
Multi-Omic Approaches to Investigate Molecular Mechanisms in Peach Post-Harvest Ripening


Abstract
:1. Introduction
2. The Peach Ripening Process
2.1. Compounds Involved
2.2. Volatilome and Profiling Techniques
3. Omics Advances in Rosaceae
3.1. Genomic and Transcriptomic Peach Profiles
3.2. Metabolomic Peach Profiles
4. Peach Gene Expression and Correlation with VOCs
5. Multi-Omics Approaches and the Peach Post-Harvest Ripening Process
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aranzana, M.J.; Decroocq, V.; Dirlewanger, E.; Eduardo, I.; Gao, Z.S.; Gasic, K.; Arús, P. Prunus genetics and applications after de novo genome sequencing: Achievements and prospects. Hortic. Res. 2019, 6, 1–25. [Google Scholar]
- Sánchez, G.; Besada, C.; Badenes, M.L.; Monforte, A.J.; Granell, A. A non-targeted approach unravels the volatile network in peach fruit. PLoS ONE 2012, 7, e38992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gapper, N.; McQuinn, R.P.; Giovannoni, J.J. Molecular and genetic regulation of fruit ripening. Plant Mol. Biol. 2013, 82, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Aubert, C.; Gunata, Z.; Ambid, C.; Baumes, R. Changes in Physicochemical Characteristics and Volatile Constituents of Yellow- and White-Fleshed Nectarines during Maturation and Artificial Ripening. J. Agric. Food Chem. 2003, 51, 3083–3091. [Google Scholar] [CrossRef]
- Zhang, B.; Shen, J.Y.; Wei, W.W.; Xi, W.P.; Xu, C.J.; Ferguson, I.; Chen, K. Expression of Genes Associated with Aroma Formation Derived from the Fatty Acid Pathway during Peach Fruit Ripening. J. Agric. Food Chem. 2010, 58, 6157–6165. [Google Scholar] [CrossRef]
- Robertson, J.A.; Meredith, F.I.; Horvat, R.J.; Senter, S.D. Effect of cold storage and maturity on the physical and chemical characteristics and volatile constituents of peaches (cv. Cresthaven). J. Agric. Food Chem. 1990, 38, 620–624. [Google Scholar] [CrossRef]
- Muto, A.; Müller, C.T.; Bruno, L.; McGregor, L.; Ferrante, A.; Chiappetta, A.; Bitonti, M.B.; Rogers, H.J.; Spadafora, N.D. Fruit volatilome profiling through GC × GC-ToF-MS and gene expression analyses reveal differences amongst peach cultivars in their response to cold storage. Sci. Rep. 2020, 10, 18333. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, C.; Li, S.; Yang, L.; Wang, Y.; Zhao, J.; Jiang, Q. Volatile characteristics of 50 peaches and nectarines evaluated by HP-SPME with GC-MS. Food Chem. 2009, 116, 356–364. [Google Scholar] [CrossRef]
- Aubert, C.; Milhet, C. Distribution of the volatile compounds in the different parts of a white-fleshed peach (Prunus persica L. Batsch). Food Chem. 2007, 102, 375–384. [Google Scholar] [CrossRef]
- Song, C.; Hong, X.; Zhao, S.; Liu, J.; Schulenburg, K.; Huang, F.C.; Franz-Oberdorf, K.; Wilfried Schwab, W. Glucosylation of 4-hydroxy-2,5-dimethyl-3(2H)-furanone, the key strawberry flavor compound in strawberry fruit. Plant Physiol. 2016, 171, 139–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttery, R.G.; Teranishi, R.; Ling, L.C.; Turnbaugh, J.G. Quantitative and sensory studies on tomato paste volatiles. J. Agric Food Chem. 1990, 38, 336–340. [Google Scholar] [CrossRef]
- Zhu, J.; Xiao, Z. Characterization of the key aroma compounds in peach by gas chromatography–olfactometry, quantitative measurements and sensory analysis. Eur. Food Res. Technol. 2019, 245, 129–141. [Google Scholar] [CrossRef]
