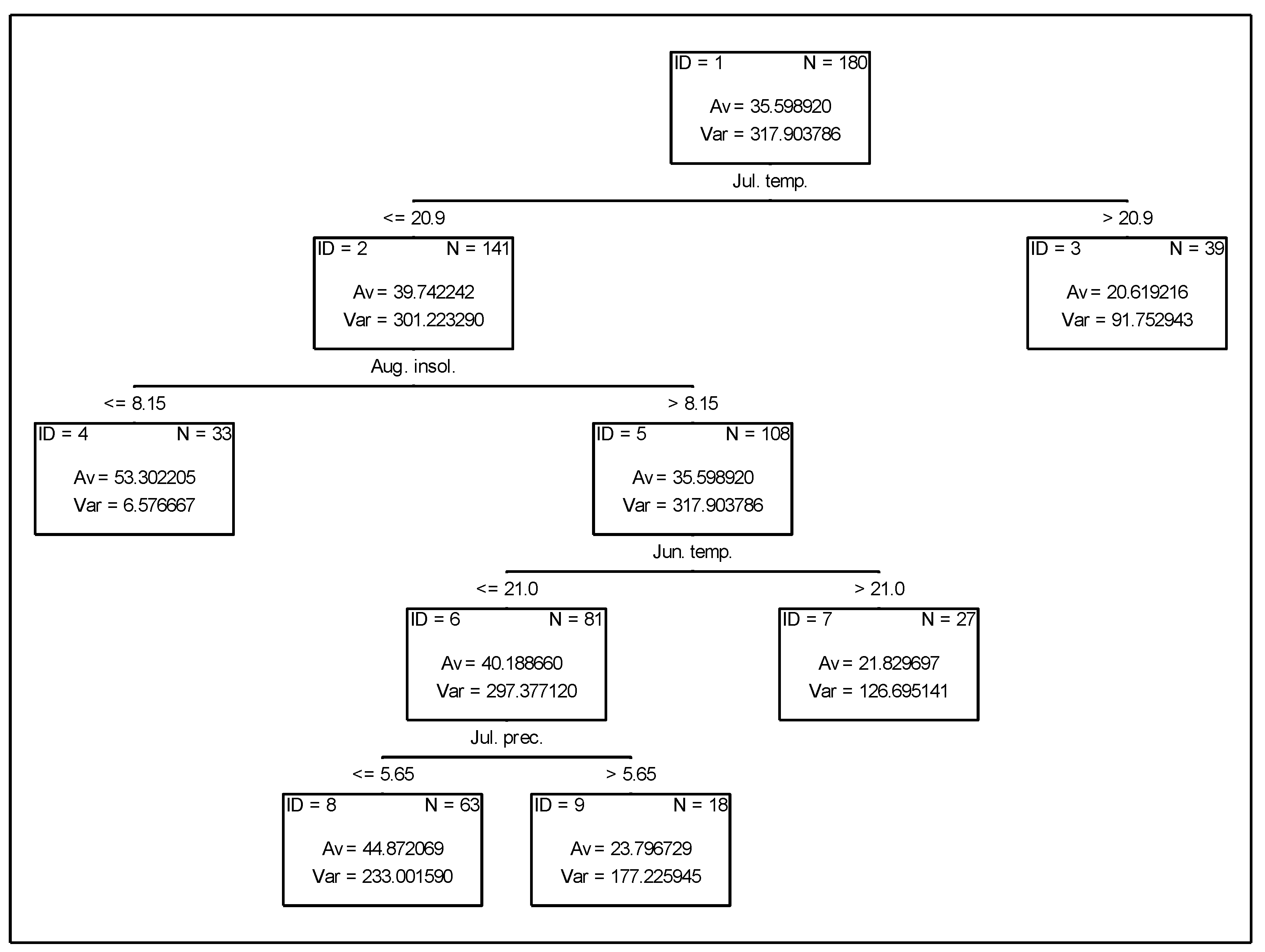
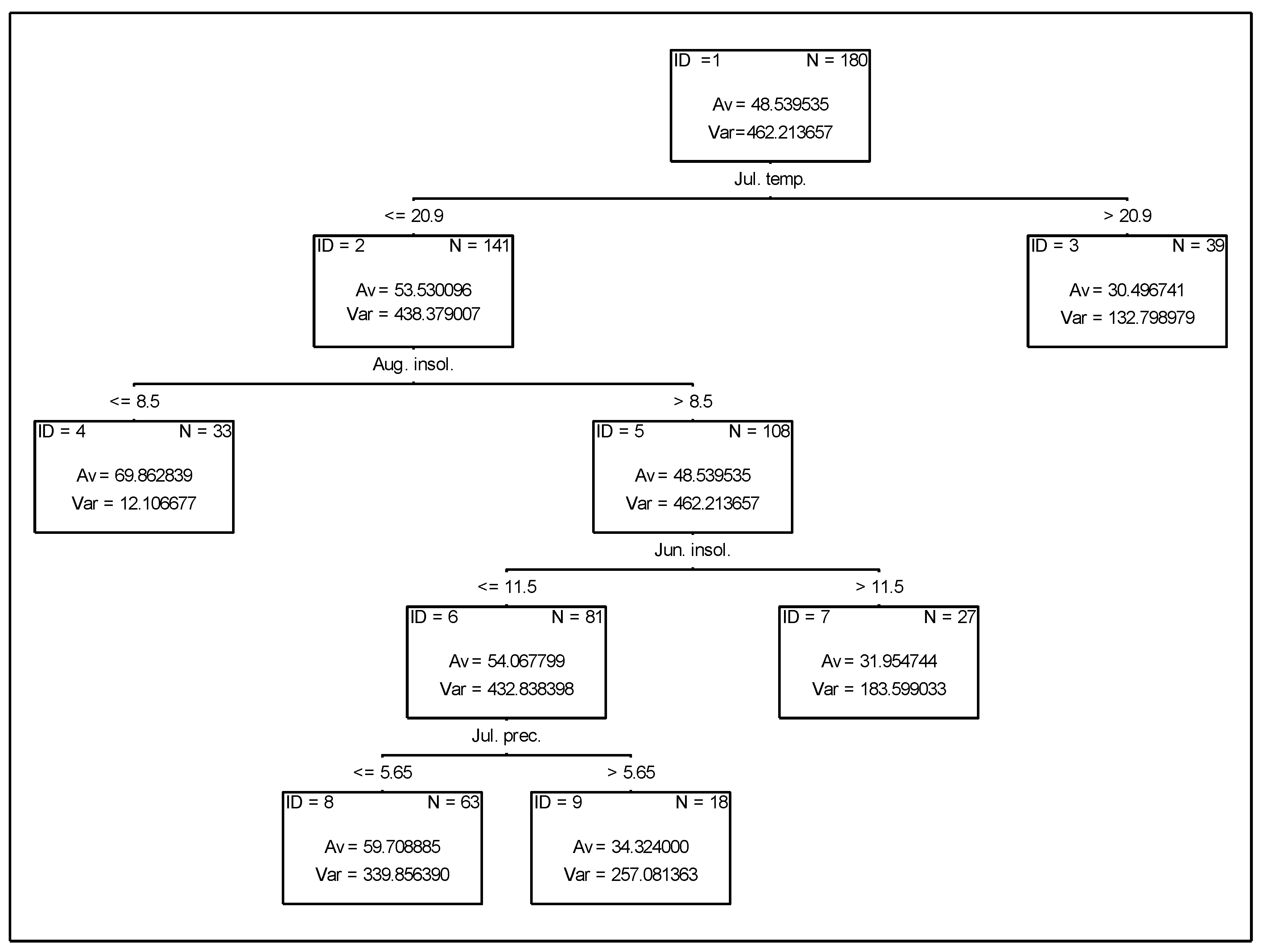
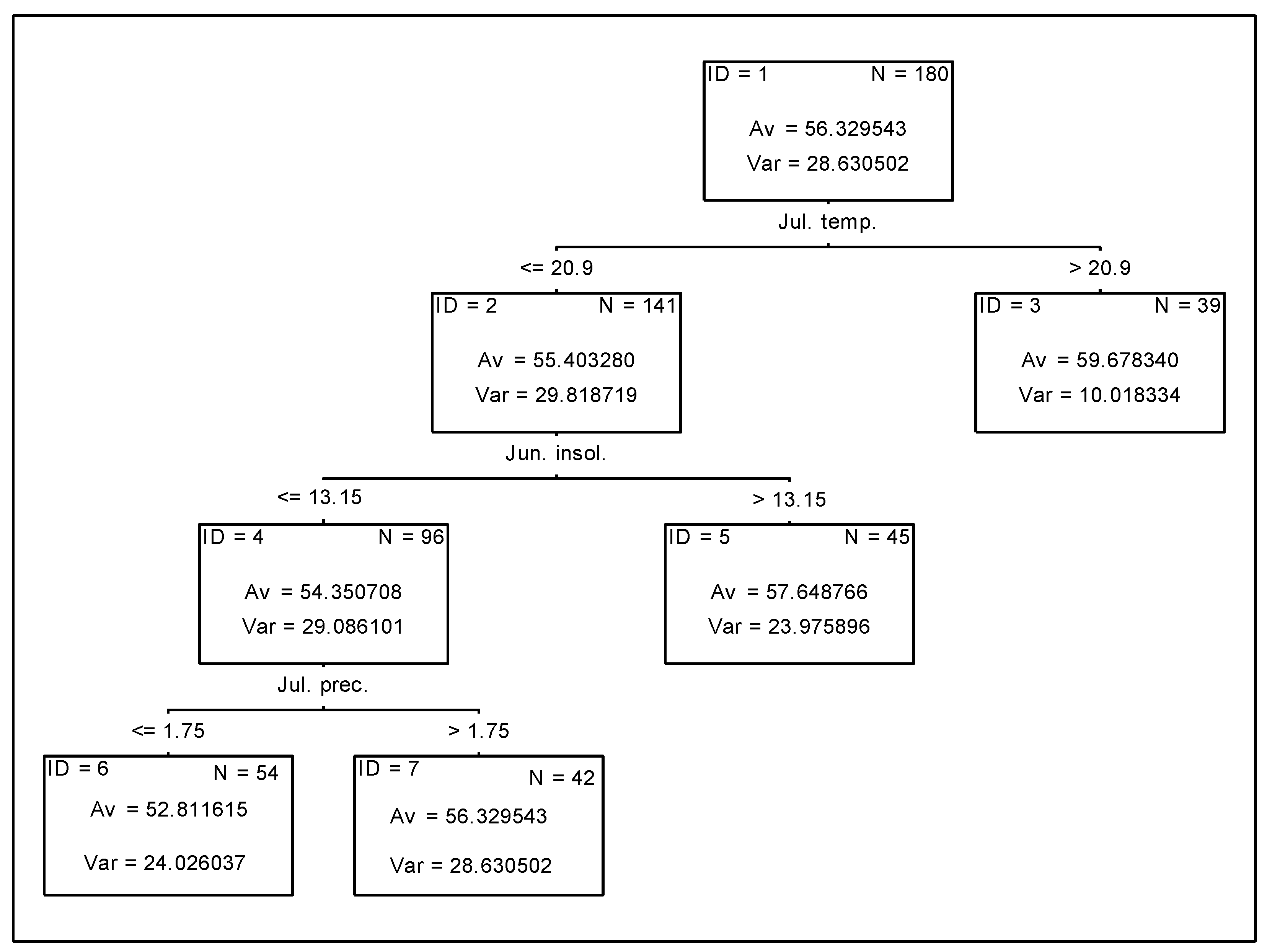
3.2. Analysis of the Effect of Meteorological Conditions on Nitrogen Fixation from the Atmosphere
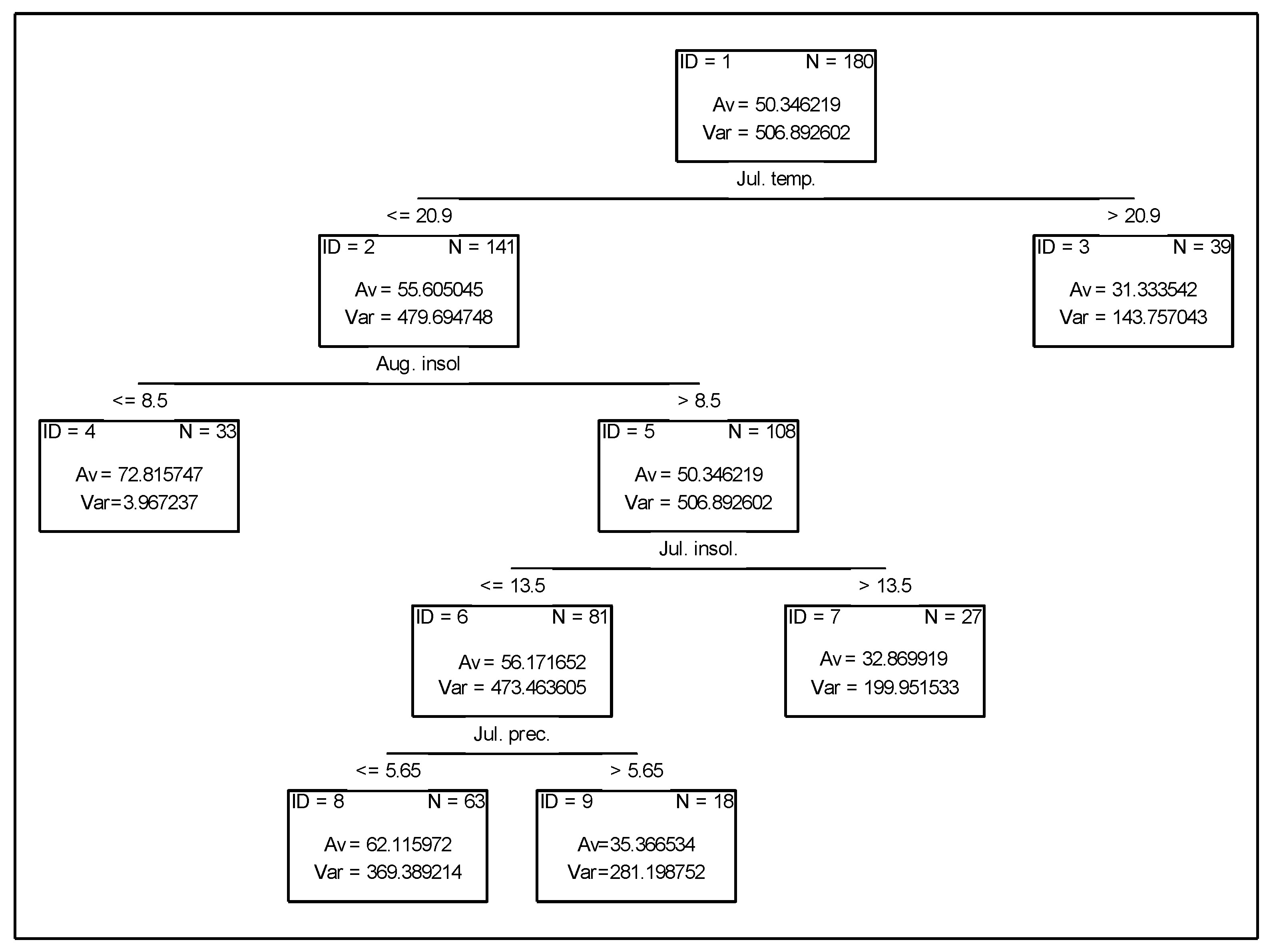
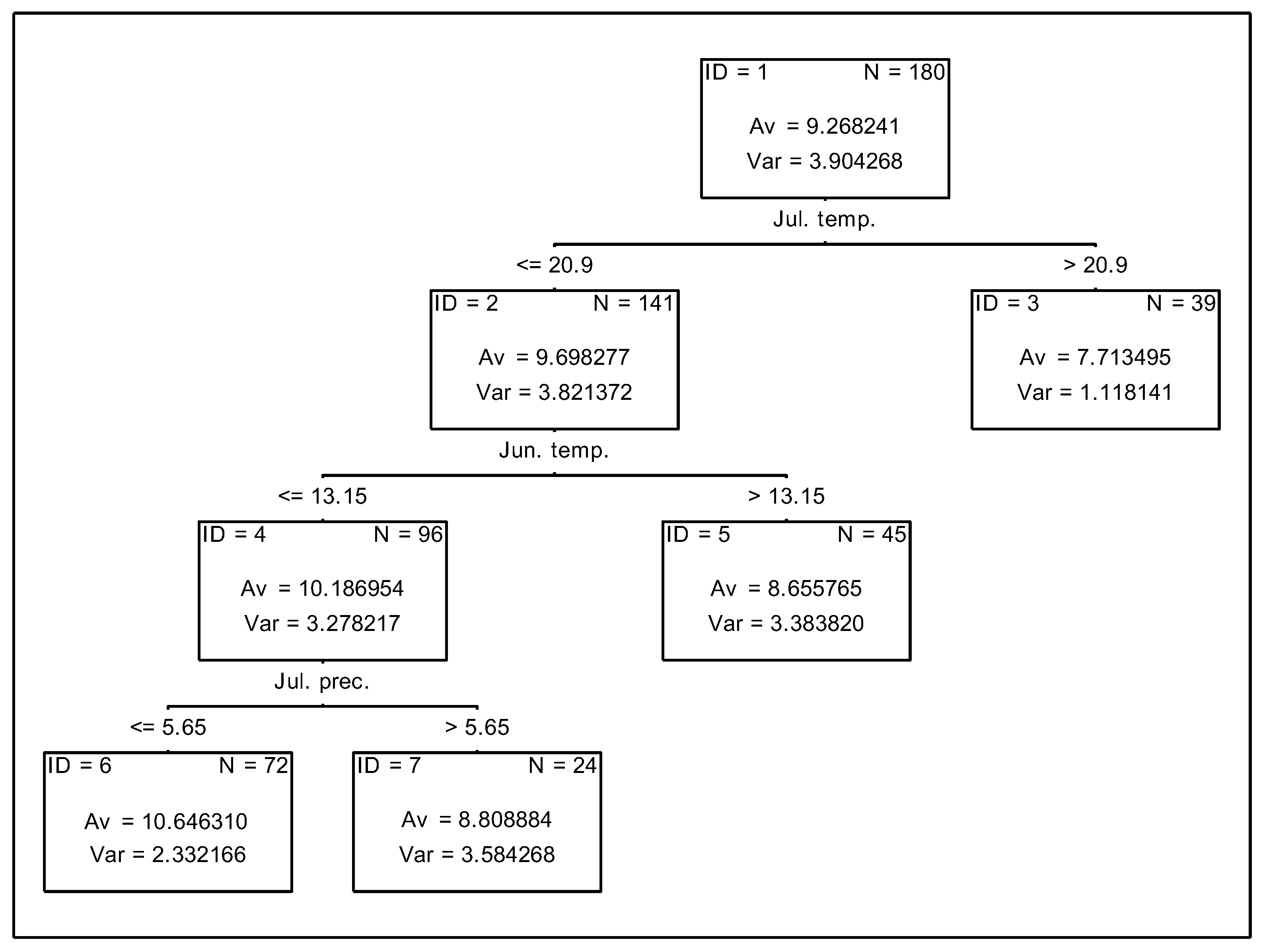
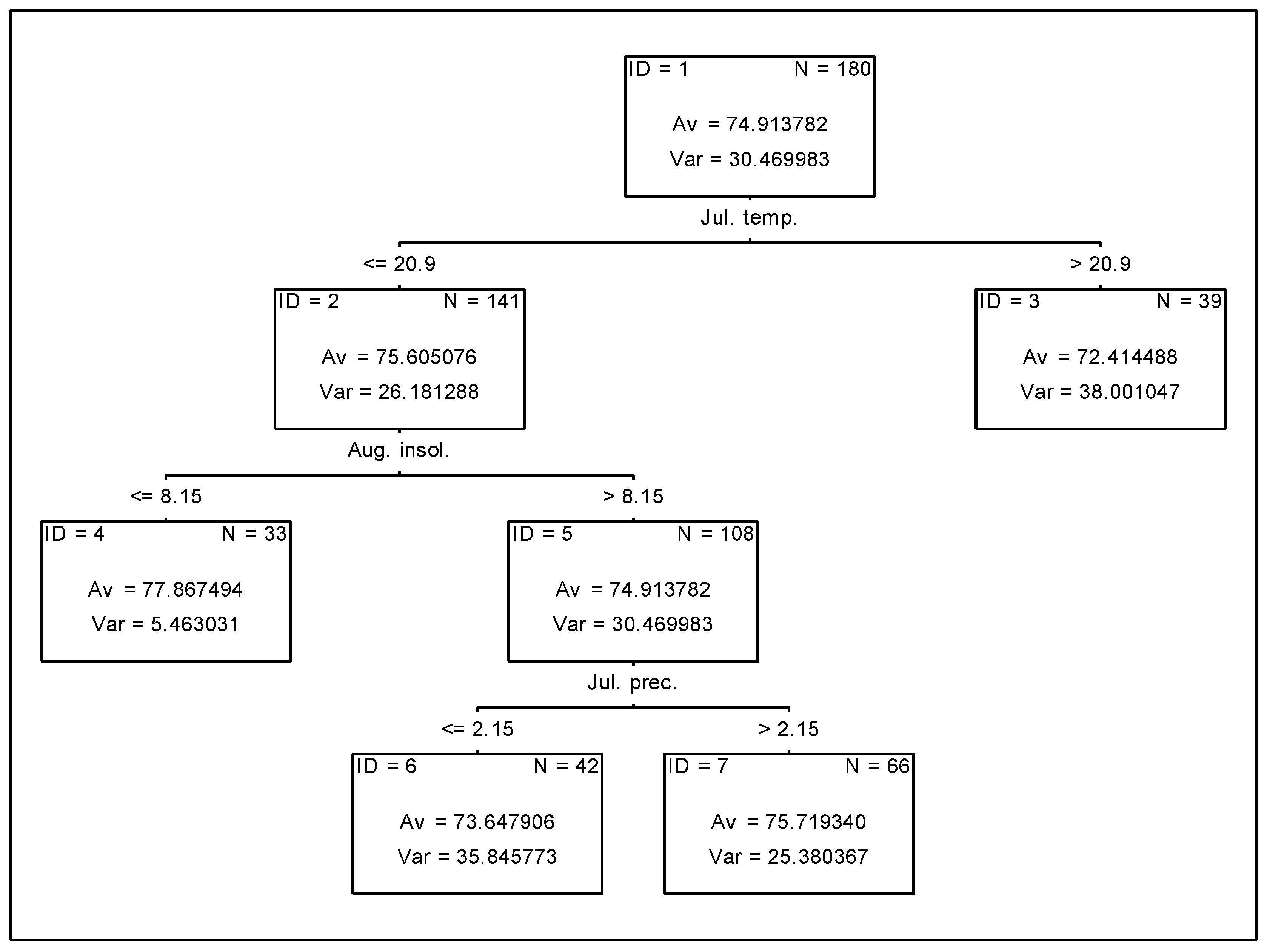
3.3. Analysis of the Effect of Meteorologial Conditions on Nitrogen Uptake from Fertiliser
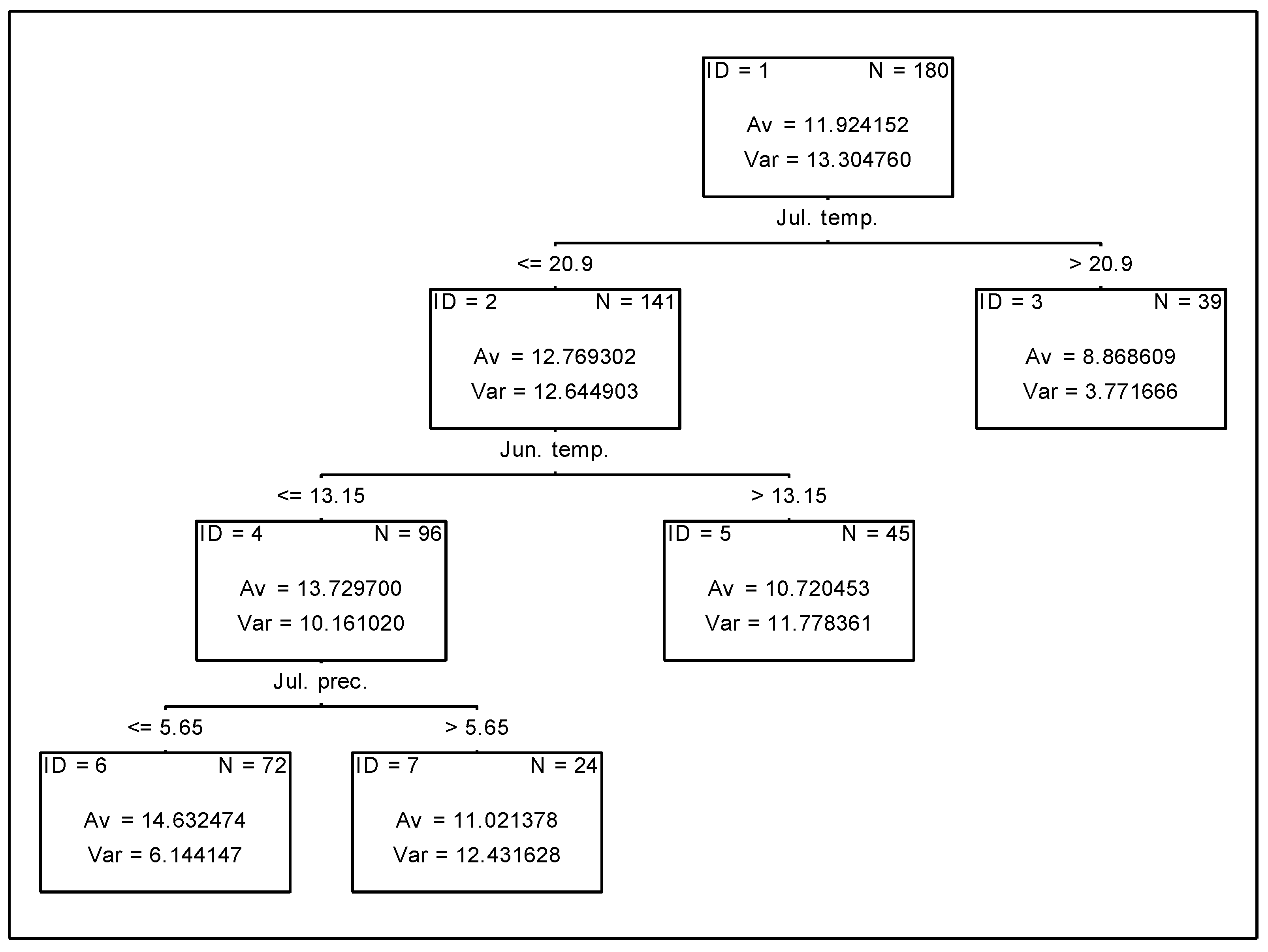
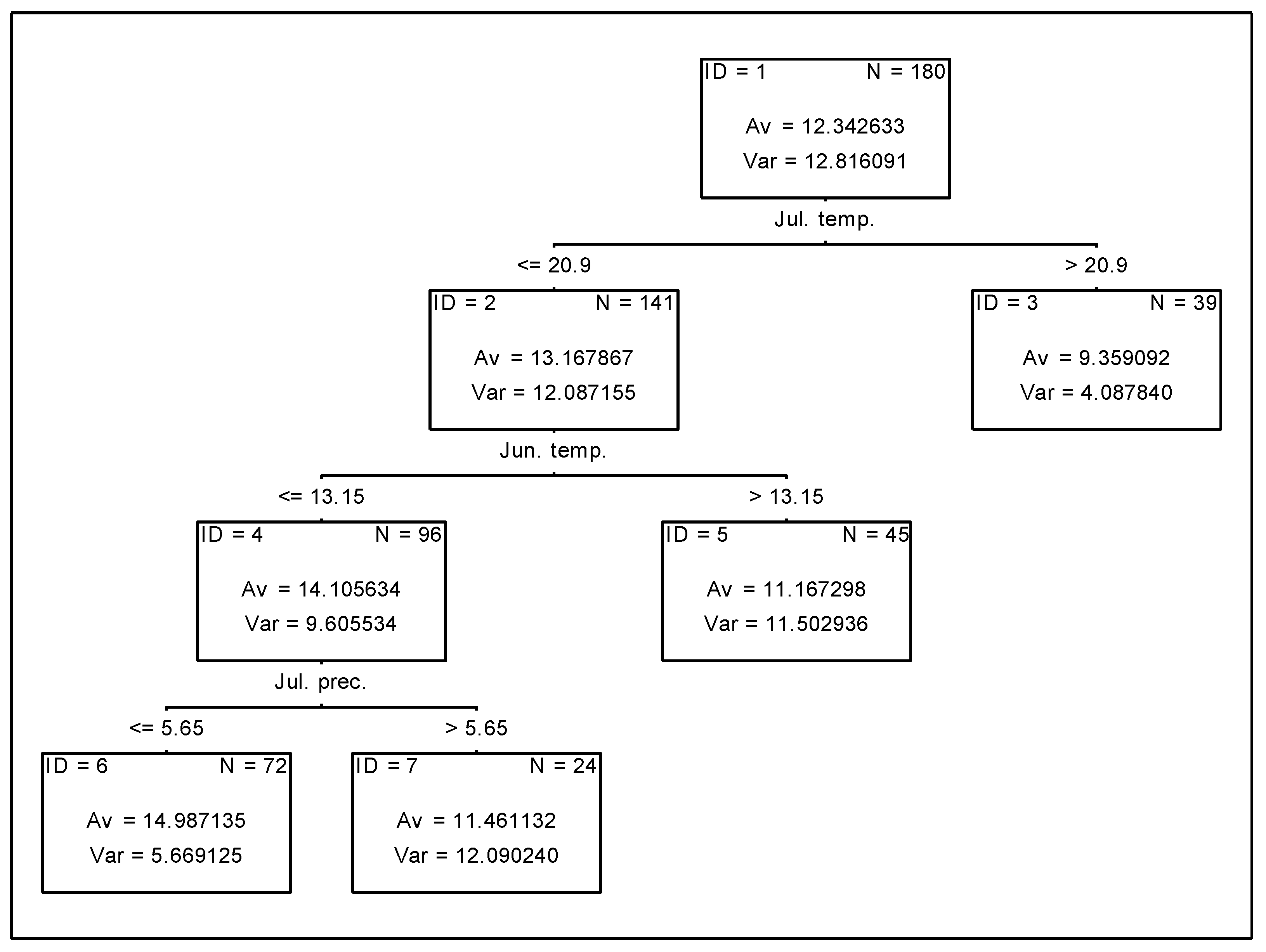
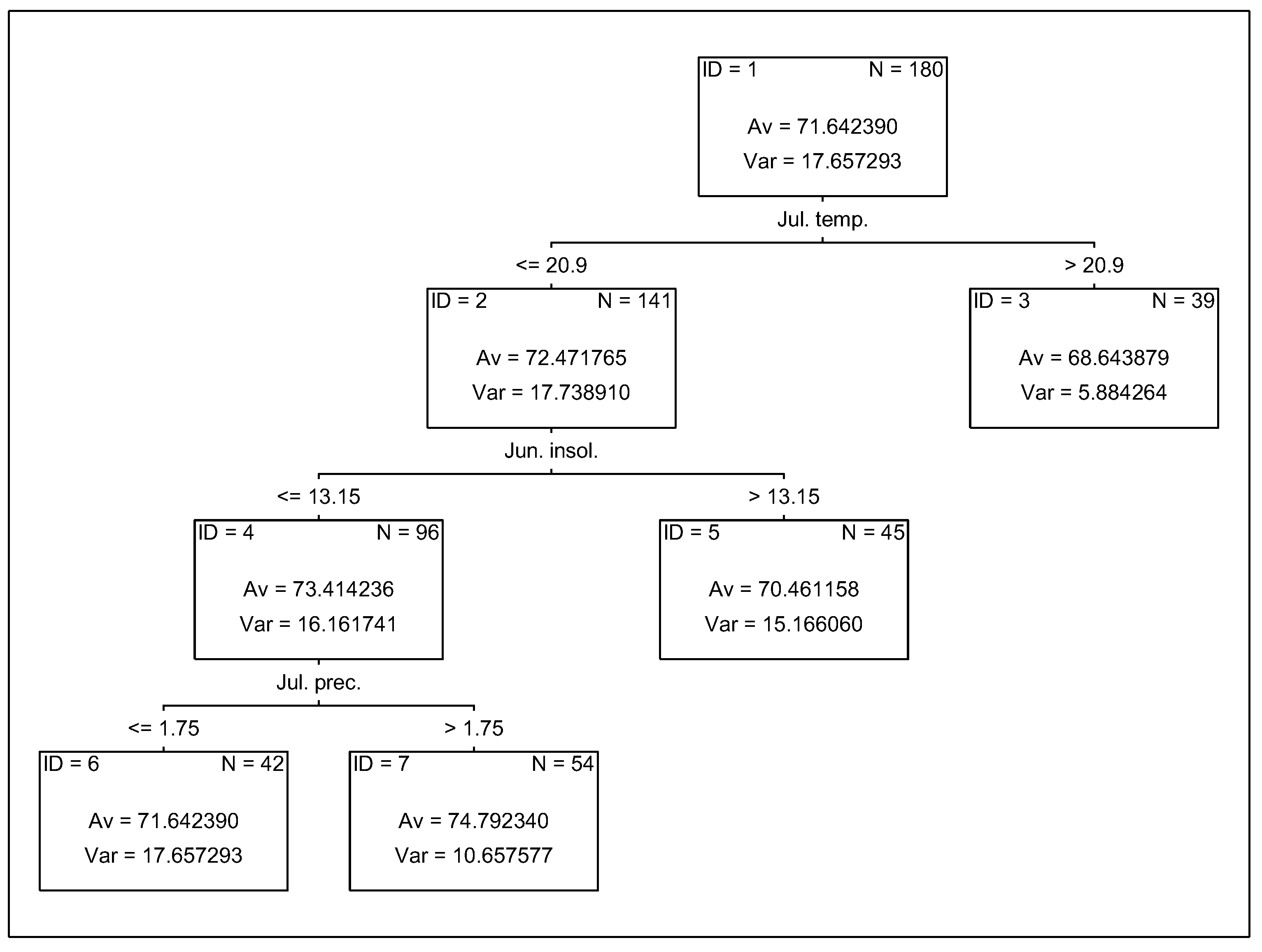
3.4. Analysis of the Effect of Meteorological Conditions on Nitrogen Uptake from Soil
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flechard, C.R.; Ambus, P.; Skiba, U.; Rees, R.M.; Hensen, A.; van Amstel, A.; van den Pol-van Dasselaar, A.; Soussana, J.-F.; Jones, M.; Clifton- Brown, J.; et al. Effects of climate and management intensity on nitrous oxide emissions in grassland systems across Europe. Agric. Ecosyst. Environ. 2007, 121, 135–152. [Google Scholar] [CrossRef]
