


Kinetics of Phosphorus Uptake through Roots of Habanero Pepper (Capsicum chinense Jacq. cv. Mayapán)

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions
2.2. Experimental Procedure
3. Results
3.1. Characteristics of the Habanero Pepper Seedlings
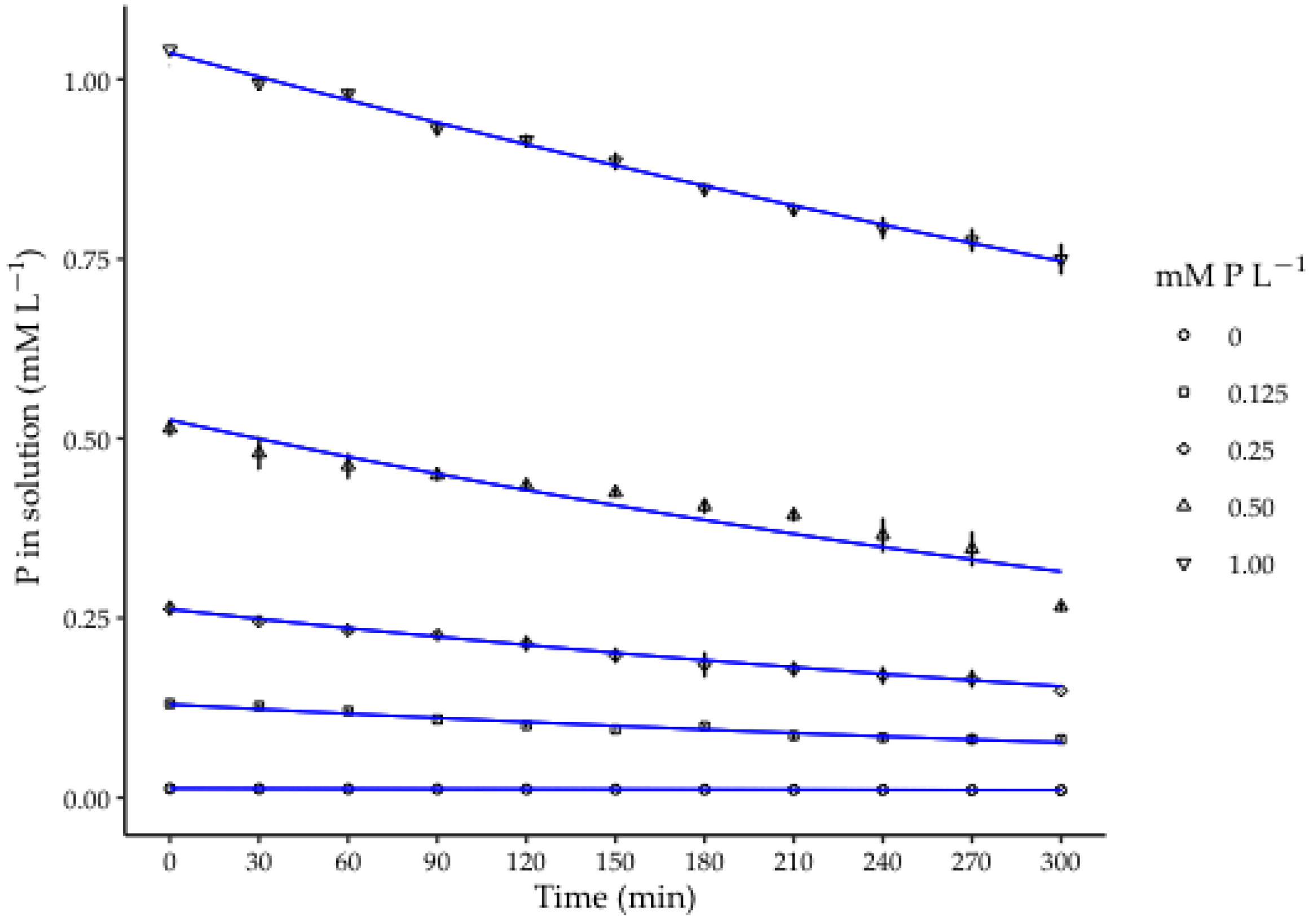
3.2. Curves of P Depletion by Habanero Pepper Roots
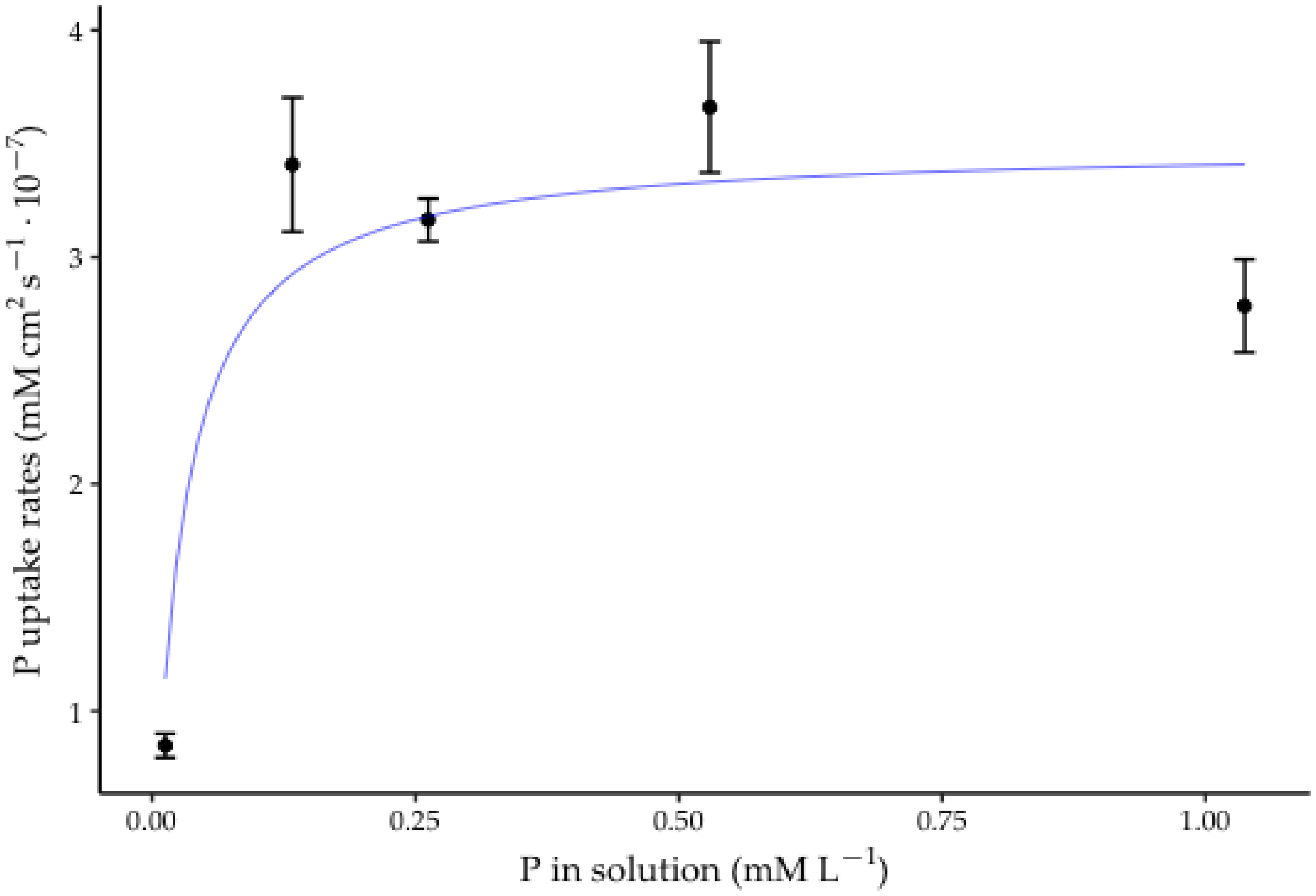
3.3. P-Uptake Rates by Roots
3.4. Parameters of the P-Uptake Kinetic of Habanero Pepper Roots
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ajmera, I.; Hodgman, T.C.; Lu, C. An Integrative Systems Perspective on Plant Phosphate Research. Genes 2019, 10, 139. [Google Scholar] [CrossRef]
- Alewell, C.; Ringeval, B.; Ballabio, C.; Robinson, D.A.; Panagos, P.; Borrelli, P. Global Phosphorus Shortage Will Be Aggravated by Soil Erosion. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Mitra, G.N. Regulation of Nutrient Uptake by Plants: A Biochemical and Molecular Approach; Springer India: New Delhi, India, 2015; pp. 1–195. [Google Scholar]
- Bai, Q.; Shen, Y.; Huang, Y. Advances in Mineral Nutrition Transport and Signal Transduction in Rosaceae Fruit Quality and Postharvest Storage. Front. Plant Sci. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 3–651. [Google Scholar]
- Zhang, F.; Sun, Y.; Pei, W.; Jain, A.; Sun, R.; Cao, Y.; Wu, X.; Jiang, T.; Zhang, L.; Fan, X.; et al. Involvement of OsPht1;4 in Phosphate Acquisition and Mobilization Facilitates Embryo Development in Rice. Plant J. 2015, 82, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lv, S.; Jiang, P.; Li, Y. Roles, Regulation, and Agricultural Application of Plant Phosphate