- Xi, W.; Zheng, Q.; Lu, J.; Quan, J. Comparative analysis of three types of peaches: Identification of the key individual characteristic flavor compounds by integrating consumers’ acceptability with flavor quality. Hortic. Plant J. 2017, 3, 1–12. [Google Scholar] [CrossRef]
- Biale, J.B.; Young, R.E. Respiration and ripening in fruits-retrospective and prospect. In Recent Advances in the Biochemistry of Fruits and Vegetables; Friend, J., Rhodes, M.J.C., Eds.; Academic Press: New York, NY, USA, 1981; pp. 1–39. [Google Scholar]
- Seymour, G.B.; Østergaard, L.; Chapman, N.H.; Knapp, S.; Martin, C. Fruit development and ripening. Annu. Rev. Plant Biol. 2013, 64, 219–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Defilippi, B.G.; Manriquez, D.; Luengwilai, K.; Gonzalez-Aguero, M. Aroma Volatiles: Biosynthesis and Mechanisms of Modulation during Fruit Ripening. Adv. Bot. Res. 2009, 50, 1–37. [Google Scholar]
- Liu, M.C.; Pirrello, J.; Chervin, C.; Roustan, J.P.; Bouzayen, M. Ethylene Control of Fruit Ripening: Revisiting the Complex Network of Transcriptional Regulation. Plant Physiol. 2015, 169, 2380–2390. [Google Scholar] [CrossRef] [Green Version]
- Yun, Z.; Jin, S.; Ding, Y.; Wang, Z.; Gao, H.; Pan, Z.; Xu, J.; Cheng, Y.; Deng, X. Comparative transcriptomics and proteomics analysis of citrus fruit, to improve understanding of the effect of low temperature on maintaining fruit quality during lengthy post-harvest storage. J. Exp. Bot. 2012, 63, 2873–2893. [Google Scholar] [CrossRef]
- Tanou, G.; Minas, I.S.; Scossa, F.; Belghazi, M.; Xanthopoulou, A.; Ganopoulos, I.; Madesis, P.; Fernie, A.; Molassiotis, A. Exploring priming responses involved in peach fruit acclimation to cold stress. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Lurie, L.; Crisosto, C.H. Chilling injury in peach and nectarine. Postharvest Biol. Technol. 2005, 37, 195–208. [Google Scholar] [CrossRef]
- Brizzolara, S.; Manganaris, G.A.; Fotopoulos, V.; Watkins, C.B.; Tonutti, P. Primary metabolism in fresh fruits during storage. Front. Plant Sci. 2020, 11, 80. [Google Scholar] [CrossRef] [Green Version]
- Manganaris, G.A.; Crisosto, C.H. Stone fruits: Peaches, nectarines, plums, apricots. In Controlled and Modified Atmospheres for Fresh and Fresh-Cut Produce; Elsevier: Amsterdam, The Netherlands, 2020; pp. 311–322. [Google Scholar]
- Pons, C.; Martí, C.; Forment, J.; Crisosto, C.H.; Dandekar, A.M.; Granell, A. A bulk segregant gene expression analysis of a peach population reveals components of the underlying mechanism of the fruit cold response. PLoS ONE 2014, 9, e90706. [Google Scholar] [CrossRef] [PubMed]
- Salvo, M.; Rey, F.; Arruabarrena, A.; Gambetta, G.; Rodrigo, M.J.; Zacarías, L.; Lado, J. Transcriptional Analysis of C-Repeat Binding Factors in Fruit of Citrus Species with Differential Sensitivity to Chilling Injury during Postharvest Storage. Int. J. Mol. Sci. 2021, 22, 804. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J. Chilling injury and phytochemical composition of peach fruits as affected by high carbon dioxide treatment before cold storage. Hortic. Environ. Biotechnol. 2014, 55, 190–195. [Google Scholar] [CrossRef]
- Jin, P.; Shang, H.; Chen, J.; Zhu, H.; Zhao, Y.; Zheng, Y. Effect of 1-Methylcyclopropene on Chilling Injury and Quality of Peach Fruit during Cold Storage. J. Food Sci. 2011, 76, 485–491. [Google Scholar] [CrossRef]
- Abdipour, M.; Hosseinifarahi, M.; Naseri, N. Combination method of UV-B and UV-C prevents post-harvest decay and improves organoleptic quality of peach fruit. Sci. Hortic. 2019, 256, 108564. [Google Scholar] [CrossRef]
