

Pathogenic and Genetic Characterization of Fusarium verticillioides Strains Collected from Maize and Sorghum Kernels

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fusarium spp. Isolation
2.2. Molecular Identification and Mating Type Determination
2.3. Total Fumonisin Production
2.4. Pathogenicity Assays
2.5. Microsatellite Marker Analysis
2.6. Statistical Analysis
3. Results
3.1. Incidence of Fusarium verticillioides and Related Species in Maize and Sorghum Samples
3.2. Mating Type Determination
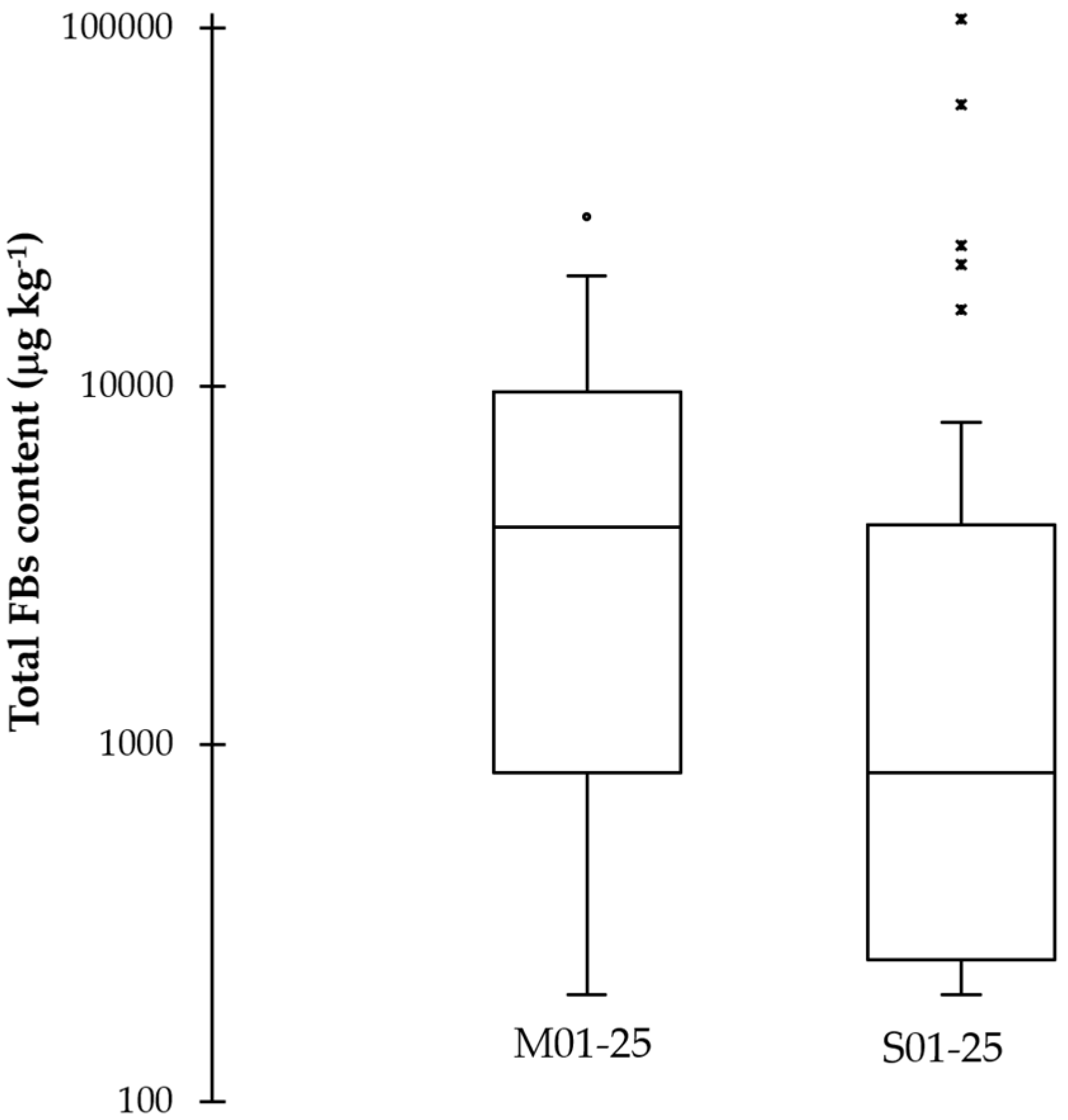
3.3. Fumonisin Quantification
3.4. Effect of Fusarium verticillioides Strains on Seed Germination
3.5. Effect of Fusarium verticillioides Strains on Root Development
3.6. Effect of Fusarium verticillioides Strains on Shoot Development
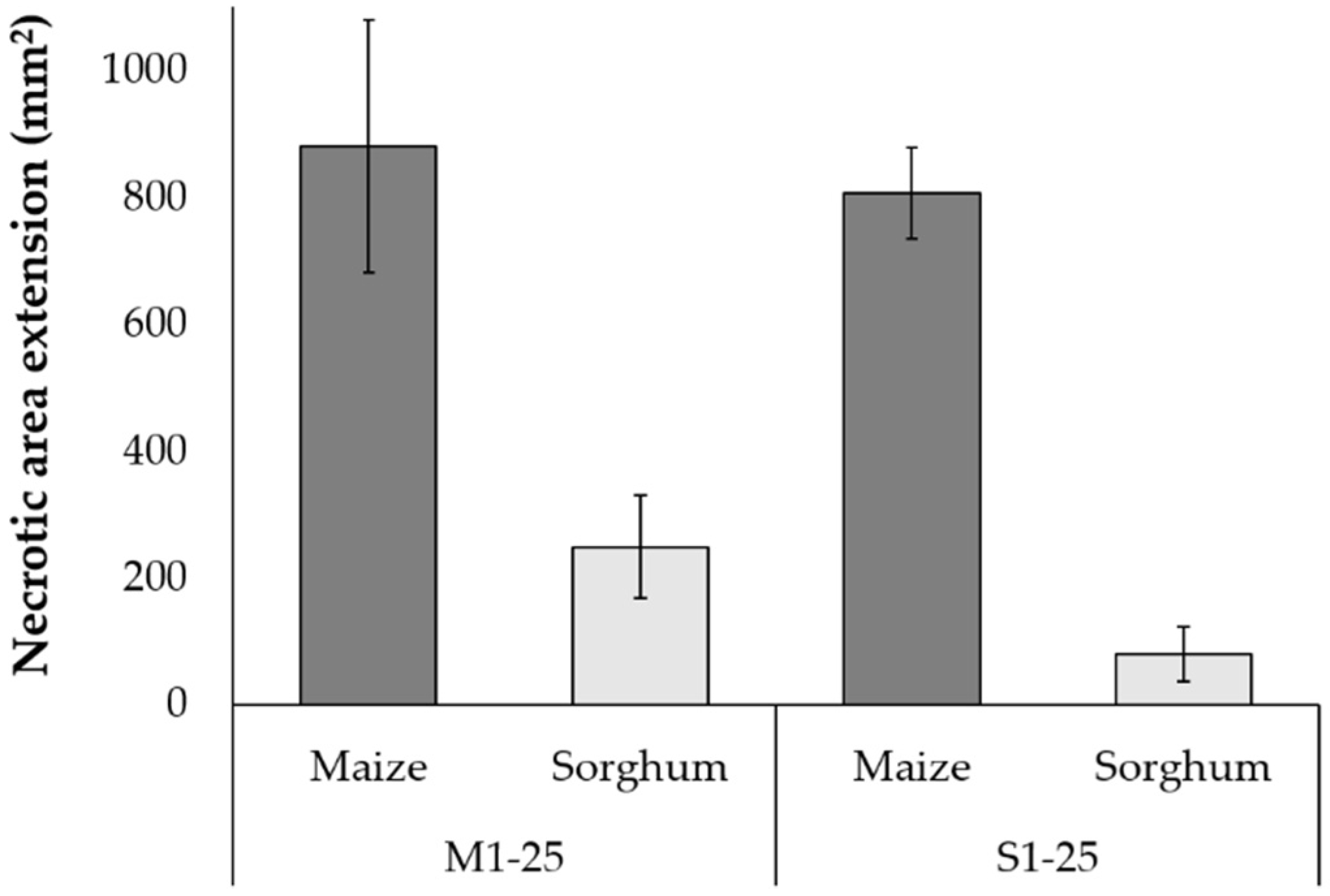
3.7. Effect of Fusarium verticillioides Strains on Detached Leaf Assays (DLA)
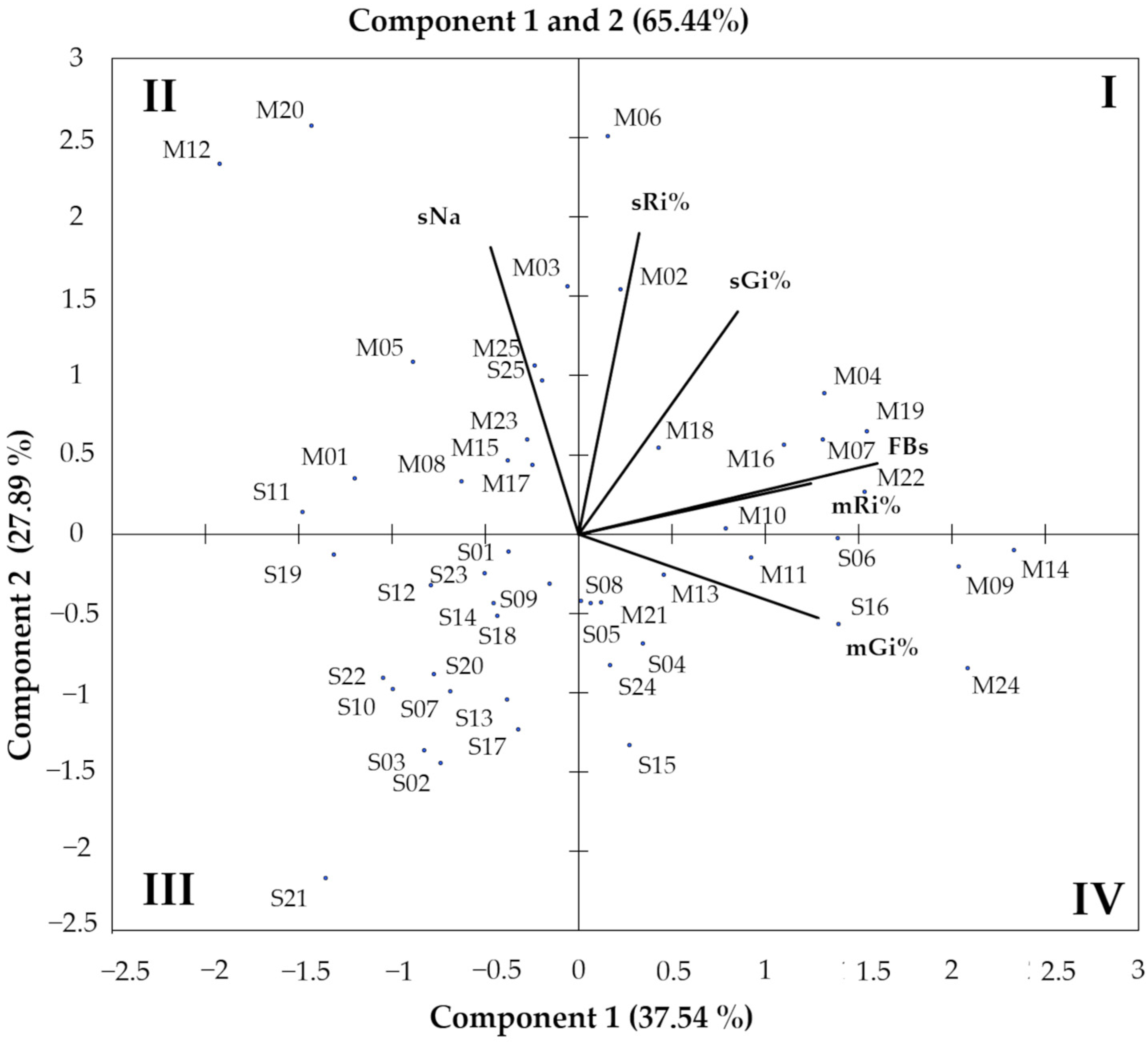
3.8. Clusterization of Fusarium verticillioides Strains Based on Pathogenic Parameters
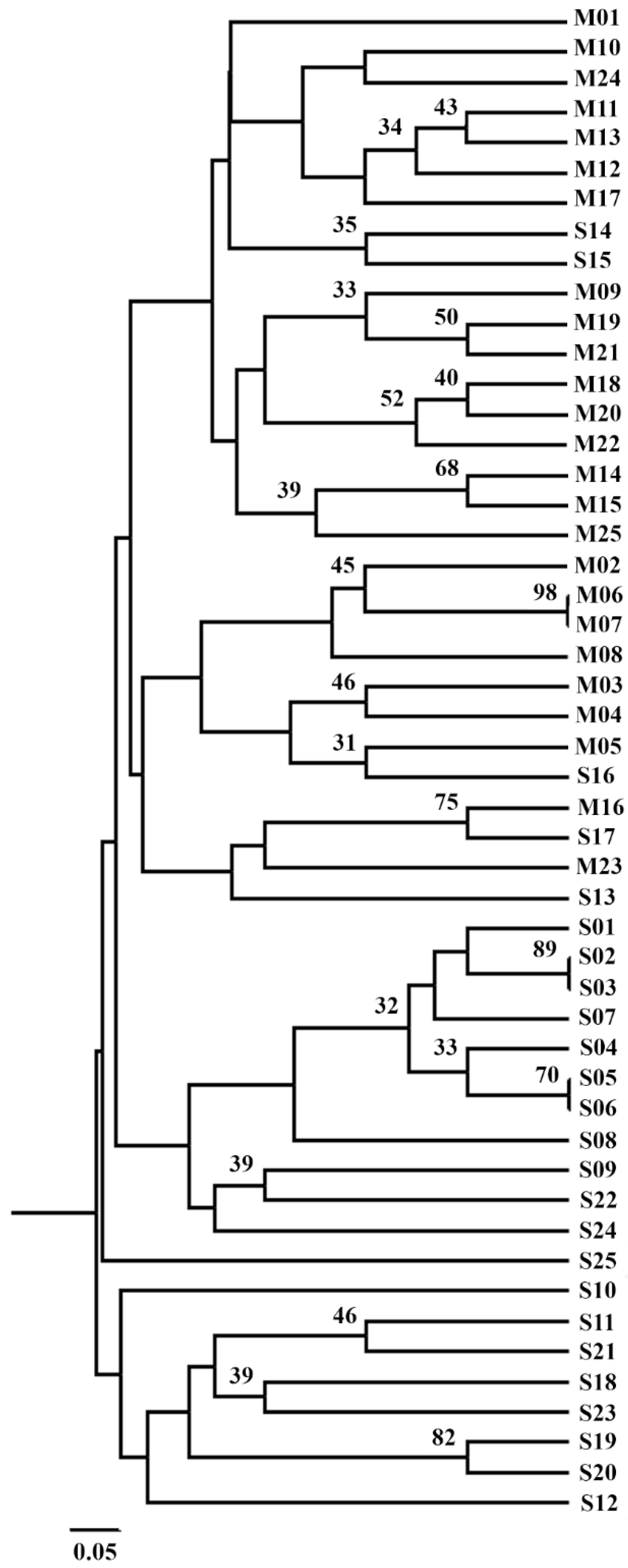
3.9. Microsatellite Marker Analysis
3.10. Fusarium verticillioides Clusterization Based on Microsatellite Markers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Warr, B.; Schandl, H.; Ayres, R.U. Long Term Trends in Resource Exergy Consumption and Useful Work Supplies in the UK, 1900 to 2000. Ecol. Econ. 2008, 68, 126–140. [Google Scholar] [CrossRef]
- Italian National Institute of Statistics. Estimate of Crop, Flower and Pot Plant Production and Area 2020. Available online: http://dati.istat.it/Index.aspx?lang=en&SubSessionId=38f169a7-56dc-470f-9fcd-d89bc3eff2c5 (accessed on 18 July 2022).
- Bruns, H.A. Controlling Aflatoxin and Fumonisin in Maize by Crop Management. J. Toxicol. Toxin Rev. 2003, 22, 153–173. [Google Scholar] [CrossRef]
- Deepa, N.; Sreenivasa, M.Y. Fumonisins: A Review on Its Global Occurrence, Epidemiology, Toxicity and Detection. J. Vet. Med. Res. 2017, 4, 1093. [Google Scholar]