Transporters. Front Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kovar, J.L.; Claassen, N. Soil-Root Interactions and Phosphorus Nutrition of Plants. In Phosphorus: Agriculture and the Environment; American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America: Madison, WI, USA, 2005; pp. 379–414. [Google Scholar]
- Yao, F.; Sun, J.; Tang, C.; Ni, W. Kinetics of Ammonium, Nitrate and Phosphate Uptake by Candidate Plants Used in Constructed Wetlands. Procedia Environ. Sci. 2011, 10, 1854–1861. [Google Scholar] [CrossRef]
- Poirier, Y.; Bucher, M. Phosphate Transport and Homeostasis in Arabidopsis. Arabidopsis Book 2002, 1, 1–35. [Google Scholar] [CrossRef]
- Claassen, N.; Barber, S.A. A Method for Characterizing the Relation between Nutrient Concentration and Flux into Roots of Intact Plants. Plant Physiol. 1974, 54, 564–568. [Google Scholar] [CrossRef]
- le Deunff, E.; Malagoli, P. An Updated Model for Nitrate Uptake Modelling in Plants. I. Functional Component: Cross-Combination of Flow–Force Interpretation of Nitrate Uptake Isotherms, and Environmental and in Planta Regulation of Nitrate Influx. Ann. Bot. 2014, 113, 991–1005. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.S.; Oki, Y.; Adachi, T. Mobilization and Acquisition of Sparingly Soluble P-Sources by Brassica Cultivars under P-Starved Environment II. Rhizospheric PH Changes, Redesigned Root Architecture and Pi-Uptake Kinetics. J. Integr. Plant Biol. 2009, 51, 1024–1039. [Google Scholar] [CrossRef]