- Kan, J.; Hui, Y.; Lin, X.; Liu, Y.; Jin, C. Postharvest ultraviolet-C treatment of peach fruit: Changes in transcriptome profile focusing on genes involved in softening and senescence. J. Food Process Preserv. 2021, 10, e15813. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Vicente, A.R.; Martínez-García, P.J.; Crisosto, C.H. Peach and nectarine. In Postharvest Physiological Disorders in Fruits and Vegetables; CRC Press: Boca Raton, FL, USA, 2019; pp. 293–304. [Google Scholar]
- Gapper, N.; Giovannoni, J.; Watkins, C. Understanding development and ripening of fruit crops in an ‘omics’ era. Hortic. Res. 2014, 1, 14034. [Google Scholar] [CrossRef] [Green Version]
- Amer, B.; Baidoo, E.E.K. Omics-Driven Biotechnology for Industrial Applications. Front. Bioeng. Biotechnol. 2021, 9, 30. [Google Scholar] [CrossRef]
- Bonghi, C.; Trainotti, L.; Botton, A.; Tadiello, A.; Rasori, A.; Ziliotto, F.; Ramina, A. A microarray approach to identify genes involved in seed-pericarp cross-talk and development in peach. BMC Plant Biol. 2011, 11, 107. [Google Scholar] [CrossRef] [Green Version]
- García-Gómez, B.E.; Salazar, J.A.; Nicolás-Almansa, M.; Razi, M.; Rubio, M.; Ruiz, D.; Martínez-Gómez, P. Molecular Bases of Fruit Quality in Prunus Species: An Integrated Genomic, Transcriptomic, and Metabolic Review with a Breeding Perspective. Int. J. Mol. Sci. 2020, 22, 333. [Google Scholar] [CrossRef]
- Trainotti, L.; Tadiello, A.; Casadoro, G. The involvement of auxin in the ripening of climacteric fruits comes of age: The hormone plays a role of its own and has an intense interplay with ethylene in ripening peaches. J. Exp. Bot. 2007, 58, 3299–3308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonghi, C.; Begheldo, M.; Ziliotto, F.; Rasori, A.; Trainotti, L.; Tadiello, A.; Tosetti, R.; Tonutti, P. Transcriptome analyses and postharvest physiology of peaches and nectarines. Acta Hortic. 2010, 877, 69–73. [Google Scholar] [CrossRef]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Romero, D.; Valero, D.; Serrano, M.; Burlo, F.; Carbonell, A.; Burgos, L.; Riquelme, F. Exogenous Polyamines and Gibberellic Acid Effects on Peach (Prunus persica L.) Storability Improvement. J. Food Sci. 2000, 65, 288–294. [Google Scholar] [CrossRef]
- Bakshi, A.; Shemansky, J.M.; Chang, C.R.; Binder, B.M. History of Research on the Plant Hormone Ethylene. J. Plant Growth Regul. 2015, 34, 809–827. [Google Scholar] [CrossRef]
- Palou, L.; Crisosto, C.H. Postharvest Treatments to Reduce the Harmful Effects of Ethylene on Apricots. Acta Hortic. 2003, 599, 31–38. [Google Scholar] [CrossRef]
- Brady, C.J. Stone fruit. In Biochemistry of Fruit Ripening; Springer: Dordrecht, The Netherlands, 1993; pp. 379–404. [Google Scholar]
- Soto, A.; Ruiz, K.B.; Ravaglia, D.; Costa, G.; Torrigiani, P. ABA may promote or delay peach fruit ripening through modulation of ripening- and hormone-related gene expression depending on the developmental stage. Plant Physiol. Biochem. 2013, 64, 11–24. [Google Scholar] [CrossRef]
- Han, M.; Zhang, T.; Zhao, C.; Zhi, J. Regulation of the expression of lipoxygenase genes in Prunus persica fruit ripening. Acta Physiol. Plant 2011, 33, 1345–1352. [Google Scholar] [CrossRef]
- Soto, A.; Ruiz, K.B.; Ziosi, V.; Costa, G.; Torrigiani, P. Ethylene and auxin biosynthesis and signaling are impaired by methyl jasmonate leading to a transient slowing down of ripening in peach fruit. J. Plant Physiol. 2012, 169, 1858–1865. [Google Scholar] [CrossRef]
