
Nodule Formation and Nitrogen Use Efficiency Are Important for Soybean to Adapt to Water and P Deficit Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Treatments
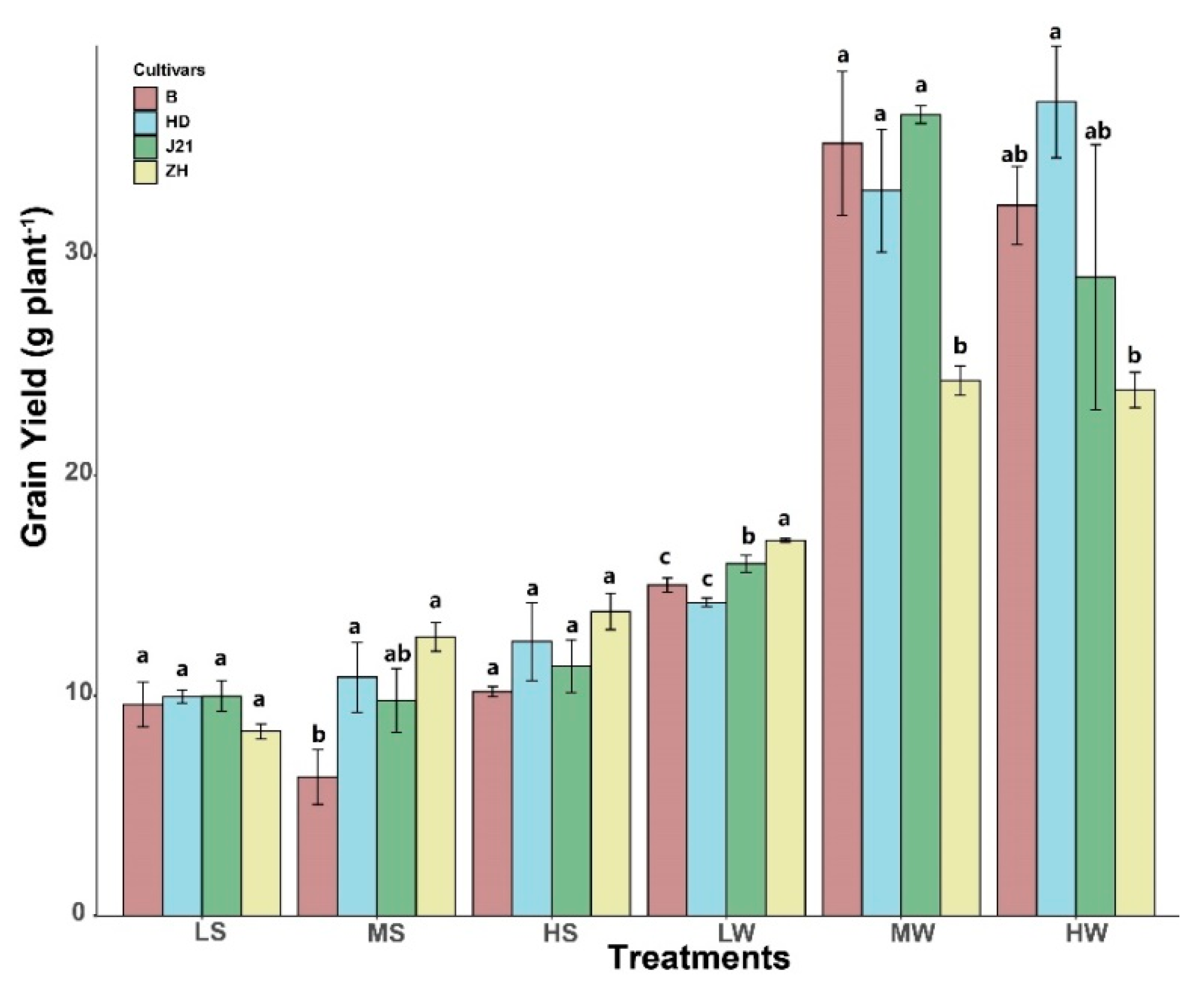
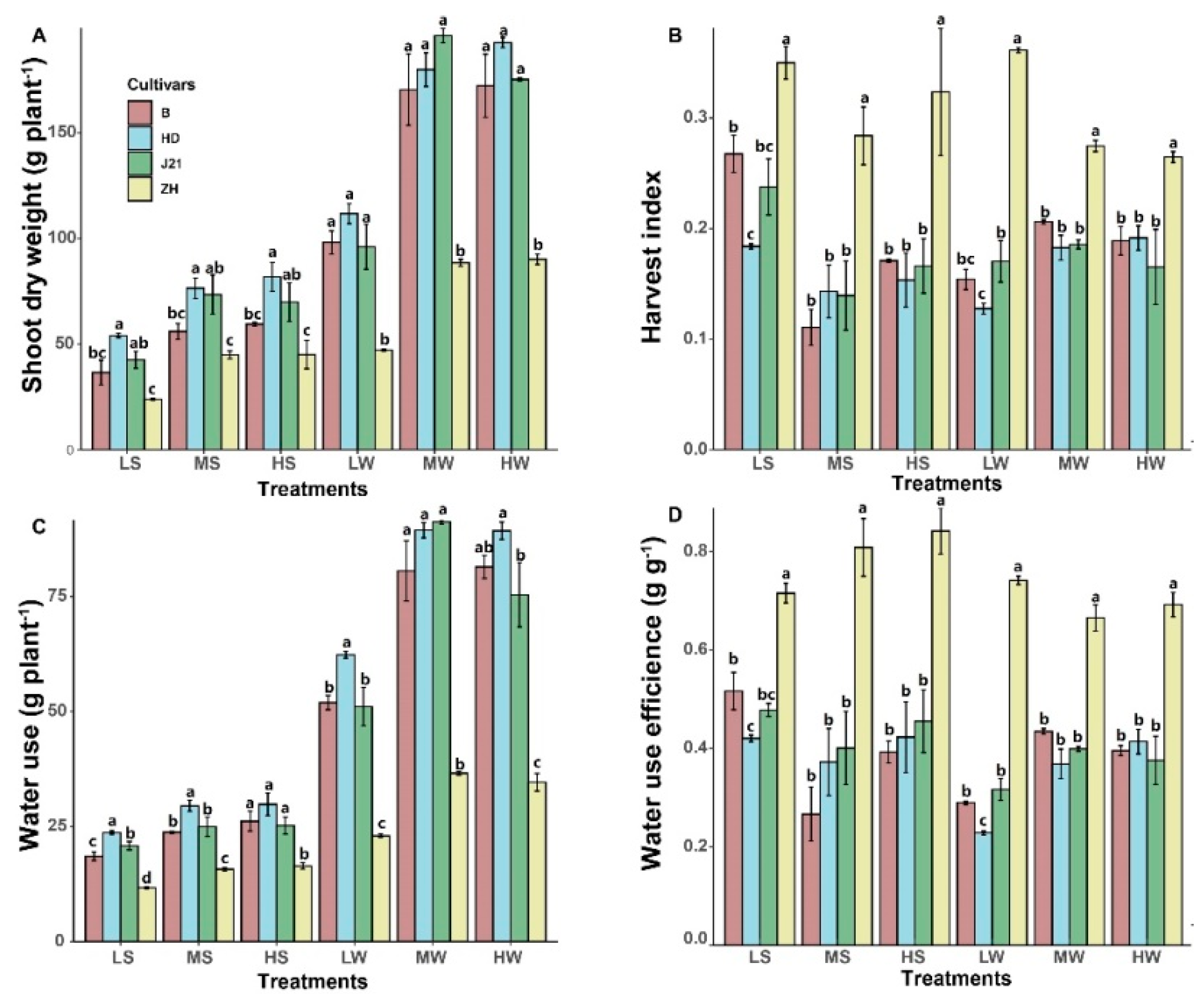
2.2. Yield Determination
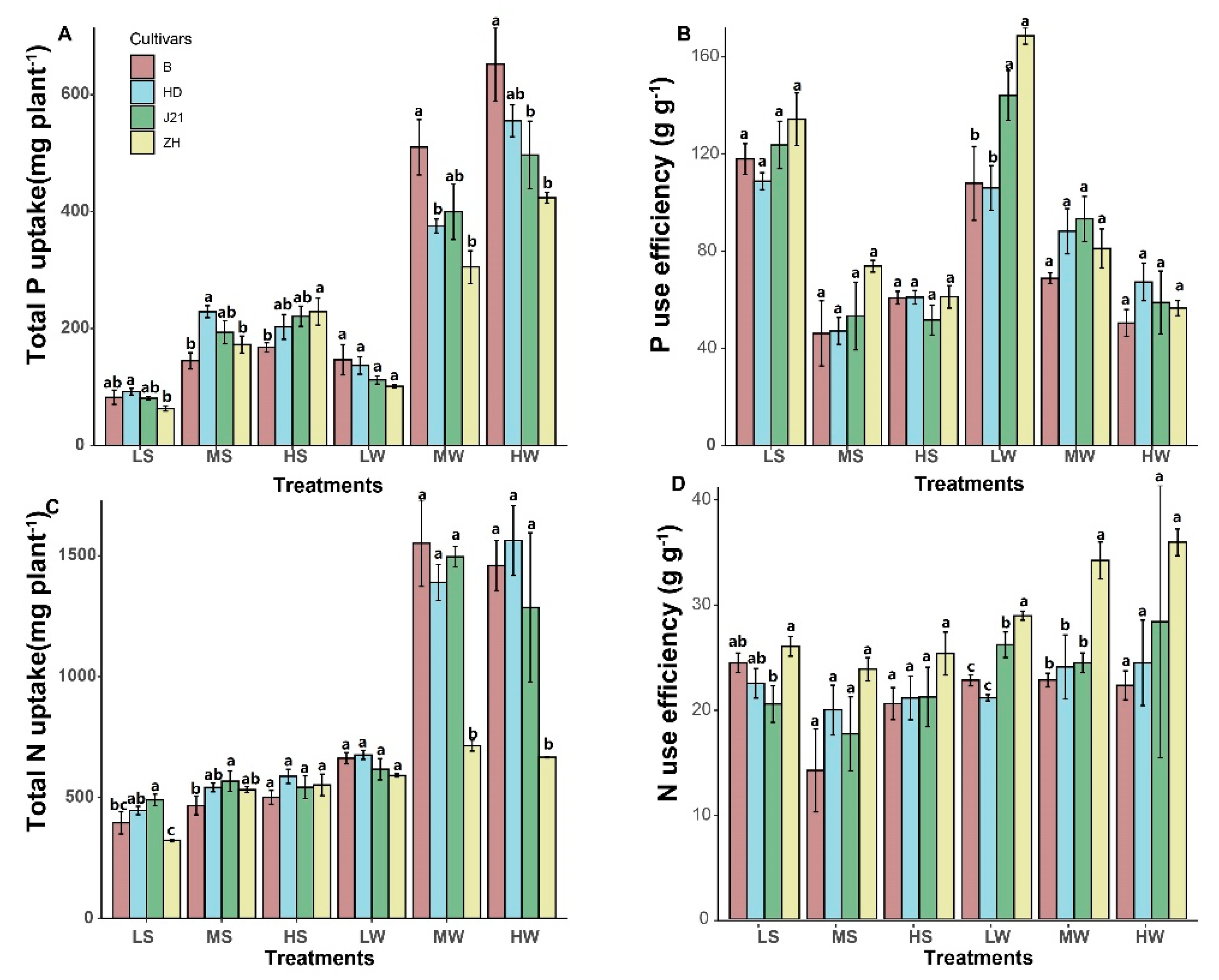
2.3. P and N Use Efficiency
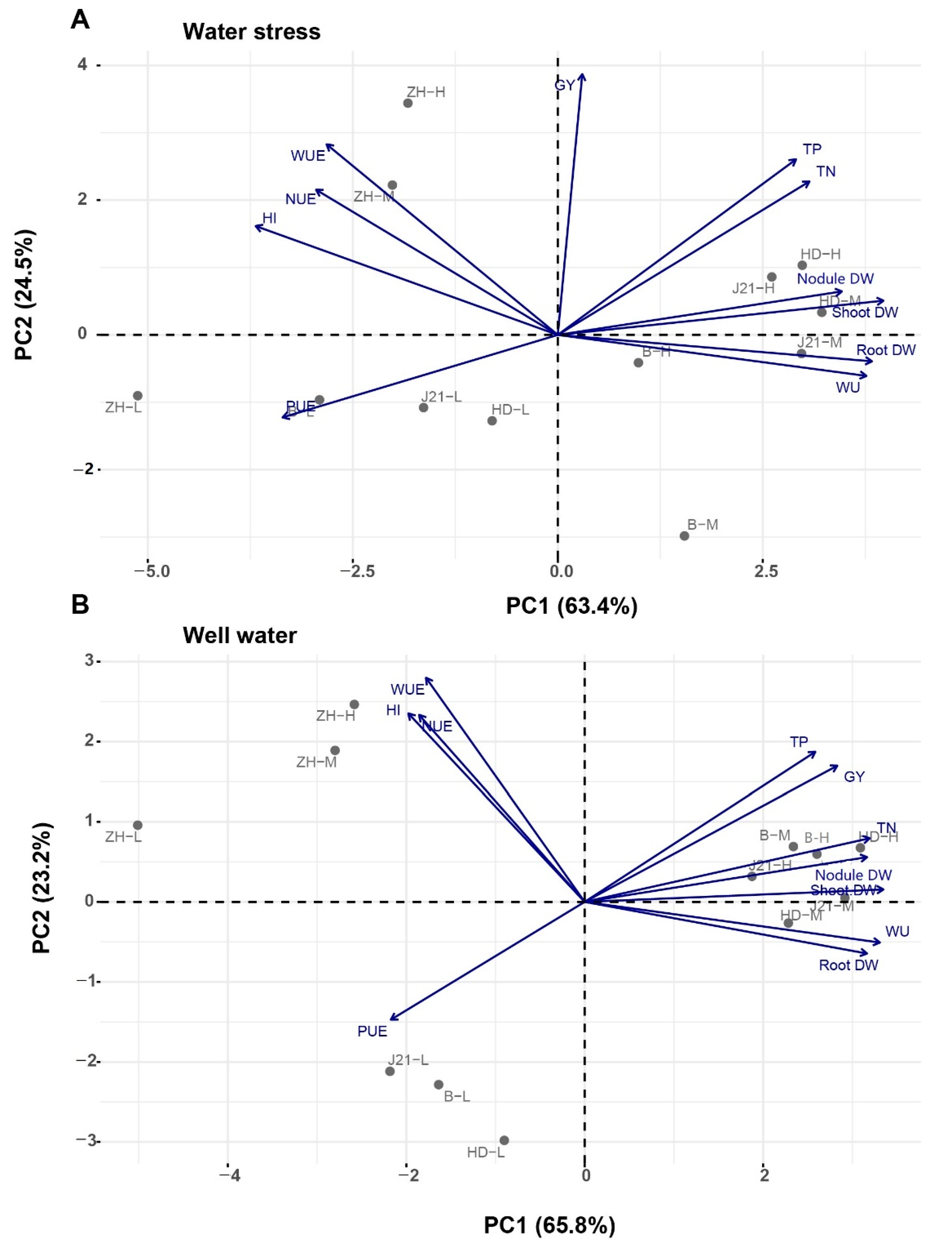
2.4. Data Analysis
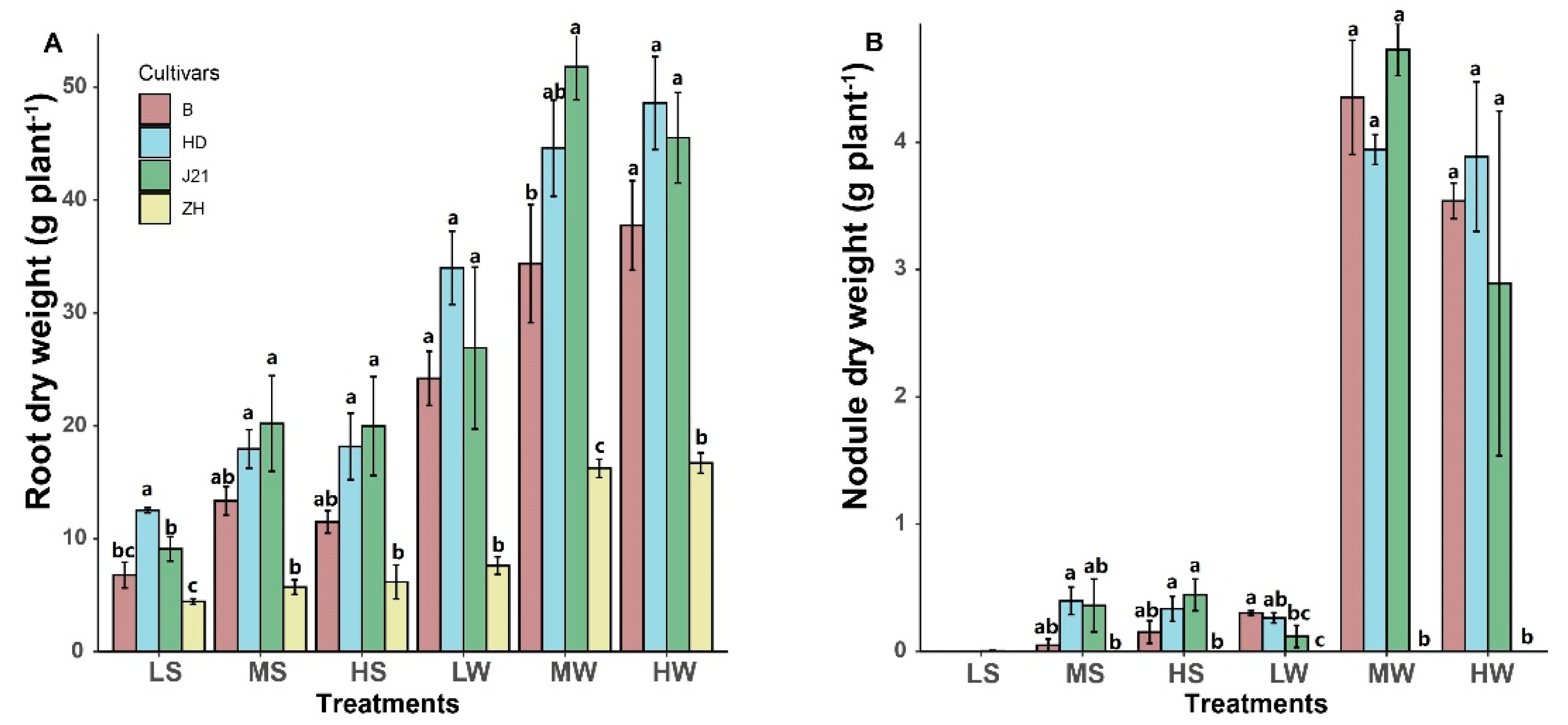
3. Results
4. Discussion
4.1. Interaction of P Supply and Water Treatment on Soybean Yield Performance
4.2. The Relationships between the Nodulation in Soybean and P and N Accumulation and Yield Performance under Different Water and P Levels
4.3. How NUE and PUE Influences Soybean Yield Performance?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, J.; Du, Y.L.; Wang, T.; Turner, N.C.; Xi, Y.; Li, F.M. Old and New Cultivars of Soya Bean (Glycine max L.) Subjected to Soil Drying Differ in Abscisic Acid Accumulation, Water Relations Characteristics and Yield. J. Agron. Crop Sci. 2016, 202, 372–383. [Google Scholar] [CrossRef]
- Jin, Y.; He, J.; Turner, N.C.; Du, Y.L.; Li, F.M. Water-Conserving and Biomass-Allocation Traits Are Associated with Higher Yields in Modern Cultivars Compared to Landraces of Soybean [Glycine Max (L.) Merr.] in Rainfed Water-Limited Environments. Environ. Exp. Bot. 2019, 168, 103883. [Google Scholar] [CrossRef]
