
Impact of Temperature on Age–Stage, Two-Sex Life Table Analysis of a Chinese Population of Bean Bug, Riptortus pedestris (Hemiptera: Alydidae)
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Host Plant Production and Insect Rearing
2.2. Experimental Setup
2.3. Estimation of Developmental Threshold Temperature and Effective Accumulated Temperature
2.4. Estimation of Population Trend Index
2.5. Statistical Analysis
3. Results
3.1. Developmental Period, Longevity and Fecundity of R. pedestris on Different Temperature
3.2. Developmental Threshold and Effective Accumulated Temperature
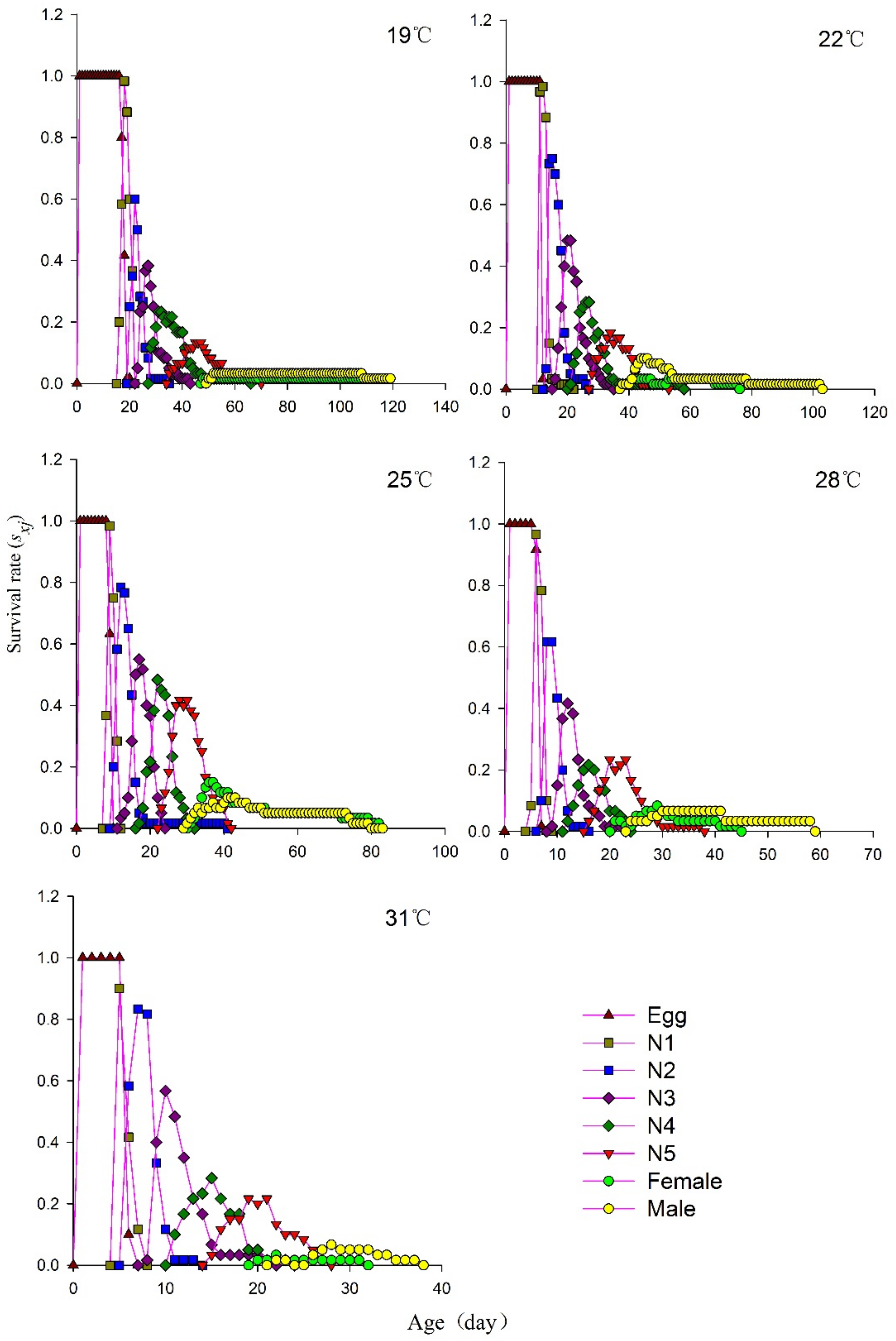
3.3. Age–Stage-Specific Survival Rate
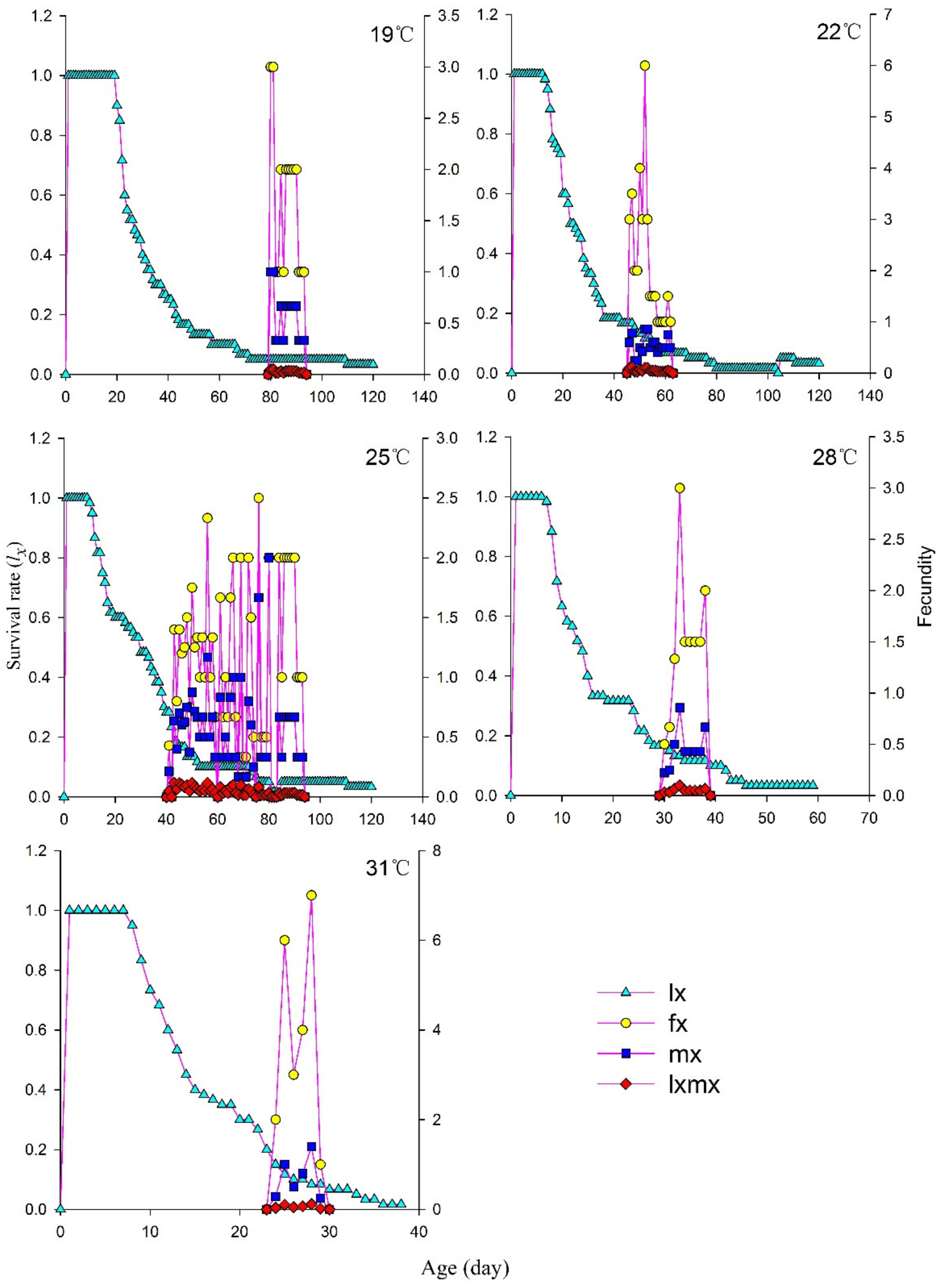
3.4. Age-Specific Survival and Age–Stage-Specific Fecundity
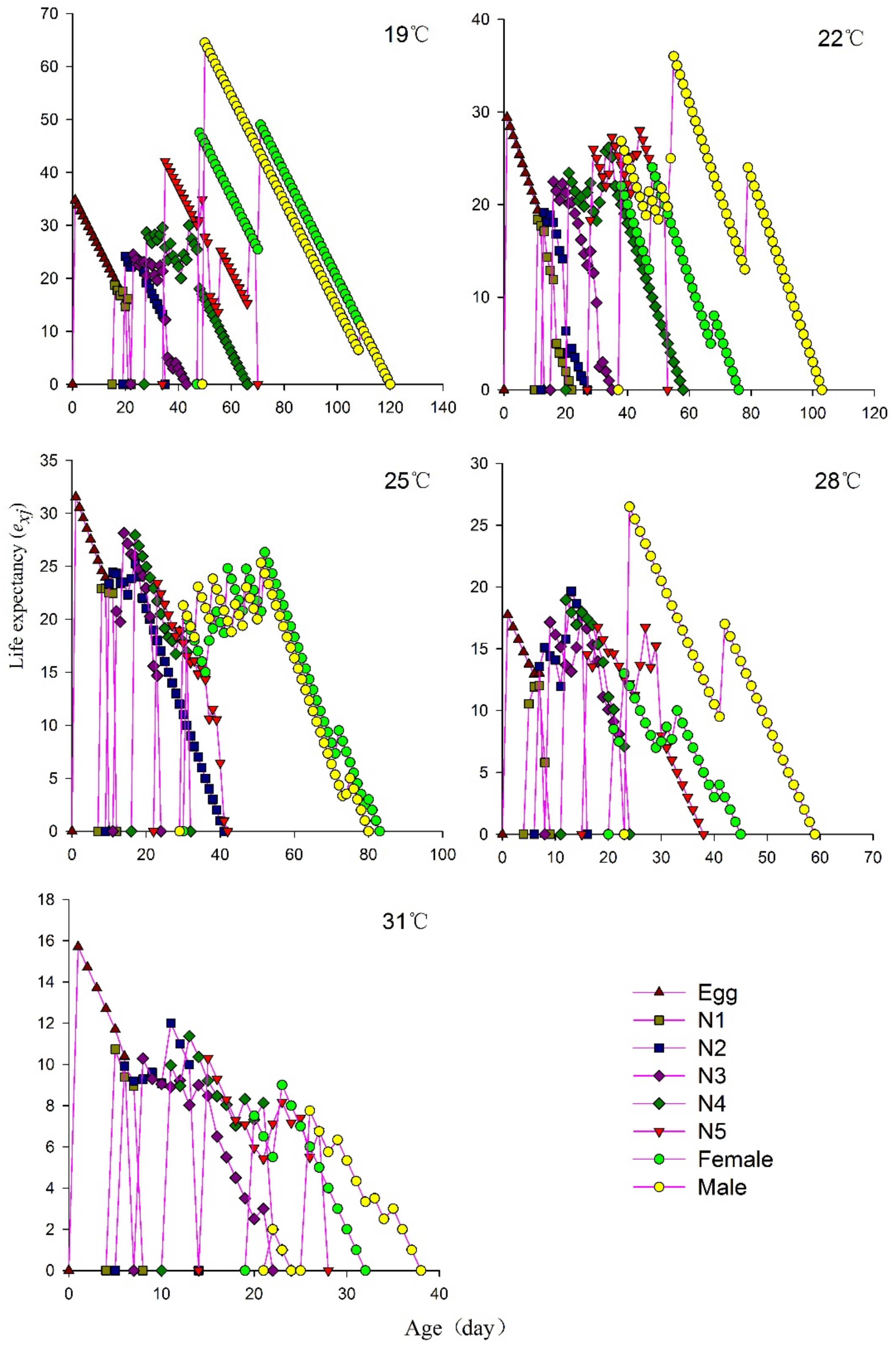
3.5. Age–Stage-Specific Life Expectancy
3.6. Age–Stage-Specific Reproductive Values
3.7. Population Parameters at Different Temperatures
3.8. Effect of Temperature on Population Parameters
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, S.S. Soybean Pest Comprehensive Prevention and Control Technology and Technology; Jilin Publishing Group Co.: Changchun, China, 2013. [Google Scholar]
- Kim, G.H.; Ahn, Y.J.; Cho, K.Y. Effect of diflubenzuron on longevity and reproduction of adult bean bug (Hemiptera: Alydidae). J. Econ. Entomol. 1992, 85, 664–668. [Google Scholar] [CrossRef]