- Stout, W.L.; Fales, S.L.; Muller, L.D.; Schnabel, R.R.; Weaver, S.R. Water quality implications of nitrate leaching from intensively grazed pasture swards in the northeast US. Agric. Ecosyst. Environ. 2000, 77, 203–210. [Google Scholar] [CrossRef]
- Trindade, H.; Coutinho, J.; Jarvis, S.; Moreira, N. Nitrogen mineralization in sandy loam soils under an intensive double-cropping forage system with dairy-cattle slurry applications. Eur. J. Agron. 2001, 15, 281–293. [Google Scholar] [CrossRef]
- Griesmann, M.; Chang, Y.; Liu, X.; Song, Y.; Haberer, G.; Crook, M.B.; Billault-Penneteau, B.; Lauressergues, D.; Keller, J.; Imanishi, L.; et al. Phylogenomics reveals multiple losses of nitrogen-fixing root nodule symbiosis. Science 2018, 361, eaat1743. [Google Scholar] [CrossRef] [Green Version]
- Kapusta, F. The role of leguminous plants in Polish agriculture. Zagadnienia Doradz. Rol. 2017, 1, 68–78. (In Polish) [Google Scholar]
- Batterman, S.A.; Hedin, L.O.; Van Breugel, M.; Ransijn, J.; Craven, D.J.; Hall, J.S. Key role of symbiotic dinitrogen fixation in tropical forest secondary succession. Nature 2013, 502, 224–227. [Google Scholar] [CrossRef]
- Batterman, S.A.; Wurzburger, N.; Hedin, L.O. Nitrogen and phosphorus interact to control tropical symbiotic N2 fixation: A test in Inga punctata. J. Ecol. 2013, 101, 1400–1408. [Google Scholar] [CrossRef]