- Feng, Y.Y.; Richards, A.R.; Jin, Y.; Siddique, K.H.M.; Li, F.M.; He, J. Yield and Water-Use Related Traits in Landrace and New Soybean Cultivars in Arid and Semi-Arid Areas of China. Field Crop Res. 2022, 283, 108559. [Google Scholar] [CrossRef]
- Yang, M.H.; Jahufer, M.Z.Z.; He, J.; Dong, R.; Hofmann, R.; Siddique, K.H.M.; Li, F.M. Effect of Traditional Soybean Breeding on Water Use Strategy in Arid and Semi-Arid Areas. Eur. J. Agron. 2020, 120, 126128. [Google Scholar] [CrossRef]
- He, J.; Du, Y.L.; Wang, T.; Turner, N.C.; Yang, R.P.; Siddique, K.H.M.; Li, F.M. Genotypic Variation in Yield, Yield Components, Root Morphology and Architecture, in Soybean in Relation to Water and Phosphorus Supply. Front. Plant Sci. 2017, 8, 1499. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.Y.; He, J.; Jin, Y.; Li, F.M. High Phosphorus Acquisition and Allocation Strategy Is Associated with Soybean Seed Yield under Water- and P-Limited Conditions. Agronomy 2021, 11, 574. [Google Scholar] [CrossRef]
- Mpelasoka, F.; Hennessy, K.; Jones, R.; Bates, B. Comparison of Suitable Drought Indices for Climate Change Impacts Assessment over Australia Towards Resource Management. Int. J. Climatol. 2008, 28, 1283–1292. [Google Scholar] [CrossRef]