- Tabuchi, K.; Moriya, S.; Ishizaki, M. Locomotion ability variation among instars of the bean bug, Riptortus pedestris (Fabricius) (Heteroptera: Alydidae) nymphs. Environ. Entomol. 2007, 36, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Parvathy, V. Ecological Perspectives and Host Plant Resistance Studies of Pod Feeders in Field Bean, Lablab purpureus (L.) Sweet. Bachelor’s Thesis, Acharya N. G. Ranga Agricultural University, Pradesh, India, 2011. [Google Scholar]
- Zhang, X.X.; Wang, M.; Wu, T.T.; Wu, C.X.; Jiang, B.J.; Guo, C.H.; Han, T.F. Physiological and molecular studies of staygreen caused by pod removal and seed injury in soybean. Crop J. 2016, 4, 435–443. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, J.J.; Li, F.Q.; Yan, Q.; Meng, L.; Li, B.P. Chemical polymorphism regulates the attractiveness to nymphs in the bean bug Riptortus pedestris. J. Pest Sci. 2020, 94, 463–472. [Google Scholar] [CrossRef]
- Chen, Q.E.; Bai, J.K. Atlas of Soybean Diseases and Insects in China; Jilin Technical Science Press: Changchun, China, 1987. [Google Scholar]
- Gao, Y.; Shi, S.S.; Xu, M.L.; Cui, J. Current research on soybean integrated pest management in China. Oil Crop Sci. 2018, 3, 215–227. [Google Scholar]
- Gao, Y.; Chen, J.H.; Shi, S.S. Research progress on soybean stink bug (Riptortus pedestris). Chin. J. Oil Crop Sci. 2019, 41, 804–815. [Google Scholar]