- Ackerman, A.; Wenndt, A.; Boyles, R. The Sorghum Grain Mold Disease Complex: Pathogens, Host Responses, and the Bioactive Metabolites at Play. Front. Plant Sci. 2021, 12, 660171. [Google Scholar] [CrossRef] [PubMed]
- Mohamed Nor, N.M.I.; Salleh, B.; Leslie, J.F. Fusarium Species from Sorghum in Thailand. Plant Pathol. J. 2019, 35, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Palmero Llamas, D.; Gil Serna, J.; Galvez Patón, L.; Curt Fernández de la Mora, M.; Cara Garcia, M.; de Tello Marquina, J. First Report of Fusarium verticillioides Causing Stalk and Root Rot of Sorghum in Spain. Plant Dis. 2012, 96, 584.2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesso, T.T.; Ochanda, N.; Little, C.R.; Claflin, L.; Tuinstra, M.R. Analysis of Host Plant Resistance to Multiple Fusarium Species Associated with Stalk Rot Disease in Sorghum [Sorghum bicolor (L.) Moench]. Field Crops Res. 2010, 118, 177–182. [Google Scholar] [CrossRef]
- Deepa, N.; Nagaraja, H.; Sreenivasa, M.Y. Prevalence of Fumonisin Producing Fusarium verticillioides Associated with Cereals Grown in Karnataka (India). Food Sci. Hum. Wellness 2016, 5, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wen, J.; Tang, Y.; Shi, J.; Mu, G.; Yan, R.; Cai, J.; Long, M. Research Progress on Fumonisin B1 Contamination and Toxicity: A Review. Molecules 2021, 26, 5238. [Google Scholar] [CrossRef]
- Hanvi, D.M.; Lawson-Evi, P.; De Boevre, M.; Goto, C.E.; De Saeger, S.; Eklu-Gadegbeku, K. Natural Occurrence of Mycotoxins in Maize and Sorghum in Togo. Mycotoxin Res. 2019, 35, 321–327. [Google Scholar] [CrossRef]
- Vismer, H.F.; Shephard, G.S.; Rheeder, J.P.; van der Westhuizen, L.; Bandyopadhyay, R. Relative Severity of Fumonisin Contamination of Cereal Crops in West Africa. Food Addit. Contam. Part A 2015, 32, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Dastjerdi, R.; Karlovsky, P. Systemic Infection of Maize, Sorghum, Rice, and Beet Seedlings with Fumonisin-Producing and Nonproducing Fusarium verticillioides Strains. Plant Pathol. J. 2015, 31, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Snyder, B.A.; Nicholson, R.L. Synthesis of Phytoalexins in Sorghum as a Site-Specific Response to Fungal Ingress. Science 1990, 248, 1637–1639. [Google Scholar] [CrossRef]