- Turner, N.C.; Molyneux, N.; Yang, S.; Xiong, Y.C.; Siddique, K.H.M. Climate Change in South-West Australia and North-West China: Challenges and Opportunities for Crop Production. Crop Pasture Sci. 2011, 62, 445–456. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.O.; White, S. The Story of Phosphorus: Global Food Security and Food for Thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Gilbert, N. Environment: The Disappearing Nutrient. Nature 2009, 461, 716–718. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Jin, Y.; Turner, N.C.; Chen, Z.; Liu, H.Y.; Wang, X.L.; Siddique, K.H.M.; Li, F.M. Phosphorus Application Increases Root Growth, Improves Daily Water Use During the Reproductive Stage, and Increases Grain Yield in Soybean Subjected to Water Shortage. Environ. Exp. Bot. 2019, 166, 103816. [Google Scholar] [CrossRef]
- Lynch, J.P. Root Phenotypes for Improved Nutrient Capture: An Underexploited Opportunity for Global Agriculture. New Phytol. 2019, 223, 548–564. [Google Scholar] [CrossRef] [PubMed]
- Rosolem, A.; Thiago, B.; Patrícia, P.; Laudelino, V.; Juliano, C. The Joint Application of Phosphorus and Ammonium Enhances Soybean Root Growth and P Uptake. Agriculture 2022, 12, 880. [Google Scholar] [CrossRef]
- He, J.; Jin, Y.; Siddique, K.H.M.; Li, F.M. Trade-Off between Root Efficiency and Root Size Is Associated with Yield Performance of Soybean under Different Water and Phosphorus Levels. Agriculture 2021, 11, 481. [Google Scholar] [CrossRef]