- Li, Y.Q.; Wang, Y.X. Causes and countermeasures for reluctant ripen and few pods of soybean in Wuyanog county in 2010. Mod. Agric. Technol. 2012, 1, 117–118. [Google Scholar]
- Hua, F.J.; Zhang, K.; Cao, P.P.; Wang, Y.Z.; Gao, F.J.; Wang, C.Y. Influence factors and prevention measures of podding without pea in summer soybean. Shandong Agric. Sci. 2019, 51, 156–162. [Google Scholar]
- Hassall, C.; Thompson, D.J.; French, G.C.; Harvey, I.F. Historical changes in the phenology of British odonata are related to climate. Glob. Chang. Biol. 2007, 13, 83–95. [Google Scholar] [CrossRef]
- Khadioli, N.; Tonnang, Z.E.H.; Muchugu, E.; Ong’amo, G.; Achia, T.; Kipchirchir, I.; Kroschel, J.; Le Ru, B. Effect of temperature on the phenology of Chilo partellus (Swinhoe) (Lepidoptera, Crambidae) simulation and visualization of the potential future distribution of C. partellus in Africa under warmer temperatures through the development of life-table parameters. Bull. Entomol. Res. 2014, 104, 809–822. [Google Scholar]
- Régnière, J.; Powell, J.; Bentz, B.; Nealis, V. Effects of temperature on development, survival and reproduction of insects: Experimental design, data analysis and modeling. J. Insect Physiol. 