- Griffiths, M.; York, L.M. Targeting Root Ion Uptake Kinetics to Increase Plant Productivity and Nutrient Use Efficiency1[Open]. Plant Physiol. 2020, 182, 1854–1868. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Piñeros, M.A.; Wang, Z.; Wang, W.; Li, C.; Wu, Z.; Kochian, L.V.; Wu, P. Phosphate Transporters OsPHT1;9 and OsPHT1;10 are Involved in Phosphate Uptake in Rice. Plant Cell Environ. 2014, 37, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, N.H.; Andersen, F.O.; Jensen, H.S. Phosphate Uptake Kinetics for Four Species of Submerged Freshwater Macrophytes Measured by a 33P Phosphate Radioisotope Technique. Aquat. Bot. 2016, 128, 58–67. [Google Scholar] [CrossRef]
- Kavka, M.; Polle, A. Phosphate Uptake Kinetics and Tissue-Specific Transporter Expression Profiles in Poplar (Populus × canescens) at Different Phosphorus Availabilities. BMC Plant Biol. 2016, 16, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rae, A.L.; Cybinski, D.H.; Jarmey, J.M.; Smith, F.W. Characterization of Two Phosphate Transporters from Barley; Evidence for Diverse Function and Kinetic Properties among Members of the Pht1 Family. Plant Mol. Biol. 2003, 53, 27–36. [Google Scholar] [CrossRef]
- Kelly, J.M.; Kelly, J.K. Phosphorus and Potassium Uptake Kinetics in Red Maple Seedlings. Forest Sci. 2001, 47, 397–402. [Google Scholar] [CrossRef]
- Wang, K.; Hu, Q.; Wei, Y.; Yin, H.; Sun, C.; Liu, G. Uptake Kinetics of NH+4, NO3− and H2PO−4 by Typha orientalis, Acorus calamus L., Lythrum salicaria L., Sagittaria trifolia L. and Alisma plantago-aquatica Linn. Sustainability 2021, 13, 434. [Google Scholar] [CrossRef]
- Borges-Gómez, L.; Chuc-Puc, J.; Escamilla-Bencomo, A.; Medina-Lara, F. Kinetics of Potassium Absorption by Habanero Chili (Capsicum chinense Jacq.) Roots. Agrociencia 2006, 40, 431–440. [Google Scholar]
