1. Introduction
Durum wheat (
Triticum turgidum subsp.
durum (Desf.) Husnot) is an essential crop for food and feeding products. Its cultivation area reached 17 million ha worldwide in 2019, and global production was around 38.1 million tonnes [
1]. In the Mediterranean basin, the cultivated area is tending to increase [
2,
3], due to the adaptation of the crop to stress prone conditions in terms of both abiotic [
3,
4,
5] and biotic adversities [
6,
7]. Moreover, many research projects aim to increase durum wheat’s stability in contrasting environments by utilizing wild crop relatives [
8,
9,
10,
11].
Developing an improved elite cultivar through breeding programs is a time-consuming and costly process, but is necessary for wheat’s adaptation to continuously changing environmental conditions. Thus, the longevity of the elite varieties is of the utmost importance. Relevant research in many crops has encouraged intense selection at nil competition (e.g., ultra-low density) as a useful tool for either upgrading or avoiding a gradual degradation of the genetic background to maintain uniformity and ensure optimum quality of breeder seed for longer time periods [
12]. Moreover, the long-term selection studies on maize for oil and protein undertook for almost a century at the University of Illinois [
13] showed that continuous intracultivar breeding is a workable option for handling intracultivar variation. A different approach from the classical breeding program is the valuable exploitation of the intra-cultivar phenotypic variation through selection at nil competition, as shown in numerous studies on plants such as corn (
Zea mays L.) [
14], soybean [
Glycine max (L.) Merr.] [
15], wheat (
Triticum aestivum L.) [
16], cotton (
Gossypium hirsutum L.) [
17], snap bean (
Phaseolus vulgaris L.) [
18] and rice (
Oryza sativa L.) [
19]. This technique, i.e., the selection at nil competition, is a short-time tool for selection within landraces to breed for high yielding and stable varieties [
20,
21,
22,
23,
24]. Many projects using molecular techniques have revealed genetic heterogeneity within cultivars [
19,
25,
26]. Advances in DNA-based techniques contributed to discovering genetic variation at the genomic level and verified significant heterogeneity [
27]. It is well documented that the genome is more variable than previously assumed [
13,
28] and has endogenous mechanisms to be flexible and dynamic, characterized by high plasticity. In her Nobel Prize-winning lecture, McClintock [
29] argued that the genomes of many organisms are susceptible to unusual and unexpected events and can modify themselves in response to environmental forces. The genome undergoes constant remodelling and restructuring by utilizing an array of different mechanisms, such as intragenic recombination, unequal crossing over, DNA methylation, excision or insertion of transposable elements, gene duplication and genetic restoration events [
30,
31,
32,
33,
34]. All the above-mentioned factors enlarge the genetic variation within the improved cultivars [
35,
36] and provide evidence that inbred lines contain genetic variation that can be further utilized in a breeding program, and the impact of intra-cultivar genetic heterogeneity may be significant [
25,
37].
The study of this de novo variation is feasible by applying the honeycomb design [
38], which permits optimal conditions to maximize the phenotypic expression and thus [
39] allows continuous selection within the elite cultivars. Elite cultivars are not homogeneous but rather heterogeneous, within which selections can be made to maintain or improve uniformity and further improve desirable agronomic traits [
40]. The intra-cultivar selection relies on distinguishing outstanding genotypes within a narrow gene pool. Many reports confirm that intra-cultivar selection under ULD (i.e., nil competition) maximizes the phenotypic expression and differentiation of the genοtype [
40,
41,
42]. Tokatlidis [
12] reports that intense selection at ULD is a useful technique to upgrade or avoid gradual degradation of the genetic background. The importance of non-stop intra-cultivar selection was highlighted by Fasoula [
41] and Fasoula and Fasoula [
43]. Honeycomb intra-cultivar selection in wheat [
16,
39,
41], maize [
14], cotton [
17,
40,
44] and soybean [
15,
45] succeeded in upgrading cultivars for yield, stability and important agronomic traits related to product quality and tolerance to stresses. While there have been case studies of intra-cultivar breeding with bread wheat at ULD [
16,
39,
41], there is a lack of information about continuous selection in durum wheat.