- Donahue, J.M.; Bai, H.; Almtarfi, H.; Zakeri, H.; Fritschi, F.B. The Quantity of Nitrogen Derived from Symbiotic N Fixation but Not the Relative Contribution of N Fixation to Total N Uptake Increased with Breeding for Greater Soybean Yields. Field Crop Res. 2020, 259, 107945. [Google Scholar] [CrossRef]
- Wang, X.; Gao, Y.; Zhang, H.; Shao, Z.; Sun, B.; Gao, Q. Enhancement of Rhizosphere Citric Acid and Decrease of NO3−/NH4+ Ratio by Root Interactions Facilitate N Fixation and Transfer. Plant Soil 2020, 447, 169–182. [Google Scholar] [CrossRef]
- Hidaka, A.; Kitayama, K. Divergent Patterns of Photosynthetic Phosphorus-Use Efficiency Versus Nitrogen-Use Efficiency of Tree Leaves Along Nutrient-Availability Gradients. J. Ecol. 2009, 97, 984–991. [Google Scholar] [CrossRef]
- Haefele, S.M.; Jabbar, S.M.A.; Siopongco, J.D.L.C.; Tirol-Padre, A.; Amarante, S.T.; Sta Cruz, P.C.; Cosico, W.C. Nitrogen Use Efficiency in Selected Rice (Oryza Sativa L.) Genotypes under Different Water Regimes and Nitrogen Levels. Field Crop Res. 2008, 107, 137–146. [Google Scholar] [CrossRef]
- Araújo, S.S.; Beebe, S.; Crespi, M.; Delbreil, B.; González, E.M.; Gruber, V.; Lejeune-Henaut, I.; Link, W.; Monteros, M.J.; Prats, E.; et al. Abiotic Stress Responses in Legumes: Strategies Used To cope with Environmental Challenges. Crit. Rev. Plant Sci. 2015, 34, 237–280. [Google Scholar] [CrossRef]