2012, 58, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.S.; Guo, J.Y.; Michaud, J.P.; Li, M.; Wan, F.H. Variation in cold hardiness among geographic populations of the ragweed beetle, Ophraella communa LeSage (Coleoptera: Chrysomelidae), a biological control agent of Ambrosia artemisiifolia L. (Asterales: Asteraceae), in China. Biol. Invasions 2011, 13, 659–667. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, J.P.; Yang, Y.Z.; Lu, Y.H. Transcriptome analysis of Propylaea quatuordecimpunctata L. (Coleoptera: Coccinellidae) under high temperature stress. Agriculture 2022, 12, 1088. [Google Scholar]
- Ma, L.; Gao, S.; Wen, J.; Zong, S.; Xu, Z. Effective accumulated temperature and developmental threshold temperature for Semanotus bifasciatus (Motschulsky) in Beijing. For. Stud. China 2008, 10, 125–129. [Google Scholar] [CrossRef]
- Ahn, J.J.; Choi, K.S.; Koh, S. Effects of temperature on the development, fecundity, and life table parameters of Riptortus pedestris (Hemiptera: Alydidae). Appl. Entomol. Zool. 2018, 54, 63–74. [Google Scholar] [CrossRef]
- Chen, J.H.; Cui, J.; Tang, J.W.; Bi, R.; Zhang, J.P.; Shi, S.S. Effects of temperature on the growth and development of Riptortus pedestris Fabricius. Chin. J. Oil Crop Sci. 2018, 40, 579–584. [Google Scholar]
- Ji, Y.; Li, G.; Zhou, C.; Yin, S. Influence of temperature on the development and reproduction of Cinara cedri (Hemiptera: Aphidoidea: Lachninae). Bull. Entomol. Res. 2021, 111, 579–584. [Google Scholar] [CrossRef]