Although there is considerable variability within a variety that can be used for intra-variety selection [
39,
40,
46], the small number of seeds of individual plants does not allow the measurement of quality characteristics such as nutrient concentrations via the destructive classical analytical methods. The seeds are of course necessary for the breeding process though. A solution to this problem could be the use of the near-infrared spectroscopy (NIR) technology for non-destructive analysis of small seed samples (protein, starch, gluten, etc.) [
47]. This would allow the analysis of the seed quality of various genotypes without destroying the seeds and would enable sowing in the next growing season to continue the selection process. This method has become the most widely used alternative to conventional analytical methods due to its rapidity, simplicity, accuracy, cost-effectiveness and potential for routine analysis and quantification of nutrients in food products and crops [
48]. Despite the small seed quantity required for measuring seed quality traits using NIR, the seed quantity limitation can still be a challenge when assessing individual plants, especially when plants are evaluated under dense stands. Thus, there is a lack of studies on the evaluation of durum wheat seed quality traits on an individual plant basis. The ULD approach tackles the problem since durum wheat plants grown far apart can produce enough seeds for quality evaluation and selection and line advancement in the next generation.
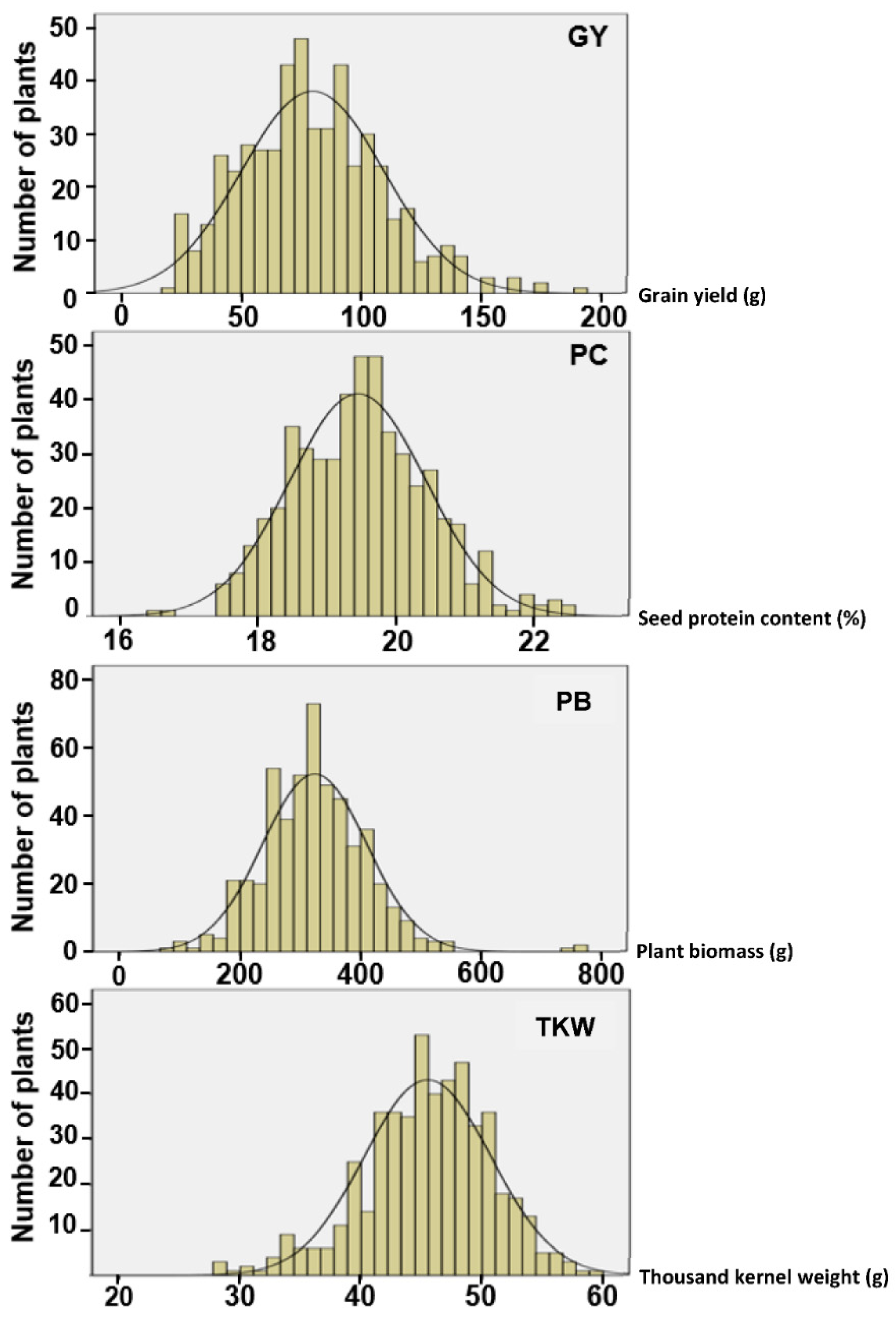
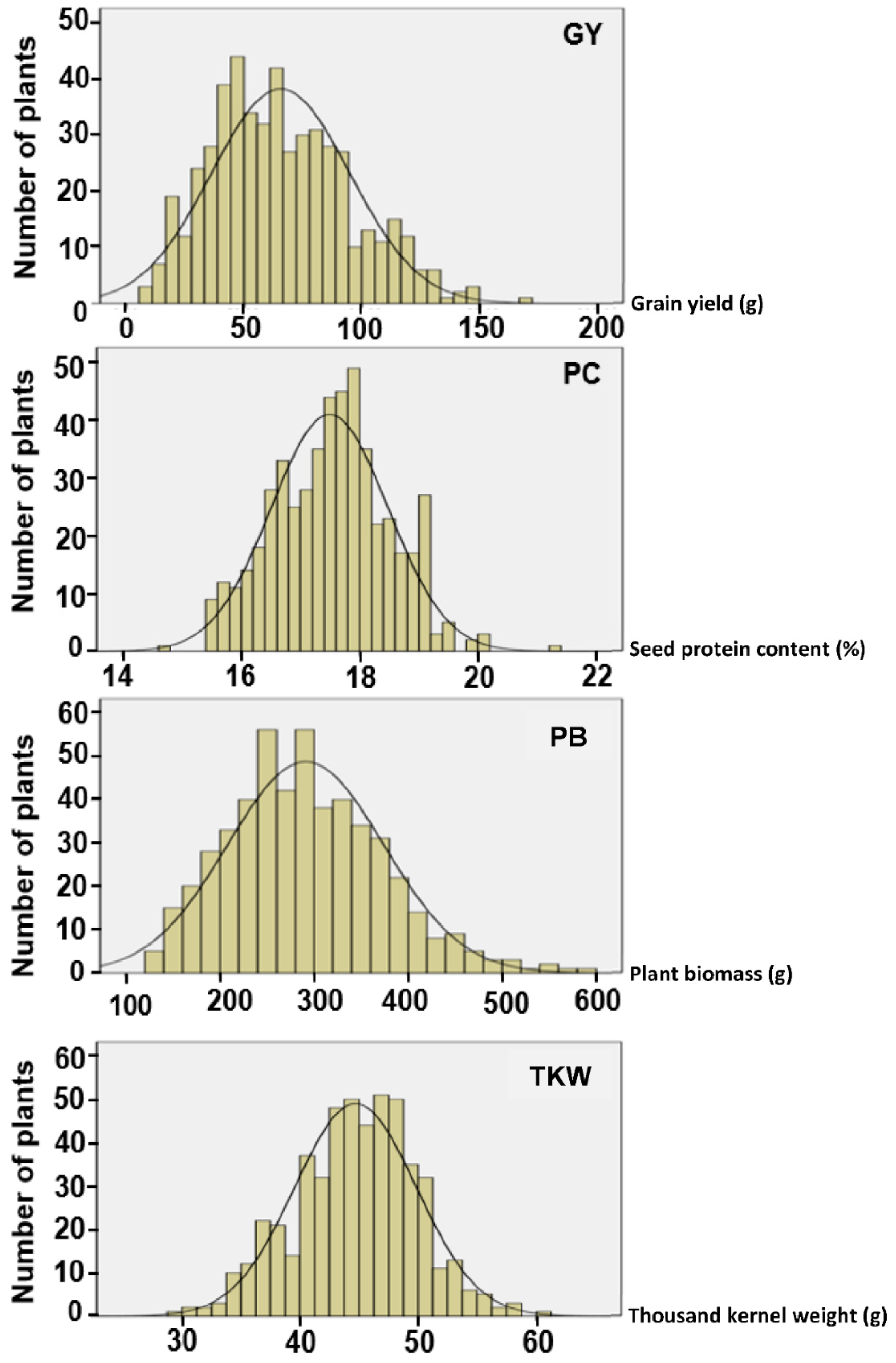
Thus, the main objectives of this study were to assess the phenotypic variations in grain yield (GY) and protein content (PC) within two elite commercial durum wheat cultivars and evaluate the potential of selection for GY and PC, to investigate whether the variations are exploitable and could result in GY and PC upgrades for those cultivars.
4. Discussion
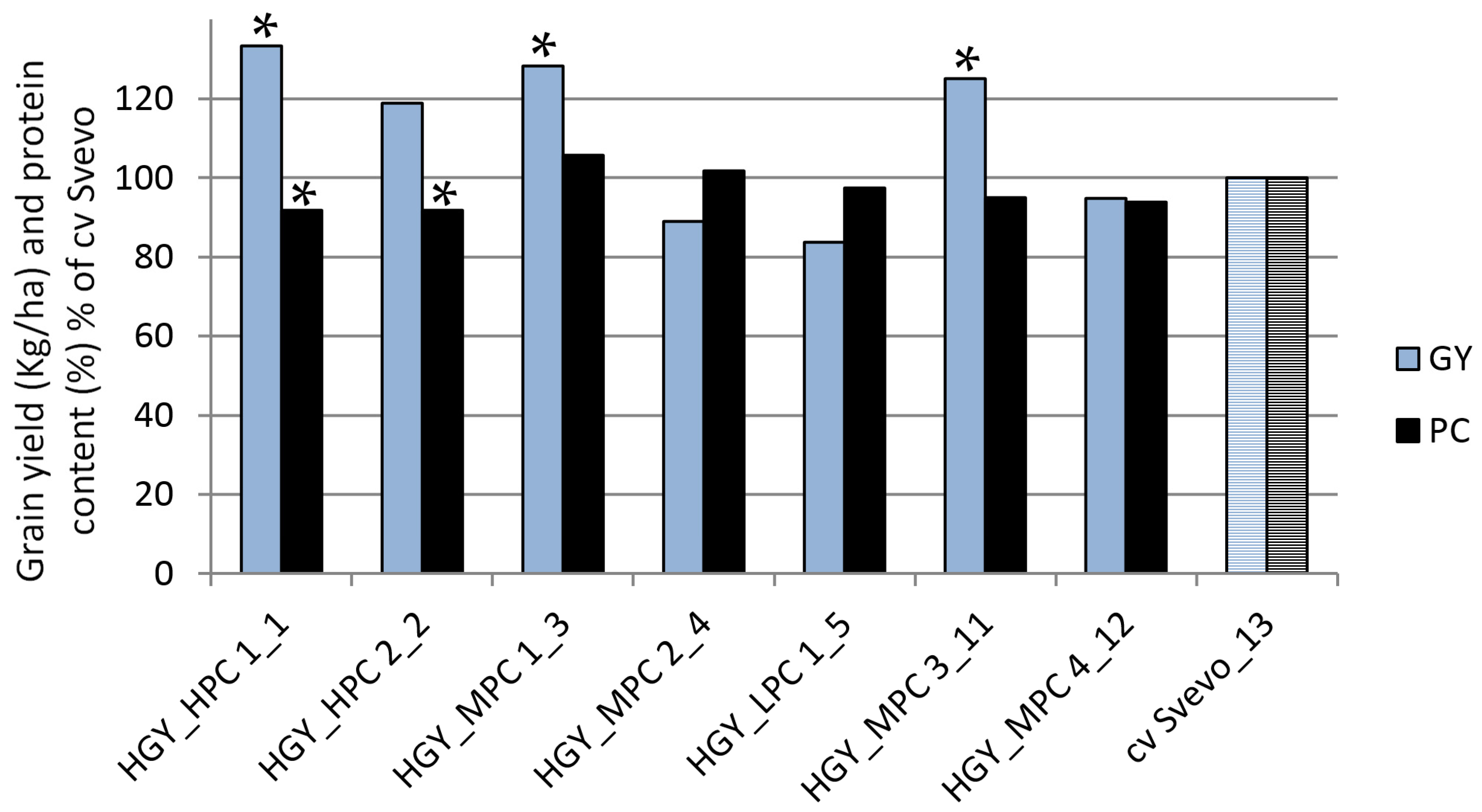
This study developed two distinct sets of durum wheat lines by applying divergent single-plant selection under ultra-low plant density to two elite durum wheat commercial cultivars. The results revealed that selection under ultra-low density is an effective tool to select simultaneously for GY and PC. Both traits are of major importance in durum wheat breeding [
52]. However, PC in durum wheat is negatively associated with GY [
53,
54], so breeding for higher yield can decrease PC [
55]. Nevertheless, as Simmonds [
56] highlighted, despite the consensus on a strong negative correlation between GY and PC in cereals, a positive expected relationship also holds by making some compromises between the attainable higher yield and the protein concentration. Indeed, in our case, we identified that three lines