- An, N.; Wei, W.; Qiao, L.; Zhang, F.; Christie, P.; Jiang, R.; Dobermann, A.; Goulding, K.W.T.; Fan, J.; Fan, M. Agronomic and Environmental Causes of Yield and Nitrogen Use Efficiency Gaps in Chinese Rice Farming Systems. Eur. J. Agron. 2018, 93, 40–49. [Google Scholar] [CrossRef]
- Wu, Z.; Luo, J.; Han, Y.; Hua, Y.; Guan, C.; Zhang, Z. Low Nitrogen Enhances Nitrogen Use Efficiency by Triggering NO3– Uptake and Its Long-Distance Translocation. J. Agric. Food Chem. 2019, 67, 6736–6747. [Google Scholar] [CrossRef] [PubMed]
- Muurinen, S.; Slafer, G.A.; Peltonen-Sainio, P. Breeding Effects on Nitrogen Use Efficiency of Spring Cereals under Northern Conditions. Crop Sci. 2006, 46, 561–568. [Google Scholar] [CrossRef]
- Bingham, I.J.; Karley, A.J.; White, P.J.; Thomas, W.T.B.; Russell, J.R. Analysis of Improvements in Nitrogen Use Efficiency Associated with 75 Years of Spring Barley Breeding. Eur. J. Agron. 2012, 42, 49–58. [Google Scholar] [CrossRef]
- Sadras, V.O.; Lawson, C. Nitrogen and Water-Use Efficiency of Australian Wheat Varieties Released between 1958 and 2007. Eur. J. Agron. 2013, 46, 34–41. [Google Scholar] [CrossRef]