- Ali, M.Y.; Naseem, T.; Arshad, M.; Ashraf, I.; Rizwan, M.; Tahir, M.; Rizwan, M.; Sayed, S.; Ullah, M.I.; Khan, R.R.; et al. Host-plant variations affect the biotic potential, survival, and population projection of Myzus persicae (Hemiptera: Aphididae). Insects 2021, 12, 375. [Google Scholar] [CrossRef]
- Ali, S.; Li, S.; Jaleel, W.; Khan, M.M.; Wang, J.; Zhou, X. Using a two-sex life table tool to calculate the fitness of Orius strigicollis as a predator of Pectinophora gossypiella. Insects 2020, 11, 275. [Google Scholar] [CrossRef]
- Ahn, J.J.; Choi, K.S. Population parameters and growth of Riptortus pedestris (Fabricius) (Hemiptera: Alydidae) under fluctuating temperature. Insects 2022, 13, 113. [Google Scholar] [CrossRef]
- Li, D.M.; Wang, M.M. Rapid development starting point estimates and effective accumulated temperature method research. Insect Knowl. 1986, 23, 184–187. [Google Scholar]
- Li, T.L.; Wang, Y.Q.; Ma, J.F.; Liu, L.; Hao, Y.T.; Dong, C.; Gan, Y.J.; Dong, Z.P.; Wang, Q.Y. The effects of temperature on the development of the moth Athetis lepigone, and a prediction of field occurrence. J. Insect Sci. 2013, 13, 1–13. [Google Scholar]
- Chi, H. Two Sex–MS Chart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. Available online: https://www.researchgate.net/publication/267522110_Age-stage_two-sex_life_table_Theory_data_analysis_and_application (accessed on 28 February 2020).