- McCallum, M.H.; Peoples, M.B.; Connor, D.J. Contributions of nitrogen by field pea (Pisum sativum L.) in a continuous cropping sequence compared with a lucerne (Medicago sativa L.)-based pasture ley in the Victorian Wimmera. Aust. J. Agric. Res. 1999, 51, 13–22. [Google Scholar] [CrossRef]
- Fatima, Z.; Zia, M.; Chaudhary, M.F. Interactive effect of Rhizobium strains and P on soybean yield, nitrogen fixation and soil fertility. Pak. J. Bot. 2007, 39, 255. [Google Scholar]
- Carlsson, G.; Huss-Danell, K. How to quantify biological nitrogen fixation in forage legumes in the field. In Biological Nitrogen Fixation: Towards Poverty Alleviation through Sustainable Agriculture; Dakora, F.D., Chimphango, S.B.M., Valentine, A.J., Elmerich, C., Newton, W.E., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 47–48. [Google Scholar]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Ann. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef] [Green Version]
- Lewandowska, S. Influence of environmental conditions on yields and chemical composition of soybean grown in Opolskie Voivodeship. In Monographs CCXVIII 2019; Wrocław University of Environmental and Life Sciences Publishing House: Wrocław, Poland, 2019. [Google Scholar]
- Mourtzinis, S.; Specht, J.E.; Conley, S.P. Defining optimal soybean sowing dates across the US. Sci. Rep. 2019, 9, 2800. [Google Scholar] [CrossRef] [Green Version]
- Setiyono, T.D.; Weiss, A.; Specht, J.; Bastidas, A.M.; Cassman, K.G.; Dobermann, A. Understanding and modeling the effect of temperature and daylength on soybean phenology under high-yield conditions. Field Crop. Res. 2007, 100, 257–271. [Google Scholar] [CrossRef]
- Nico, M.; Miralles, D.J.; Kantolic, A.G. Natural post-flowering photoperiod and photoperiod sensitivity: Roles in yield determining processes in soybean. Field Crop. Res. 2019, 231, 141–152. [Google Scholar] [CrossRef]
- Sediyama, T. Tecnologias de Produção e Usos da Soja; Mecenas Publishing: Londrina, Brazil, 2009; ISBN 9788589687089. [Google Scholar]
- Warzecha, E. Development and characteristics of selected soybean traits under the climatic conditions of Poland. Acta Agrobot. 1983, 36, 191–202. (In Polish) [Google Scholar] [CrossRef] [Green Version]