- Jambunathan, R.; Kherdekar, M.S.; Stenhouse, J.W. Sorghum Grain Hardness and Its Relationship to Mold Susceptibility and Mold Resistance. J. Agric. Food Chem. 1992, 40, 1403–1408. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, A.; Davidson, C.E. Estimation Method for Serial Dilution Experiments. J. Microbiol. Methods 2014, 107, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 0-470-27646-0. [Google Scholar]
- Tuite, J. Plant Pathological Methods. Fungi and Bacteria; CABI: Wallingford, UK, 1969; p. 239. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Mulè, G.; Susca, A.; Stea, G.; Moretti, A. A Species-Specific PCR Assay Based on the Calmodulin Partial Gene for Identification of Fusarium verticillioides, F. proliferatum and F. subglutinans. Eur. J. Plant Pathol. 2004, 110, 495–502. [Google Scholar] [CrossRef]
- Steenkamp, E.T.; Wingfield, B.D.; Coutinho, T.A.; Zeller, K.A.; Wingfield, M.J.; Marasas, W.F.; Leslie, J.F. PCR-Based Identification of MAT-1 and MAT-2 in the Gibberella fujikuroi Species Complex. Appl. Environ. Microbiol. 2000, 66, 4378–4382. [Google Scholar] [CrossRef] [Green Version]
- Yates, I.E.; Arnold, J.W.; Hinton, D.M.; Basinger, W.; Walcott, R.R. Fusarium verticillioides Induction of Maize Seed Rot and Its Control. Can. J. Bot. 2003, 81, 422–428. [Google Scholar] [CrossRef]
- Browne, R.A.; Cooke, B.M. Development and Evaluation of an in Vitro Detached Leaf Assay Forc Pre-Screening Resistance to Fusarium Head Blight in Wheat. Eur. J. Plant Pathol. 2004, 110, 91–102. [Google Scholar] [CrossRef]
- Xu, R.; ZhenDong, Z.; HongJie, L.; CanXing, D.; XiaoMing, W. SSR marker development and analysis of genetic diversity of Fusarium verticillioides isolated from maize in China. Sci. Agric. Sin. 2012, 45, 52–66. [Google Scholar]