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; Chapman and Hall: New York, NY, USA, 1993. [Google Scholar]
- Mou, D.F.; Lee, C.C.; Smith, C.L.; Chi, H. Using viable eggs to accurately determine the demographic and predation potential of Harmonia dimidiata (Coleoptera: Coccinellidae). J. Appl. Entomol. 2015, 139, 579–591. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Goodman, D. Optimal Life Histories, Optimal Notation, and the Value of Reproductive Value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [PubMed]
- Yang, H.M.; Macoris, M.L.G.; Galvani, K.C.; Andrighetti, M.T.M.; Wanderley, D.M.V. Assessing the effects of temperature on the population of Aedes aegypti, the vector of dengue. Epidemiol. Infect. 2009, 137, 1188–1202. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.L.; Guo, H.M.; Sun, S.L. Effects of temperature on growth, development, and reproduction of peach borer. Chin. J. Entomol. 2012, 55, 561–569. [Google Scholar]
- Milosavljević, I.; McCalla, K.A.; Ratkowsky, D.A.; Hoddle, M.S. Effects of constant and fluctuating temperatures on development rates and longevity of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae). J. Econ. Entomol. 2019, 112, 1062–1072. [Google Scholar] [CrossRef]
- Available online: https://m.tianqi.com/qiwen/city_cangzhou/?ivk_sa=1024320u (accessed on 1 September 2022).
- Ma, B.; Jiang, H.Y. Effective accumulated temperature law and its application. Heilongjiang Sci. Technol. Inf. 2007, 20, 161. [Google Scholar]
- Xie, H.; Chen, L.J.; Han, J.; Wang, C. Damage characteristics and control methods of Riptortus pedestris in soybean. Soybean Sci. Technol. 2016, 6, 11–13. [Google Scholar]
- Chi, H. Timing of control based on the stage structure of pest populations: A simulation approach. J. Econ. Entomol. 1990, 83, 1143–1150. [Google Scholar] [CrossRef]
- Field, C.B.; Barros, T.F.; Stocker, D.; Qin, D.J.; Dokken, K.L.; Ebi, M.D.; Mastrandrea, K.J.; Mach, G.K.; Plattner, S.K.; Allen, M.; et al. (Eds.) Managing the risks of extreme events and disasters to advance climate change adaptation. In A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2012; 582p. [Google Scholar]
- Charlotte, L.O.; Peter, M.; Tim, N. Agriculture and climate change are reshaping insect biodiversity worldwide. Nature 2022, 605, 97–102. [Google Scholar]
- Hu, Y.L.; Tian, X.Y.; Chen, L.; Ding, N.; Li, X.F.; Gao, Y.; Shi, S.S. Analysis of fertility and population trend of experimental population of Riptortus pedestris fed on soybean pods. Chin. J. Oil Crop Sci. 2022, 44, 460–466. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Stage | Developmental Period (Days) | ||||