- Camara, G.M.S.; Sediyama, T.; Dourado-Neto, D.; Bernardes, M.S. Influence of photoperiod and air temperature on the growth, flowering and maturation of the soybean (Glycine max. L. Merrill). Sci. Agric. 1997, 54, 149–154. [Google Scholar] [CrossRef]
- Alsajri, F.A.; Wijewardana, C.; Irby, J.T.; Bellaloui, N.; Krutz, L.J.; Golden, B.; Gao, W.; Reddy, K.R. Developing functional relationships between temperature and soybean yield and seed quality. Agron. J. 2020, 112, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Janas, K.; Cvirkova, M.; Pałągiewicz, A.; Eder, J. Alterations in phenylpropanoid content in soybean roots during low temperature acclimation. Plant Physiol. Biochem. 2000, 38, 587–593. [Google Scholar] [CrossRef]
- Markowski, A. Influence of initial seed moisture and temperature conditions during germination and emergence on seedling survival and yields of soybean (Glycine max L. Merr.). Acta Agrobot. 1982, 35, 43–59. [Google Scholar] [CrossRef] [Green Version]
- Gass, T.; Schori, A.; Fossati, A.; Soldati, A.; Stamp, P. Cold tolerance of soybean (Glycine max (L.) Merr.) during the reproductive phase. Eur. J. Agron. 1996, 5, 71–88. [Google Scholar]