- Leyva-Madrigal, K.Y.; Larralde-Corona, C.P.; Calderón-Vázquez, C.L.; Maldonado-Mendoza, I.E. Genome Distribution and Validation of Novel Microsatellite Markers of Fusarium verticillioides and Their Transferability to Other Fusarium Species. J. Microbiol. Methods 2014, 101, 18–23. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley-Inter-Science: New York, NY, USA, 1969. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Vallvé, S.; Palau, J.; Romeu, A. Horizontal Gene Transfer in Glycosyl Hydrolases Inferred from Codon Usage in Escherichia coli and Bacillus subtilis. Mol. Biol. Evol. 1999, 16, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.R.; Rohlf, F.J. The Comparison of Dendrograms by Objective Methods. Taxon 1962, 11, 33–40. [Google Scholar] [CrossRef]
- Sauerborn, J.; Sprich, H.; Mercer-Quarshie, H. Crop Rotation to Improve Agricultural Production in Sub-Saharan Africa. J. Agron. Crop Sci. 2000, 184, 67–72. [Google Scholar] [CrossRef]
- Wu, A.-L.; Jiao, X.-Y.; Fan, F.-F.; Wang, J.-S.; Guo, J.; Dong, E.-W.; Wang, L.-G.; Shen, X.-M. Effect of Continuous Sorghum Cropping on the Rhizosphere Microbial Community and the Role of Bacillus amyloliquefaciens in Altering the Microbial Composition. Plant Growth Regul. 2019, 89, 299–308. [Google Scholar] [CrossRef]
- Jaime-Garcia, R.; Cotty, P.J. Crop Rotation and Soil Temperature Influence the Community Structure of Aspergillus flavus in Soil. Soil Biol. Biochem. 2010, 42, 1842–1847. [Google Scholar] [CrossRef]
- Phillips, N.J.; Uyemoto, J.K.; Wilson, D.L. Maize Chlorotic Mottle Virus and Crop Rotation: Effect of Sorghum on Virus Incidence. Plant Dis. 1982, 66, 376–379. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Fermin-Pérez, R.A.; Prom, L.K.; Cooper, E.A.; Bean, S.; Rooney, W.L. Genome-Wide Association Mapping of Grain Mold Resistance in the US Sorghum Association Panel. Plant Genome 2019, 12, 170099. [Google Scholar] [CrossRef] [Green Version]