- Official Gazette of the Federation [DOF] General Declaration of the Denomination of Origin Protection Habanero Pepper from Yucatán Federación. Available online: https://dof.gob.mx/nota_detalle.php?codigo=5063633&fecha=10/10/2008#gsc.tab=0 (accessed on 24 August 2022).
- Aviles-Baeza, W.I.; Lozano-Contreras, M.G.; Ramírez-Silva, J.H. Evaluation of Habanero Pepper (Capsicum chinense Jacq.) Varieties under Shade House Conditions in Yucatan, Mexico. OAlib 2021, 8, 1–10. [Google Scholar] [CrossRef]
- Meneses-Lazo, R.E.; Garruña, R. The Habanero Pepper (Capsicum chinense Jacq.) as a Study Plant Model in México. Trop. Subtrop. Agroecosystems 2020, 23, 1–17. [Google Scholar]
- Steiner, A.A. A Universal Method for Preparing Nutrient Solutions of a Certain Desired Composition. Plant Soil. 1961, 15, 134–154. [Google Scholar] [CrossRef]
- Steiner, A.A. The Universal Nutrient Solution. In Proceedings of the 6th International Congress on Soilless (ISOSC), Lunteren, The Netherlands, 29 April–5 May 1984; pp. 633–650. [Google Scholar]
- Burton, J.D.; Riley, J.P. Determination of Soluble Phosphate, and Total Phosphorus in Sea-Water and of Total Phosphorus in Marine Muds. Mikrochim. Acta 1956, 44, 1350–1365. [Google Scholar] [CrossRef]
- Pace, J.; Lee, N.; Naik, H.S.; Ganapathysubramanian, B.; Lübberstedt, T. Analysis of Maize (Zea mays L.) Seedling Roots with the High-Throughput Image Analysis Tool ARIA (Automatic Root Image Analysis). PLoS ONE 2014, 9, e108255. [Google Scholar] [CrossRef]
- Pierret, A.; Gonkhamdee, S.; Jourdan, C.; Maeght, J.L. IJ_Rhizo: An Open-Source Software to Measure Scanned Images of Root Samples. Plant Soil 2013, 373, 531–539. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 20 April 2022).