|---|---|---|---|---|---|
| 19 °C | 22 °C | 25 °C | 28 °C | 31 °C | |
| Egg | 17.25 ± 0.11 a | 11.03 ± 0.02 b | 8.63 ± 0.06 c | 5.93 ± 0.04 d | 5.10 ± 0.04 d |
| First instar | 3.78 ± 0.08 a | 3.04 ± 0.08 a | 2.43 ± 0.08 b | 1.93 ± 0.05 c | 1.39 ± 0.06 c |
| Second instar | 4.70 ± 0.36 a | 4.43 ± 0.20 a | 3.34 ± 0.20 a | 2.26 ± 0.25 b | 2.17 ± 0.18 b |
| Third instar | 5.79 ± 0.54 a | 5.33 ± 0.40 a | 3.51 ± 0.24 a | 3.23 ± 0.37 b | 2.67 ± 0.31 b |
| Fourth instar | 9.78 ± 0.74 a | 6.93 ± 0.58 b | 5.83 ± 0.27 b | 4.17 ± 0.38 c | 4.53 ± 0.23 c |
| Fifth instar | 16.50 ± 2.05 a | 11.67 ± 0.97 b | 8.89 ± 0.56 b | 6.90 ± 0.64 c | 6.57 ± 0.41 c |
| Egg to adult | 55.00 ± 3.66 a | 42.44 ± 1.34 b | 34.53 ± 0.67 c | 25.40 ± 0.91 d | 24.43 ± 0.98 d |
| Female longevity | 95.50 ± 24.50 a | 64.00 ± 8.33 b | 51.73 ± 5.38 c | 33.83 ± 3.25 d | 27.50 ± 4.50 e |
| Male longevity | 114.50 ± 5.50 a | 64.83 ± 8.91 b | 55.38 ± 6.43 c | 50.50 ± 4.91 d | 31.80 ± 2.44 e |
| APOP | 32.00 ± 0.09 a | 8.00 ± 0.71 b | 7.50 ± 0.31 c | 5.00 ± 0.05 d | 4.00 ± 0.34 e |
| TPOP | 80.00 ± 0.29 a | 46.00 ± 0.70 b | 43.30 ± 1.35 c | 30.00 ± 0.07 d | 24.00 ± 0.82 e |
| Fecundity | 24.00 ± 11.83 d | 45.00 ± 15.36 b | 58.00 ± 28.59 a | 30.00 ± 9.77 c | 23.00 ± 11.09 d |
| Developmental Stage | Developmental Threshold Temperature (°C) | Effective Accumulative Temperature (Degree-Days) | Coefficient of Variation (cv) |
|---|---|---|---|
| Egg | 13.31 ± 0.00 b | 95.91 ± 0.07 ef | 0.06 ± 0.00 e |
| First instar | 11.74 ± 0.21 bc | 29.78 ± 0.44 h | 0.08 ± 0.01 d |
| Second instar | 11.61 ± 0.60 bc | 40.93 ± 2.38 gh | 0.12 ± 0.06 c |
| Third instar | 10.43 ± 1.36 c | 54.83 ± 6.82 g | 0.09 ± 0.03 d |
| Fourth instar | 10.57 ± 0.27 bc | 82.20 ± 1.64 f | 0.09 ± 0.00 d |
| Fifth instar | 11.78 ± 0.23 bc | 118.82 ± 1.69 d | 0.04 ± 0.01 e |
| Nymph | 10.91 ± 0.35 bc | 260.51 ± 1.83 b | 0.12 ± 0.05 c |
| Female adult | 16.21 ± 0.13 a | 114.22 ± 2.35 de | 0.18 ± 0.02 b |
| Male adult | 17.13 ± 0.42 a | 147.35 ± 5.79 c | 0.47 ± 0.05 a |
| Generation | 13.44 ± 0.05 b | 582.04 ± 0.05 a | 0.04 ± 0.05 e |
| Temperature °C | |||||
|---|---|---|---|---|---|
| 19 | 22 | 25 | 28 | 31 | |
| Net reproductive rate (R0) | 0.63 ± 0.32 b | 1.52 ± 0.77 ab | 2.36 ± 1.69 a | 0.79 ± 0.40 b | 0.60 ± 0.31 b |
| Mean generation time T (d) | 86.76 ± 0.04 a | 53.55 ± 0.09 b | 56.44 ± 2.04 b | 41.34 ± 9.86 c | 27.50 ± 0.02 d |