- Lanza, F.E.; Zambolim, L.; da Costa, R.V.; Queiroz, V.A.V.; Cota, L.V.; da Silva, D.D.; de Souza, A.G.C.; Figueiredo, J.E.F. Prevalence of Fumonisin-Producing Fusarium Species in Brazilian Corn Grains. Crop Prot. 2014, 65, 232–237. [Google Scholar] [CrossRef]
- Silva, J.J.; Viaro, H.P.; Ferranti, L.S.; Oliveira, A.L.M.; Ferreira, J.M.; Ruas, C.F.; Ono, E.Y.S.; Fungaro, M.H.P. Genetic Structure of Fusarium Verticillioides Populations and Occurrence of Fumonisins in Maize Grown in Southern Brazil. Crop Prot. 2017, 99, 160–167. [Google Scholar] [CrossRef]
- Divakara, S.T.; Santosh, P.; Aiyaz, M.; Venkata Ramana, M.; Hariprasad, P.; Nayaka, S.C.; Niranjana, S.R. Molecular Identification and Characterization of Fusarium Spp. Associated with Sorghum Seeds. J. Sci. Food Agric. 2014, 94, 1132–1139. [Google Scholar] [CrossRef]
- Yassin, M.A.; El-Samawaty, A.-R.; Bahkali, A.; Moslem, M.; Abd-Elsalam, K.A.; Hyde, K.D. Mycotoxin-Producing Fungi Occurring in Sorghum Grains from Saudi Arabia. Fungal Divers. 2010, 44, 45–52. [Google Scholar] [CrossRef]
- Fraser, J.A.; Heitman, J. Chromosomal Sex-Determining Regions in Animals, Plants and Fungi. Curr. Opin. Genet. Dev. 2005, 15, 645–651. [Google Scholar] [CrossRef]
- Fraser, J.A.; Heitman, J. Evolution of Fungal Sex Chromosomes. Mol. Microbiol. 2004, 51, 299–306. [Google Scholar] [CrossRef]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Structure and Evolution of the Fusarium Mating Type Locus: New Insights from the Gibberella fujikuroi Complex. Fungal Genet. Biol. 2011, 48, 731–740. [Google Scholar] [CrossRef] [Green Version]
- da Silva, V.N.; Fernandes, F.M.C.; Cortez, A.; Ribeiro, D.H.B.; de Almeida, A.P.; Hassegawa, R.H.; Corrêa, B. Characterization and Genetic Variability of Fusarium verticillioides Strains Isolated from Corn and Sorghum in Brazil Based on Fumonisins Production, Microsatellites, Mating Type Locus, and Mating Crosses. Can. J. Microbiol. 2006, 52, 798–804. [Google Scholar] [CrossRef]
- Covarelli, L.; Stifano, S.; Beccari, G.; Raggi, L.; Lattanzio, V.M.T.; Albertini, E. Characterization of Fusarium verticillioides Strains Isolated from Maize in Italy: Fumonisin Production, Pathogenicity and Genetic Variability. Food Microbiol. 2012, 31, 17–24. [Google Scholar] [CrossRef]