derived from cv Svevo significantly surpassed the source material in terms of GY, with one maintaining a high PC concurrently. In addition, another two lines derived from cv Maestrale outperformed the source material in GY, even though the difference did not reach significant levels. Still, these lines had significantly higher PC than the original genotype at the same time, implying that intra-cultivar selection under ultra-low density was effective at upgrading at least one target trait.
Numerous studies have been conducted on the simultaneous selection of PC and GY in wheat, employing index selection [
57,
58,
59,
60], tailored with genome-wide association mapping and prediction models [
52,
61], providing evidence of efficient concurrent selection, even though results sometimes can be genotype or environment dependent. However, all these studies have been processed with very diverse sets of germplasm. In our study, though, we evaluated and distinguished for yield and PC among lines of very narrow genetic diversity, since these lines were derived through intra-cultivar selection. While breeders commonly believe that intra-cultivar selection within a fairly homogeneous germplasm cannot lead to high amounts of genetic gain due to limitations of the genetic variance component, new debates have come to light in this regard. Selection within the bread wheat cultivar Nestos for two generations at 1.2 plants∙m
2 confirmed an improvement in GY of up to 22% per unit area under typical farmers’ stands [
39]. In barley, two years of selection under ultra-low density within the cultivar Athenaida resulted in lines that showed average yield increases of up to 2.5 times over the original genotype when tested under typical farm densities in two locations [
41]. Similar results on the efficiency of intra-cultivar selection have also been obtained in other predominately self-pollinated crops, other than cereals. Tokatlidis et al. [
17] selected three cotton commercial cultivars for two years at the density of 1.15 plants∙m
−2, and succeeded in identifying lines that, when tested under typical crop density, there were up to 12% higher cotton seed yield on average across the sites and years compared to the original cultivars. Furthermore, in terms of quality traits, selecting for protein and oil content within three soybean cultivars using a density of 1.4 plants∙m
−2, Fasoula and Boerma [
15] reported considerable variation for these two traits within each of the three cultivars. Our results confirm the above studies not only for the higher GY achieved when we selected within two elite durum wheat cultivars, but at the same time for the simultaneous upgrading of the PC, which in the case of the one cultivar (cv Maestrale) reached significant levels in comparison to the original genotype.