- Shen, Q.; Wen, Z.; Dong, Y.; Li, H.; Miao, Y.; Shen, J. The Responses of Root Morphology and Phosphorus-Mobilizing Exudations in Wheat to Increasing Shoot Phosphorus Concentration. AoB Plants 2018, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sattelmacher, B.; Klotz, F.; Marschner, H. Influence of the Nitrogen Level on Root Growth and Morphology of Two Potato Varieties Differing in Nitrogen Acquisition. Plant Soil 1990, 123, 131–137. [Google Scholar] [CrossRef]
- Chaudhary, M.I.; Adu-Gyamfi, J.J.; Saneoka, H.; Nguyen, N.T.; Suwa, R.; Kanai, S.; El-Shemy, H.A.; Lightfoot, D.A.; Fujita, K. The Effect of Phosphorus Deficiency on Nutrient Uptake, Nitrogen Fixation and Photosynthetic Rate in Mashbean, Mungbean and Soybean. Acta Physiol. Plant 2008, 30, 537–544. [Google Scholar] [CrossRef]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The Impacts of Phosphorus Deficiency on the Photosynthetic Electron Transport Chain1[OPEN]. Plant Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef]
- Yan, X.; Wei, Z.; Hong, Q.; Lu, Z.; Wu, J. Phosphorus Fractions and Sorption Characteristics in a Subtropical Paddy Soil as Influenced by Fertilizer Sources. Geoderma 2017, 295, 80–85. [Google Scholar] [CrossRef]
- Nievola, C.C.; Carvalho, C.P.; Carvalho, V.; Rodrigues, E. Rapid Responses of Plants to Temperature Changes. Temperature 2017, 4, 371–405. [Google Scholar] [CrossRef]
- Jungk, A.; Asher, C.J.; Edwards, D.G.; Meyer, D. Influence of Phosphate Status on Phosphate Uptake Kinetics of Maize (Zea mays) and Soybean (Glycine max). Plant Soil 1990, 124, 175–182. [Google Scholar] [CrossRef]
- Brix, H.; Lorenzen, B.; Mendelssohn, I.A.; McKee, K.L.; Miao, S.L. Can Differences in Phosphorus Uptake Kinetics Explain the Distribution of Cattail and Sawgrass in the Florida Everglades? BMC Plant Biol. 2010, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechtaoui, N.; Rabiu, M.K.; Raklami, A.; Oufdou, K.; Hafidi, M.; Jemo, M. Phosphate-Dependent Regulation of Growth and Stresses Management in Plants. Front Plant Sci. 2021, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H. Enhanced Phosphorus Fertilizer Use Efficiency with Microorganisms. In Nutrient Dynamics for Sustainable Crop Production; Meena, R.S., Ed.; Springer: Singapore, 2020; pp. 215–245. [Google Scholar]
- Wright, M.R. An Introduction to Chemical Kinetics Library of Congress Cataloging-in-Publication Data; Wiley Publishing, Inc.: Chichester, UK; London, UK, 2004; pp. 1–429. [Google Scholar]
- Rausch, C.; Daram, P.; Brunner, S.; Jansa, J.; Laloi, M.; Leggewie, G.; Amrhein, N.; Bucher, M. A Phosphate Transporter Expressed in Arbuscule-Containing Cells in Potato. Nature 2001, 414, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Ren, H.; Gu, M.; Zhao, J.; Sun, S.; Zhang, X.; Chen, J.; Wu, P.; Xu, G. The Phosphate Transporter Gene OsPht1;8 Is Involved in Phosphate Homeostasis in Rice. Plant Physiol 2011, 156, 1164–1175. [Google Scholar] [CrossRef] [PubMed]