- Meng, T.Y.; Zhang, X.B.; Ge, J.L.; Chen, X.; Zhu, G.L.; Chen, Y.L.; Zhou, G.S.; Wei, H.H.; Dai, Q.G. Improvements in Grain Yield and Nutrient Utilization Efficiency of Japonica Inbred Rice Released since the 1980s in Eastern China. Field Crop Res. 2022, 277, 108427. [Google Scholar] [CrossRef]
- Ebmeyer, H.; Hoffmann, C.M. Efficiency of Nitrogen Uptake and Utilization in Sugar Beet Genotypes. Field Crop Res. 2021, 274, 108334. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A Modified Single Solution Method for the Determination of Phosphate in Natural Waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Jin, J.; Wang, G.; Liu, X.; Pan, X.; Herbert, S.J.; Tang, C. Interaction between Phosphorus Nutrition and Drought on Grain Yield, and Assimilation of Phosphorus and Nitrogen in Two Soybean Cultivars Differing in Protein Concentration in Grains. J. Plant Nutr. 2006, 29, 1433–1449. [Google Scholar] [CrossRef]
- Gupta, A.; Andrés, R.; Ana, I.C. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Flexas, J.; Díaz-Espejo, A.; Conesa, M.A.; Coopman, R.E.; Douthe, C.; Gago, J.; Gallé, A.; Galmés, J.; Medrano, H.; Ribas-Carbo, M.; et al. Mesophyll Conductance to Co2 and Rubisco as Targets for Improving Intrinsic Water Use Efficiency in C3 Plants. Plant Cell Environ. 2016, 39, 965–982. [Google Scholar] [CrossRef]
- King, C.A.; Purcell, L.C. Soybean Nodule Size and Relationship to Nitrogen Fixation Response to Water Deficit. Crop Sci. 2001, 41, 1099–1107. [Google Scholar] [CrossRef]