- Staniak, M.; Stępień, A.; Czopek, K. Response of soybean (Glycine max (L.) Merr.) to selected abiotic stresses. Studia i Raporty IUNG-PIB 2018, 57, 63–74. (In Polish) [Google Scholar] [CrossRef]
- Wu, L.; Misselbrook, T.H.; Feng, L.; Wu, L. Assessment of nitrogen uptake and biological nitrogen fixation responses of soybean to nitrogen fertiliser with SPACSYS. Sustainability 2020, 12, 5921. [Google Scholar] [CrossRef]
- Santachiara, G.; Salvagiotti, F.; Rotundo, J.L. Nutritional and environmental effects on biological nitrogen fixation in soybean: A meta-analysis. Field Crop Res. 2019, 240, 106–115. [Google Scholar] [CrossRef]
- Spiekers, H.; Pothast, V. Erfolgreiche Milchviehfütterung; DLG-Verlag: Frankfurt am Main, Germany, 2004. [Google Scholar]
- Rutkowska, A. Biostimulants in modern plant cultivation. Studia i Raporty IUNG–PIB 2016, 48, 65–80. [Google Scholar] [CrossRef]
- Kozak, M.; Malarz, W.; Koteck, A.; Černý, I.; Serafin-Andrzejewska, M. The effect of different sowing rate and Asahi SL biostimulator on chemical composition of soybean seeds and postharvest residues. Oilseed Crops 2008, XXIX, 217–230. [Google Scholar]