| Intrinsic rate of increase r (per day) | −0.01 ± 0.01 b | 0.01 ± 0.01 a | 0.01 ± 0.02 a | −0.01 ± 0.01 b | −0.02 ± 0.01 b |
| Finite rate of increase λ (per day) | 0.99 ± 0.01 b | 1.01 ± 0.01 a | 1.01 ± 0.02 a | 0.99 ± 0.01 b | 0.98 ± 0.02 b |
| Number of Individuals Entering the Next Stage | |||||
|---|---|---|---|---|---|
| Developmental Stage | 19 °C | 22 °C | 25 °C | 28 °C | 31 °C |
| Egg | 100.0 ± 0.00 a | 100.0 ± 0.00 a | 100.0 ± 0.00 a | 100.0 ± 0.00 a | 100.0 ± 0.00 a |
| First instar | 82.35 ± 0.61 c | 90.32 ± 0.64 b | 95.45 ± 0.59 a | 92.31 ± 1.12 b | 83.33 ± 0.78 c |
| Second instar | 49.41 ± 6.38 c | 75.27 ± 6.59 a | 81.13 ± 4.08 a | 61.54 ± 1.21 b | 79.16 ± 7.56 a |
| Third instar | 41.18 ± 7.20 c | 48.17 ± 1.91 bc | 60.45 ± 3.83 a | 47.69 ± 6.61 bc | 58.33 ± 2.43 ab |
| Fourth instar | 26.08 ± 3.09 c | 34.62 ± 2.51 b | 52.50 ± 3.44 a | 32.31 ± 5.78 b | 34.72 ± 2.16 b |
| Fifth instar | 12.35 ± 0.51 d | 21.07 ± 6.86 c | 47.73 ± 8.65 a | 27.69 ± 8.06 b | 23.61 ± 9.19 bc |
| Adult | 5.49 ± 1.87 d | 13.55 ± 2.75 bc | 30.23 ± 0.92 a | 15.39 ± 3.57 b | 9.72 ± 0.94 cd |
| Female ratio | 0.50 ± 0.61 a | 0.33 ± 0.73 b | 0.58 ± 1.01 a | 0.6 ± 0.54 a | 0.29 ± 0.37 b |
| Number of eggs laid per female | 24.00 ± 11.83 d | 45.00 ± 15.36 b | 58.00 ± 28.59 a | 30.00 ± 9.77 c | 23.00 ± 11.09 e |
| Number of eggs expected in the following generation | 65.88 ± 3.65 c | 259.35 ± 18.73 b | 789.00 ± 23.78 a | 277.02 ± 19.19 b | 64.83 ± 6.38 c |
| Population trend index (I) | 0.66 ± 0.72 c | 2.59 ± 1.23 b | 7.89 ± 2.34 a | 2.77 ± 0.50 b | 0.65 ± 0.23 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, X.-Y.; Gao, Y.; Ali, M.Y.; Li, X.-H.; Hu, Y.-L.; Li, W.-B.; Wang, Z.-J.; Shi, S.-S.; Zhang, J.-P. Impact of Temperature on Age–Stage, Two-Sex Life Table Analysis of a Chinese Population of Bean Bug, Riptortus pedestris (Hemiptera: Alydidae). Agriculture 2022, 12, 1505. https://doi.org/10.3390/agriculture12091505
Tian X-Y, Gao Y, Ali MY, Li X-H, Hu Y-L, Li W-B, Wang Z-J, Shi S-S, Zhang J-P. Impact of Temperature on Age–Stage, Two-Sex Life Table Analysis of a Chinese Population of Bean Bug, Riptortus pedestris (Hemiptera: Alydidae). Agriculture. 2022; 12(9):1505. https://doi.org/10.3390/agriculture12091505
Chicago/Turabian StyleTian, Xin-Yue, Yu Gao, Muhammad Yasir Ali, Xiao-Hua Li, Ying-Lu Hu, Wen-Bo Li, Zi-Jie Wang, Shu-Sen Shi, and Jin-Ping Zhang. 2022. "Impact of Temperature on Age–Stage, Two-Sex Life Table Analysis of a Chinese Population of Bean Bug, Riptortus pedestris (Hemiptera: Alydidae)" Agriculture 12, no. 9: 1505. https://doi.org/10.3390/agriculture12091505