- Das, I.K.; Kumar, B.S.V.; Ratnavathi, C.V.; Komala, V.V.; Annapurna, A.; Seetharama, N. Toxigenicity of Fusarium Isolates and Fumonisin B1 Contamination in Rainy Season Sorghum (Sorghum bicolor). Indian J. Agric. Sci. 2010, 80, 724–729. [Google Scholar]
- Deepa, N.; Rakesh, S.; Sreenivasa, M.Y. Morphological, Pathological and Mycotoxicological Variations among Fusarium verticillioides Isolated from Cereals. 3 Biotech 2018, 8, 105. [Google Scholar] [CrossRef]
- Venturini, G.; Assante, G.; Toffolatti, S.L.; Vercesi, A. Pathogenicity Variation in Fusarium verticillioides Populations Isolated from Maize in Northern Italy. Mycoscience 2013, 54, 285–290. [Google Scholar] [CrossRef]
- Jensen, F. Relationships between Seed Germination, Fumonisin Content, and Fusarium verticillioides Infection in Selected Maize Samples from Different Regions of Costa Rica. Plant Pathol. 1998, 47, 609–614. [Google Scholar]
- Dykes, L.; Rooney, L.W. Sorghum and Millet Phenols and Antioxidants. J. Cereal Sci. 2006, 44, 236–251. [Google Scholar] [CrossRef]
- Maurya, S.; Singh, R.; Singh, D.P.; Singh, H.B.; Srivastava, J.S.; Singh, U.P. Phenolic Compounds of Sorghum vulgare in Response to Sclerotium rolfsii Infection. J. Plant Interact. 2007, 2, 25–29. [Google Scholar] [CrossRef]
- Shen, S.; Huang, R.; Li, C.; Wu, W.; Chen, H.; Shi, J.; Chen, S.; Ye, X. Phenolic Compositions and Antioxidant Activities Differ Significantly among Sorghum Grains with Different Applications. Molecules 2018, 23, 1203. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum Grain: From Genotype, Nutrition, and Phenolic Profile to Its Health Benefits and Food Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef] [Green Version]
- Nida, H.; Lee, S.; Li, Y.; Mengiste, T. Transcriptome Analysis of Early Stages of Sorghum Grain Mold Disease Reveals Defense Regulators and Metabolic Pathways Associated with Resistance. BMC Genom. 2021, 22, 295. [Google Scholar] [CrossRef]