- Ziosi, V.; Bonghi, C.; Bregoli, A.M.; Trainotti, L.; Stefania Biondi, S.; Sutthiwal, S.; Kondo, S.; Costa, G.; Torrigiani, P. Jasmonate-induced transcriptional changes suggest a negative interference with the ripening syndrome in peach fruit. J. Exp. Bot. 2008, 59, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Peña-Cortés, H.; Barrios, P.; Dorta, F.; Polanco, V.; Sanchez, C.; Sanchez, E.; Ramırez, I. Involvement of jasmonic acid and derivatives in plant responses to pathogens and insects and in fruit ripening. J. Plant Growth Regul. 2005, 23, 246–260. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Honda, C.; Kondo, S. Effect of lowtemperature stress on abscisic acid, jasmonates polyamines in apples. Plant Growth Regul. 2007, 52, 199–206. [Google Scholar] [CrossRef]
- Yao, H.; Tian, S. Effects of pre- and post-harvest application of salicylic acid or methyl jasmonate on inducing disease resistance of sweet cherry in fruit storage. Postharvest Biol. Technol. 2005, 35, 253–262. [Google Scholar] [CrossRef]
- Ravaglia, D.; Espley, R.V.; Henry-Kirk, R.A.; Andreotti, C.; Ziosi, V.; Hellens, R.P.; Costa, G.; Allan, A.C. Transcriptional regulation of flavonoid biosynthesis in nectarine (Prunus persica) by a set of R2R3 MYB transcription factors. BMC Plant Biol. 2013, 13, 68. [Google Scholar] [CrossRef] [Green Version]
- Winterhalter, P.; Rouseff, R.L. Carotenoid-Derived Aroma Compounds. ACS Symposium Series 802; American Chemical Society: Washington, DC, USA, 2002. [Google Scholar]
- Brandi, F.; Bar, E.; Mourgues, F.; Horváth, G.; Turcsi, E.; Giuliano, G.; Liverani, A.; Tartarini, S.; Lewinsohn, E.; Rosati, C. Study of Redhaven peach and its white-fleshed mutant suggests a key role of CCD4 carotenoid dioxygenase in carotenoid and norisoprenoid volatile metabolism. BMC Plant Biol. 2011, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.; Yousef, G.; Guzman, I.; Chebrolu, K.; Werner, D.J.; Parker, M.; Gasic, K.; Perkins-Veazie, P. Variation of Carotenoids and Polyphenolics in Peach (Prunus persica L.) and Implications on Breeding for Modified Phytochemical Profiles. J. Am. Soc. Hortic. Sci. 2014, 139, 676. [Google Scholar] [CrossRef] [Green Version]
- Aubert, C.; Bony, P.; Chalot, G.; Landry, P.; Lurol, S. Effects of storage temperature, storage duration, and subsequent ripening on the physicochemical characteristics, volatile compounds, and phytochemicals of western red nectarine (Prunus persica L. Batsch). J. Agric. Food Chem. 2014, 62, 4707–4724. [Google Scholar] [CrossRef]
- Caprioli, I.; Lafuente, M.T.; Rodrigo, M.; Mencarelli, F. Influence of Postharvest Treatments on Quality, Carotenoids, and Abscisic Acid Content of Stored “Spring Belle” Peach (Prunus persica) Fruit. J. Agric. Food Chem. 2009, 57, 7056–7063. [Google Scholar] [CrossRef]
- Aubert, C.; Chalot, G.; Sébastien, L.; Ronjon, A.; Cottet, V. Relationship between fruit density and quality parameters, levels of sugars, organic acids, bioactive compounds and volatiles of two nectarine cultivars, at harvest and after ripening. Food Chem. 2019, 297, 124954. [Google Scholar] [CrossRef]