- Dokwal, D.; Romsdahl, T.B.; Kunz, D.A.; Alonso, A.P.; Dickstein, R. Phosphorus Deprivation Affects Composition and Spatial Distribution of Membrane Lipids in Legume Nodules. Plant Physiol. 2021, 185, 1847–1859. [Google Scholar] [CrossRef] [PubMed]
- Teng, W.; He, X.; Tong, Y.P. Transgenic Approaches for Improving Use Efficiency of Nitrogen, Phosphorus and Potassium in Crops. J. Integr. Agric. 2017, 16, 2657–2673. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Messina, C.D.; Beatty, A.; Samples, M. Assessment across the United States of the Benefits of Altered Soybean Drought Traits. Agron. J. 2010, 102, 475–482. [Google Scholar] [CrossRef]
- He, J.; Du, Y.L.; Wang, T.; Turner, N.C.; Yang, R.P.; Jin, Y.; Xi, Y.; Zhang, C.; Cui, T.; Fang, X.W.; et al. Conserved Water Use Improves the Yield Performance of Soybean (Glycine max (L.) Merr.) under Drought. Agric. Water Manag. 2017, 179, 236–245. [Google Scholar] [CrossRef]
- Leiser, W.L.; Rattunde, H.F.W.; Piepho, H.-P.; Weltzien, E.; Diallo, A.; Toure, A.; Haussmann, B.I.G. Phosphorous Efficiency and Tolerance Traits for Selection of Sorghum for Performance in Phosphorous-Limited Environments. Crop Sci. 2015, 55, 1152–1162. [Google Scholar] [CrossRef]