- Furihata, T.; Suzuki, M.; Sakurai, H. Kinetic Characterization of Two Phosphate Uptake Systems with Different Affinities in Suspension-Cultured Catharanthus Roseus Protoplasts. Plant Cell Physiol. 1992, 33, 1151–1157. [Google Scholar] [CrossRef]
- Daram, P.; Brunner, S.; Rausch, C.; Steiner, C.; Amrhein, N.; Bucher, M. Pht2;1 Encodes a Low-Affinity Phosphate Transporter from Arabidopsis. Plant Cell 1999, 11, 2153–2166. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Treatment mM L−1 | NO3− | SO42− | H2PO4− | K+ | Ca2+ | Mg2+ |
|---|---|---|---|---|---|---|
| mM L−1 | ||||||
| 0.01 | 15 | 2 | 0 | 7 | 4 | 2 |
| 0.125 | 15 | 2 | 0.125 | 7.125 | 4 | 2 |
| 0.25 | 15 | 2 | 0.25 | 7.25 | 4 | 2 |
| 0.50 | 15 | 2 | 0.5 | 7.5 | 4 | 2 |
| 1.00 | 15 | 2 | 1 | 8 | 4 | 2 |
| Parameter | Mean | Lower Limit | Upper Limit |
|---|---|---|---|
| Root length (cm) | 250.33 | 233.44 | 267.23 |
| Root surface area (cm2) | 7.06 | 6.25 | 7.87 |
| Root diameter (mm) | 1.05 | 0.99 | 1.13 |
| Root dry weight (mg plant−1) | 38.44 | 36.03 | 41.18 |
| Shoot dry weight (mg plant−1) | 261.95 | 245.49 | 280.2 |
| P concentration (mg plant−1) | 1.39 | 1.28 | 1.50 |
| Treatment | Cmax | Cmin | Model | R2 |
|---|---|---|---|---|
| (mM P L−1) | ||||
| 0 | 0.013 | 0.010 | y = 0.013e−0.00065x | 0.994 |
| 0.125 | 0.131 | 0.081 | y = 0.128e−0.001752x | 0.956 |
| 0.25 | 0.264 | 0.149 | y = 0.262e−0.00182x | 0.994 |
| 0.50 | 0.514 | 0.266 | y = 0.53e−0.00171x | 0.899 |
| 1.00 | 1.04 | 0.75 | y = 1.04e−0.00109x | 0.996 |
| Initial Concentration of P (mM L−1) | P-Uptake Rate (mM cm2 s−1) | VC (%) |
|---|---|---|
| 0.01 | 8.47 × 10−8 b | 2.65 |
| 0.13 | 3.41 × 10−7 a | 11.64 |
| 0.26 | 3.16 × 10−7 a | 2.97 |
| 0.53 | 3.66 × 10−7 a | 6.93 |
| 1.04 | 2.78 × 10−7 a | 7.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yam-Chimal, C.; Ramírez-y-Avilés, L.; Navarro-Alberto, J.; Solorio-Sánchez, F.; Villanueva-Couoh, E.; Noh-Medina, J. Kinetics of Phosphorus Uptake through Roots of Habanero Pepper (Capsicum chinense Jacq. cv. Mayapán). Agriculture 2022, 12, 1389. https://doi.org/10.3390/agriculture12091389
Yam-Chimal C, Ramírez-y-Avilés L, Navarro-Alberto J, Solorio-Sánchez F, Villanueva-Couoh E, Noh-Medina J. Kinetics of Phosphorus Uptake through Roots of Habanero Pepper (Capsicum chinense Jacq. cv. Mayapán). Agriculture. 2022; 12(9):1389. https://doi.org/10.3390/agriculture12091389
Chicago/Turabian StyleYam-Chimal, Carmen, Luis Ramírez-y-Avilés, Jorge Navarro-Alberto, Francisco Solorio-Sánchez, Eduardo Villanueva-Couoh, and José Noh-Medina. 2022. "Kinetics of Phosphorus Uptake through Roots of Habanero Pepper (Capsicum chinense Jacq. cv. Mayapán)" Agriculture 12, no. 9: 1389. https://doi.org/10.3390/agriculture12091389