- Cheynier, V. Phenolic Compounds: From Plants to Foods. Phytochem. Rev. 2012, 11, 153–177. [Google Scholar] [CrossRef]
- Dicko, M.H.; Gruppen, H.; Barro, C.; Traore, A.S.; van Berkel, W.J.H.; Voragen, A.G.J. Impact of Phenolic Compounds and Related Enzymes in Sorghum Varieties for Resistance and Susceptibility to Biotic and Abiotic Stresses. J. Chem. Ecol. 2005, 31, 2671–2688. [Google Scholar] [CrossRef] [PubMed]
- Takele, A.; Farrant, J. Enzymatic Antioxidant Defence Mechanisms of Maize and Sorghum after Exposure to and Recovery from Pre- and Post-Flowering Dehydration. Acta Agron. Hung. 2009, 57, 445–459. [Google Scholar] [CrossRef]
- Ortiz, C.S.; Richards, C.; Terry, A.; Parra, J.; Shim, W. Genetic Variability and Geographical Distribution of Mycotoxigenic Fusarium verticillioides Strains Isolated from Maize Fields in Texas. Plant Pathol. J. 2015, 31, 203–211. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus Name | Repeated Motif | Primer Name and Sequence (5′–3′) |
|---|---|---|
| Fv-98 | ATCC | Fv-98F AAACAAGATGCGGTCCATTC |
| Fv-98R GGATCGGAGGAGAATCAACA | ||
| Fv-114 | GTCT | Fv-114F CGAATGCCTTGATCTGCTTC |
| Fv-114R GAGAATCCTGTTTGCGTGGT | ||
| Fv-120 | TTG | Fv-120F GTAGCGCGGTAAGAAGATGC |
| Fv-120R AGTCGAAGCCCAACTGAAGA | ||
| Fv-312 | CAGA | Fv-312F TTTCCGAATTCCTGGATCTG |
| Fv-312R GACGCAGTTTGCACAAGGTA | ||
| Fv-338 | AGCAG | Fv-338F TAGACCAGGCAGACGAGACA |
| Fv-338R TGTGAGTGGGTGAGAGTGGA |
| F. verticillioides | F. proliferatum | F. oxysporum | Other Fusaria * | FBs Content | |||||
|---|---|---|---|---|---|---|---|---|---|
| CFU g−1 | % | CFU g−1 | % | CFU g−1 | % | CFU g−1 | % | μg kg−1 | |
| Maize | 9757 ± 3063 | 88.8 | 461 ± 145 | 4.2 | 654 ± 205 | 6.0 | 110 ± 18 | 1.0 | 4050 ± 304 |
| Sorghum | 388 ± 68 | 58.8 | 186 ± 102 | 28.2 | 20 ± 18 | 3.0 | 65 ± 19 | 9.9 | <200 |
| Maize | Sorghum | |||
|---|---|---|---|---|
| Strain | FBs μg kg−1 | Strain | FBs μg kg−1 | |
| MAT-1 | M06 | 20,300 | S04 | 24,800 |
| M17 | 13,250 | S09 | 21,900 | |