- Dalla Valle, Z.; Mignani, I.; Spinardi, A.; Galvano, F.; Ciappellano, S. The antioxidant profile of three different peaches cultivars (Prunus persica) and their short-term effect on antioxidant status in human. Eur. Food Res. Technol. 2007, 225, 167–172. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Drogoudi, P.; Goulas, V.; Tanou, G.; Georgiadou, E.C.; Pantelidis, G.; Paschalidis, K.; Fotopoulos, V.; Manganaris, A. Deciphering the interplay among genotype, maturity stage and low-temperature storage on phytochemical composition and transcript levels of enzymatic antioxidants in Prunus persica fruit. Plant Physiol. Biochem. 2017, 119, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, W.; Yin, X.; Su, M.; Sun, C.; Li, X.; Chen, K. Phenolic composition and antioxidant properties of different peach [Prunus persica (L.) Batsch] cultivars in China. Int. J. Mol. Sci. 2015, 16, 5762–5778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drogoudi, P.; Pantelidis, G.E.; Goulas, V.; Manganaris, G.A.; Ziogas, V.; Manganaris, A. The appraisal of qualitative parameters and antioxidant contents during postharvest peach fruit ripening underlines the genotype significance. Postharvest Biol. Technol. 2016, 115, 142–150. [Google Scholar] [CrossRef]
- Sevenants, M.R.; Jennings, W.G. Volatile components of peach. II. J. Food Sci. 1996, 31, 81–86. [Google Scholar] [CrossRef]
- Do, J.Y.; Salunkhe, D.K.; Olson, L.E. Isolation, identification and comparison of the volatiles of peach fruit as related to harvest maturity and artificial ripening. J. Food Sci. 1969, 34, 618–621. [Google Scholar] [CrossRef]
- Spencer, M.D.; Pangborn, R.M.; Jennings, W.G. Gas chromatographic and sensory analysis of volatiles from cling peaches. J. Agric. Food Chem. 1978, 26, 725–732. [Google Scholar] [CrossRef]
- Horvat, R.J.; Chapman, G.W.; Robertson, J.A.; Meredith, F.I.; Scorza, R.; Callahan, A.M.; Morgens, P. Comparison of the volatile compounds from several commercial peach cultivars. J. Agric. Food Chem. 1990, 38, 234–237. [Google Scholar] [CrossRef]
- Jia, H.; Okamoto, G. Distribution of volatile compounds in peach fruit. J. Jpn. Soc. Hortic. Sci. 2001, 70, 223–225. [Google Scholar] [CrossRef]
- Eduardo, I.; Chietera, G.; Bassi, D.; Rossini, L.; Vecchietti, A. Identification of key odor volatile compounds in the essential oil of nine peach accessions. J. Sci. Food Agric. 2010, 90, 1146–1154. [Google Scholar] [CrossRef]
- Li, X.W.; Jiang, J.; Zhang, L.P.; Yu, Y.; Ye, Z.W.; Wang, X.; Zhang, H.; Arús, P.; Jia, H.; Gao, Z.S. Identification of volatile and softening-related genes using digital gene expression profiles in melting peach. Tree Genet. Genomes 2015, 11, 71. [Google Scholar] [CrossRef]
- Zhang, L.; Li, H.; Gao, L.; Qi, Y.; Fu, W.; Li, X.; Zhou, X.; Gao, Q.; Gao, Z.; Jia, H. Acyl-CoA oxidase 1 is involved in γ-decalactone release from peach (Prunus persica) fruit. Plant Cell Rep. 2017, 36, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Derail, C.; Hofmann, T.; Schieberle, P. Differences in Key Odorants of Handmade Juice of Yellow-Flesh Peaches (Prunus persica L.) Induced by the Workup Procedure. J. Agric. Food Chem. 1999, 47, 4742–4745. [Google Scholar] [CrossRef] [PubMed]
- Bellincontro, A.; Morganti, F.; DeSantis, D.; Botondi, R.; Mencarelli, F. Inhibition of ethylene via different ways affects LOX and ADH activities, and related volatiles compounds in peach (cv. ‘Royal Gem’). Acta Hortic. 2005, 682, 445–452. [Google Scholar] [CrossRef]
- Aragüez, I.; Valpuesta, V. Metabolic engineering of aroma components in fruits. Biotechnol. J. 2013, 8, 1144–1158. [Google Scholar] [CrossRef]