Evidence from molecular markers studies elucidate further the concept of exploitable intra-cultivar variation within homogeneous germplasm and further enhance our results. Using RFLP and microsatellite markers, Olufowote et al. [
19] detected significant variation within rice landraces and cultivars that were assumed to be pure lines. In barley, Ben Ghanem et al. [
50] detected variation within landraces and elite commercial varieties for the
RYd2 and
RYd3 alleles that allow resistance to barley yellow dwarf virus (BYDV). The intrinsic amount of genetic variation detected within homogeneous gene pools can be attributed either to latent genetic diversity or to mechanisms that can generate de novo variation. Hence, residual heterozygosity, due to differential segregation of polymorphic chromosomal regions in the breeding process’s successive generations, has been concluded to be one source of structural intra-cultivar genetic variation in soybean [
37]. On the other hand, additional heterogeneity might stem from de novo generated variation, as a result of spontaneous mutations [
62,
63] or via genetic and epigenetic mechanisms, such as intragenic recombination, unequal crossing over, gene duplications or deletions, DNA methylation, excision or insertion of transposable elements, chromatin alterations and others [
32,
64,
65,
66].
Detecting exploitable intra-cultivar variation, though, is a challenging procedure. Potential response to selection relies on the ability to distinguish outstanding genotypes within a narrow gene pool [
12]. Thus, all the above studies that recorded significant progress by applying intra-cultivar variation have been conducted via selection under ultra-low density, in conditions that practically eliminate plant to plant interference and simulate a nil-competition regime. Notwithstanding, the argument that usually pertains among breeders is that selection should be applied under a dense stand regime similar to farming conditions [
46,
67]. At the same time, though, the consensus among breeders is that the optimum environment for selection is the one that maximizes genetic variation and hence responds to selection [
68,
69]. However, the ultra-low planting density regime will ensure the conditions of nil-competition among plants and allow for maximum phenotypic expression of their genetic differences, ultimately facilitating the selection of desirable genotypes [
12,
70]. Moreover, Tokatlidis [
46] raised another issue as a counterargument to the claim of a lack of connection between plants spaced apart for selection purposes and those densely grown by questioning whether segregating generations can simulate farming conditions. He highlighted that solid seeded conditions and nil-competition may not correlate when heterogeneous populations are evaluated due to strong competitor genotypes and the inverse association between yielding and competitive ability [
46]. Several studies on different crops, such as rye [
71], vetch [
72] and wheat [
73], have well demonstrated this negative relationship between yielding and competitive ability. Nevertheless, as Tokatlidis [
46] mentioned, solid seeded and nil-competition do correlate well in the case of genetically homogeneous lines, something that also agrees with the findings of our study, since we found high positive correlation coefficients for GY between ultra-low density and typical crop density for both cultivars.
Overall, the development of durum wheat varieties that combine high GY and high PC is one of the significant challenges of modern plant breeding, since these are the two most critical factors determining the crop’s economic value. Thus, exploiting intra-cultivar variation through simultaneous selection for GY and PC under ultra-low density conditions seems an efficient tool to surpass the negative association between these two traits and to develop elite material characterized by high GY, stability of performance and upgraded quality traits, meeting the future demands of the market.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}