- Kocira, A.; Czerwińska, E.; Tomkiewicz, D.; Kornas, R. Microbiological evaluation of three soybean cultivars seeds after biostimulant application. Annu. Set Environ. Prot. 2018, 20, 1710–1726. [Google Scholar]
- Woś, A. Climatic regions of Poland in the light of the frequency of various weather types. IGSO PAS Papers 1993, 20, 97. Available online: https://rcin.org.pl/dlibra/doccontent?id=33464 (accessed on 10 November 2022).
- World Reference Base for Soil Resources. International soil classification system for naming soils and creating legends for soil. In World Soil Resources Reports 106; Field Experiment; Food and Agriculture Organization: Rome, Italy, 2014. [Google Scholar]
- Azam, F.; Farooq, S. An appraisal of methods for measuring symbiotic nitrogen fixation in legumes. Pak. J. Biol. Sci. 2003, 6, 1631–1640. [Google Scholar] [CrossRef] [Green Version]
- Kalembasa, S.; Wysokiński, A.; Kalembasa, D. Quantitative assessment of the process of biological nitrogen reduction by yellow lupine (Lupinus luteus L.). Acta Sci. Pol. Agric. 2014, 13, 5–20. Available online: https://bibliotekanauki.pl/articles/46635 (accessed on 10 November 2022).