- Sánchez, G.; Venegas-Calerón, M.; Salas, J.J.; Monforte, A.; Badenes, M.L.; Granell, A. An integrative “omics” approach identifies new candidate genes to impact aroma volatiles in peach fruit. BMC Genom. 2013, 14, 343. [Google Scholar] [CrossRef] [Green Version]
- Visai, C.; Vanoli, M. Volatile compound production during growth and ripening of peaches and nectarines. Sci. Hortic. 1997, 70, 15–24. [Google Scholar] [CrossRef]
- Osorio, S.; Scossa, F.; Fernie, A.R. Molecular regulation of fruit ripening. Front. Plant Sci. 2013, 4, 198. [Google Scholar] [CrossRef] [Green Version]
- Farina, V.; Lo Bianco, R.; Di Marco, L. Fruit quality and flavour compounds before and after commercial harvest of the late-ripening “Fairtime” peach cultivar. Int. J. Fruit Sci. 2007, 7, 25–36. [Google Scholar] [CrossRef]
- Raffo, A.; Nardo, N.; Tabilio, M.R.; Paoletti, F. Effects of cold storage on aroma compounds of white- and yellow-fleshed peaches. Eur. Food Res. Technol. 2008, 226, 1503–1512. [Google Scholar] [CrossRef]
- Zhang, B.; Xi, W.P.; Wei, W.W.; Shen, J.Y.; Ferguson, I.; Chen, K.S. Changes in aroma-related volatiles and gene expression during low temperature storage and subsequent shelf-life of peach fruit. Postharvest. Biol. Technol. 2011, 60, 7–16. [Google Scholar] [CrossRef]
- Brizzolara, S.; Hertog, M.; Tosetti, R.; Nicolai, B.; Tonutti, P. Metabolic responses to low temperature of three peach fruit cultivars differently sensitive to cold storage. Front. Plant Sci. 2018, 9, 706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, D.D.; Powers, R. Beyond the paradigm: Combining mass spectrometry and nuclear magnetic resonance for metabolomics. Prog. Nucl. Magn. Reson. Spectrosc. 2017, 100, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroymann, J. Natural diversity and adaptation in plant secondary metabolism. Curr. Opin. Plant Biol. 2011, 14, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.W.; Su, M.Q.; Wu, H.X.; Zhou, Y.G.; Wang, S.B. Analysis of the Volatile Profile of Core Chinese Mango Germplasm by Headspace Solid-Phase Microextraction Coupled with Gas Chromatography-Mass Spectrometry. Molecules 2018, 23, 1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burzynski-Chang, E.A.; Ryona, I.; Reisch, B.I.; Gonda, I.; Foolad, M.R.; Giovannoni, J.J.; Sacks, G.L. HS-SPME-GC-MS Analyses of Volatiles in Plant Populations-Quantitating Compound × Individual Matrix Effects. Molecules 2018, 23, 2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diez-Simon, C.; Mumm, R.; Hall, R.D. Mass spectrometry-based metabolomics of volatiles as a new tool for understanding aroma and flavour chemistry in processed food products. Metabolomics 2019, 15, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjerrum, J.T. Metabonomics; Springer: New York, NY, USA, 2015. [Google Scholar]
- Goulas, V.; Minas, I.S.; Kourdoulas, P.M.; Lazaridou, A.; Molassiotis, A.N.; Gerothanassis, I.P.; Manganaris, G.A. 1H NMR Metabolic Fingerprinting to Probe Temporal Postharvest Changes on Qualitative Attributes and Phytochemical Profile of Sweet Cherry Fruit. Front. Plant Sci. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Capitani, D.; Sobolev, A.P.; Tomassini, A.; Sciubba, F.; De Salvador, F.R.; Mannina, L.; Delfini, M. Peach Fruit: Metabolic Comparative Analysis of Two Varieties with Different Resistances to Insect Attacks by NMR Spectroscopy. J. Agric. Food Chem. 2013, 61, 1718–1726. [Google Scholar] [CrossRef]
- Mohammed, J.; Belisle, C.E.; Wang, S.; Itle, R.A.; Adhikari, K.; Chavez, D.J. Volatile Profile Characterization of Commercial Peach (Prunus persica) Cultivars Grown in Georgia, USA. Horticulturae 2021, 7, 516. [Google Scholar] [CrossRef]