- Li, H.; Hu, B.; Chu, C. Nitrogen Use Efficiency in Crops: Lessons from Arabidopsis and Rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GY | Shoot DW | HI | WU | WUE | TP | TN | PUE | NUE | Root DW | Nodule DW | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes (G) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| P | *** | *** | *** | *** | *** | *** | *** | *** | *** | ||
| Water (W) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| G × P | * | ** | * | ** | *** | ||||||
| G × W | *** | *** | *** | *** | *** | *** | *** | ||||
| P × W | *** | *** | *** | *** | ** | *** | *** | ** | * | *** | |
| G × P × W | ** | ** | * | *** | * | ** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Y.; He, J.; Zhu, Y.; Siddique, K.H.M. Nodule Formation and Nitrogen Use Efficiency Are Important for Soybean to Adapt to Water and P Deficit Conditions. Agriculture 2022, 12, 1326. https://doi.org/10.3390/agriculture12091326
Jin Y, He J, Zhu Y, Siddique KHM. Nodule Formation and Nitrogen Use Efficiency Are Important for Soybean to Adapt to Water and P Deficit Conditions. Agriculture. 2022; 12(9):1326. https://doi.org/10.3390/agriculture12091326
Chicago/Turabian StyleJin, Yi, Jin He, Yonghe Zhu, and Kadambot H. M. Siddique. 2022. "Nodule Formation and Nitrogen Use Efficiency Are Important for Soybean to Adapt to Water and P Deficit Conditions" Agriculture 12, no. 9: 1326. https://doi.org/10.3390/agriculture12091326