- Kapuściński, J.; Nowak, R. The frequency of the occurrence of droughts and post-droughts periods in mid-west Poland on the example of Poznań, Wałcz and Wieluń. In Management and Conservation of the Forest Environment; Climate vs. forest; Miler, A., Ed.; Agricultural University in Poznań: Poznań, Poland, 2003; pp. 76–88. [Google Scholar]
- Bartoszek, K.; Banasiewicz, I. Agrometeorological characteristics of the vegetation period in 2005 against the background of the period of 1951–2005 in the Lublin region. Acta Agrophysica 2007, 9, 275–283. (In Polish) [Google Scholar]
- Hutorowicz, H.; Grabowski, J.; Olba-Zięty, E. Frequency of occurrence of dry spells and droughts in two mesoregions of Masurian Lakeland. Acta Agrophysica 2008, 12, 663–673. (In Polish) [Google Scholar]
- Kunert, K.J.; Vorster, B.J.; Fenta, B.A.; Kibido, T.; Dionisio, G.; Foyer, C.H. Drought stress responses in soybean roots and nodules. Front. Plant Sci. 2016, 7, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korsak-Adamowicz, M.; Starczewski, J.; Dopka, D. Oddziaływanie niektórych zabiegów agrotechnicznych na brodawkowanie soi. Fragm. Agron. 2007, 3, 232–237. (In Polish) [Google Scholar]
- Sadeghipour, O.; Abbasi, S. Soybean Response to Drought and Seed Inoculation. World Appl. Sci. J. 2012, 17, 55–56. [Google Scholar]
- Sinclair, T.R.; Serraj, R. Legume nitrogen fixation and drought. Nature 1995, 378, 344. [Google Scholar] [CrossRef]
- González, E.M.; Gordon, A.J.; James, C.L.; Arrese-Igor, C. The role of sucrose synthase in the response of soybean nodules to drought. J. Exp. Bot. 1995, 46, 1515–1523. [Google Scholar] [CrossRef]
- Arrese-Igor, C.; González, E.M.; Gordon, A.J.; Minchin, F.R.; Gálvez, L.; Royuela, M.; Cabrerizo, P.M.; Aparicio-Tejo, P.M. Sucrose synthase and nodule nitrogen fixation under drought and other environmental stresses. Symbiosis 1999, 27, 189–212. [Google Scholar]
- King, C.A.; Purcell, L.C. Inhibition of N2fixation in soybean is associated with elevated ureides and amino acids. Plant Physiol. 2005, 137, 1389–1396. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Guo, J.; Peng, Y.; Lyu, X.; Liu, B.; Sun, S.; Wang, X. Light-induced mobile factors from shoots regulate rhizobium-triggered soybean root nodulation. Science 2021, 374, 65–71. [Google Scholar] [CrossRef]
- Szparaga, A.; Kocira, S.; Kocira, A.; Czerwinska, E.; Świeca, M.; Lorencowicz, E.; Kornas, R.; Koszel, M.; Oniszczuk, T. Modification of growth, yield, and the nutraceutical and antioxidative potential of soybean through the use of synthetic biostimulants. Front. Plant Sci. 2018, 9, 1401. [Google Scholar] [CrossRef]
- Baglieri, A.; Cadili, V.; Mozzetti Monterumici, C.; Gennari, M.; Tabasso, S.; Montoneri, E. Fertilization of bean plants with tomato plants hydrolysates. Effect on biomass production, chlorophyll content and N assimilation. Sci. Hortic. 2014, 176, 194–199. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef]
- LaMenza, N.C.; Monzon, J.P.; Soecht, J.E.; Grassini, P. Is soybean yield limited by nitrogen supply? Field Crop Res. 2017, 231, 204–212. [Google Scholar]
- Salvagiotti, F.; Specht, J.E.; Cassman, K.G.; Walters, D.T.; Weiss, A.; Dobermann, A. Growth and nitrogen fixation in high-yielding soybean: Impact of nitrogen fertilization. Agron. J. 2009, 101, 958–970. [Google Scholar] [CrossRef]
- Santos, M.A.; Geraldi, I.O.; Garcia, A.A.F.; Bortolatto, N.; Schiavon, A.; Hungria, M. Mapping of QTLs associated with biological nitrogen fixation traits in soybean. Hereditas 2013, 150, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Ray, J.D.; Cregan, P.B.; King, C.A.; Davies, M.K.; Purcell, L.C. Genetics and mapping of quantitative traits for nodule number, weight, and size in soybean (Glycine max L. Merr.). Euphytica 2014, 195, 419–434. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | pH | Corg | Nt | Fet | Bt | Pav | Kav | Mgav |
|---|---|---|---|---|---|---|---|---|
| (in KCl) | g kg−1 | g kg−1 | g kg−1 | g kg−1 | (mg kg−1) | (mg kg−1) | (mg kg−1) | |
| 2017 | 7.0 | 9.5 | 0.77 | 998 | 0.71 | 55.5 | 132.7 | 26.3 |
| 2018 | 7.1 | 9.1 | 0.75 | 995 | 0.65 | 57.0 | 130.5 | 26.1 |
| Value | Classification |
|---|---|
| ≤0.4 | extremely dry |
| (0.4; 0.7> | very dry |
| (0.7;1> | dry |
| (1; 1.3> | quite dry |
| (1.3; 1.6> | optimum |
| (1.6; 2> | moderately wet |
| (2; 2.5> | wet |
| (2.5; 3> | very wet |
| >3 | extremely wet |
| Air Temperature (°C) | ||||||
| Months | May | June | July | Aug | Sept | |
| 2017 | Min. | 3.5 | 5.3 | 14.0 | 13.6 | 8.7 |
| Max. | 20.2 | 22.7 | 21.8 | 24.9 | 18.8 | |
| V | 31.9 | 17.9 | 12.3 | 17.6 | 16.9 | |
| 2018 | Min. | 9.2 | 12.4 | 12.0 | 13.5 | 7.6 |
| Max. | 21.3 | 22.4 | 23.5 | 24.9 | 21.2 | |
| V | 17.6 | 14.9 | 13.7 | 15.6 | 19.3 | |
| Precipitation (mm) | ||||||
| Months | May | June | July | Aug | Sept | |
| 2017 | Min. | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Max. | 22.0 | 12.0 | 25.0 | 20.0 | 23.0 | |
| V | 244.4 | 206.8 | 290.4 | 225.4 | 275.3 | |
| 2018 | Min. | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Max. | 10.0 | 5.0 | 19.0 | 13.0 | 19.0 | |
| V | 205.4 | 267.8 | 181.9 | 264.7 | 183.3 | |
| Insolation (h) | ||||||
| Months | May | June | July | Aug | Sept | |
| 2017 | Min. | 0.2 | 0.8 | 1.4 | 0.0 | 0.0 |
| Max. | 15.2 | 15.5 | 14.7 | 14.1 | 12.5 | |
| V | 49.0 | 45.0 | 43.4 | 40.9 | 105.7 | |
| 2018 | Min. | 0.0 | 0.6 | 0.0 | 0.0 | 0.7 |
| Max. | 15.3 | 15.5 | 14.8 | 14.1 | 11.7 | |
| V | 39.0 | 42.1 | 51.6 | 38.8 | 43.8 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radzka, E.; Rymuza, K.; Wysokiński, A. Weather Conditions and Biostimulants Influence Nitrogen Acquisition from Different Sources by Soybean Plants. Agriculture 2023, 13, 114. https://doi.org/10.3390/agriculture13010114
Radzka E, Rymuza K, Wysokiński A. Weather Conditions and Biostimulants Influence Nitrogen Acquisition from Different Sources by Soybean Plants. Agriculture. 2023; 13(1):114. https://doi.org/10.3390/agriculture13010114
Chicago/Turabian StyleRadzka, Elżbieta, Katarzyna Rymuza, and Andrzej Wysokiński. 2023. "Weather Conditions and Biostimulants Influence Nitrogen Acquisition from Different Sources by Soybean Plants" Agriculture 13, no. 1: 114. https://doi.org/10.3390/agriculture13010114