| M22 | 11,170 | S25 | 16,400 | |
| M14 | 7700 | S02 | 2490 | |
| M15 | 6880 | S08 | 1350 | |
| M04 | 6430 | S07 | 1340 | |
| M02 | 2950 | S05 | 830 | |
| M09 | 2380 | S12 | 310 | |
| M18 | 1820 | S10 | 300 | |
| M11 | 830 | S21 | 270 | |
| M13 | 700 | S23 | 240 | |
| M12 | 570 | S03 | 230 | |
| M05 | 240 | S22 | 230 | |
| M01 | 220 | S24 | 230 | |
| M25 | 200 | S11 | 200 | |
| - | - | S20 | 200 | |
| MAT-2 | M19 | 29,600 | S16 | 106,300 |
| M24 | 19,400 | S06 | 61,200 | |
| M08 | 16,780 | S13 | 7950 | |
| M23 | 9640 | S17 | 4100 | |
| M16 | 5570 | S15 | 2180 | |
| M21 | 5140 | S01 | 1930 | |
| M07 | 4050 | S14 | 590 | |
| M10 | 3100 | S18 | 290 | |
| M03 | 850 | S19 | 250 | |
| M20 | 740 | - | - | |
| Microsatellite Marker | Strains | Polymorphic Bands/Tot | H’ Index | J’ Index | BC Dissimilarity |
|---|---|---|---|---|---|
| Fv-98 | M1-25 | 3/7 | 1.040 | 0.946 | 0.50 |
| S1-25 | 6/7 | 1.223 | 0.679 | ||
| Fv-114 | M1-25 | 5/9 | 1.332 | 0.828 | 0.84 |
| S1-25 | 6/9 | 1.555 | 0.868 | ||
| Fv-120 | M1-25 | 9/11 | 2.027 | 0.926 | 0.56 |
| S1-25 | 6/11 | 1.658 | 0.925 | ||
| Fv-312 | M1-25 | 6/10 | 1.432 | 0.799 | 0.68 |
| S1-25 | 9/10 | 1.921 | 0.875 | ||
| Fv-338 | M1-25 | 6/12 | 1.457 | 0.813 | 0.68 |
| S1-25 | 11/12 | 2.123 | 0.885 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrigo, D.; Mondin, M.; Raiola, A. Pathogenic and Genetic Characterization of Fusarium verticillioides Strains Collected from Maize and Sorghum Kernels. Agriculture 2023, 13, 105. https://doi.org/10.3390/agriculture13010105
Ferrigo D, Mondin M, Raiola A. Pathogenic and Genetic Characterization of Fusarium verticillioides Strains Collected from Maize and Sorghum Kernels. Agriculture. 2023; 13(1):105. https://doi.org/10.3390/agriculture13010105
Chicago/Turabian StyleFerrigo, Davide, Massimiliano Mondin, and Alessandro Raiola. 2023. "Pathogenic and Genetic Characterization of Fusarium verticillioides Strains Collected from Maize and Sorghum Kernels" Agriculture 13, no. 1: 105. https://doi.org/10.3390/agriculture13010105