- Yunwei, N.; Jianming, D.; Zuobing, X.; Jiancai, Z. Characterization of the major aroma-active compounds in peach (Prunus persica L. Batsch) by gas chromatography–olfactometry, flame photometric detection and molecular sensory science approaches. Food Res. Int. 2021, 147, 110457. [Google Scholar]
- Spadafora, N.D.; Cocetta, G.; Cavaiuolo, M.; Bulgari, R.; Dhorajiwala, R.; Ferrante, A.; Spinardi, A.; Rogers, H.J.; Müller, C.T. A complex interaction between preharvest and post-harvest factors determines fresh-cut melon quality and aroma. Sci. Rep. 2019, 9, 2745. [Google Scholar] [CrossRef] [PubMed]
- Malorni, L.; Martignetti, A.; Cozzolino, R. Volatile compound profiles by HS GCMS for the evaluation of postharvest conditions of a peach cultivar. Ann. Chromatogr. Sep. Tech. 2015, 1, 1007. [Google Scholar] [CrossRef]
- Jung, S.; Staton, M.; Lee, T.; Blenda, A.; Svancara, R.; Abbott, A.; Main, D. GDR (Genome Database for Rosaceae): Integrated webdatabase for Rosaceae genomics and genetics data. Nucleic. Acids Res. 2008, 36, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Main, D. Genomics and bioinformatics resources for translational science in Rosaceae. Plant Biotechnol. Rep. 2014, 8, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Shulaev, V.; Korban, S.S.; Sosinski, B.; Abbott, A.G.; Aldwinckle, H.S.; Folta, K.M.; Iezzoni, A.; Main, D.; Arús, P.; Dandekar, A.M.; et al. Multiple models for Rosaceae genomics. Plant Physiol. 2008, 147, 985–1003. [Google Scholar] [CrossRef] [Green Version]
- Bonghi, C.; Manganaris, G.A. Omics Technologies: Tools for Food Science. In Systems Biology Approaches Reveal New Insights into the Molecular Mechanisms Regulating Flesh Fruit Quality; CRC Press: New York, NY, USA, 2012; p. 25. [Google Scholar]
- van Dijk, E.L.; Jaszczyszyn, Y.; Naquin, D.; Thermes, C. The third revolution in sequencing technology. Trends Genet. 2018, 34, 666–681. [Google Scholar] [CrossRef]
- Amarasinghe, S.L.; Su, S.; Dong, X.; Zappia, L.; Ritchie, M.E.; Gouil, Q. Opportunities and challenges in long-read sequencing data analysis. Genome Biol. 2020, 21, 30. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, S.; Gu, C.; Zhou, Y.; Zhou, H.; Ma, J.; Jun Cheng, J.; Han, Y. Deep RNA-Seq uncovers the peach transcriptome landscape. Plant Mol. Biol. 2013, 83, 365–377. [Google Scholar] [CrossRef]
- Lurie, S. Genomic and transcriptomic studies on chilling injury in peach and nectarine. Postharvest Biol. Technol. 2021, 174, 111444. [Google Scholar] [CrossRef]
- Horn, R.; Lecouls, A.C.; Callahan, A.; Dandekar, A.; Garay, L.; McCord, P.; Howad, W.; Chan, H.; Verde, I.; Main, D.; et al. Candidate gene database and transcript map for peach, a model species for fruit trees. Theor. Appl. Genet. 2005, 110, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Verde, I.; Abbott, A.G.; Scalabrin, S.; Jung, S.; Shu, S.; Marroni, F.; Zhebentyayeva, T.; Dettori, M.T.; Grimwood, J.; Cattonaro, F.; et al. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution. Nat. Genetics. 2013, 45, 487–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pech, J.C.; Latché, A. Contribution of genomics to postharvest biology. Stewart Postharvest Rev. 2013, 9, 1–6. [Google Scholar]
- Pech, J.C.; Purgatto, E.; Girardi, C.L.; Rombaldi, C.V.; Latché, A. Current challenges in postharvest biology of fruit ripening. Curr. Agric. Sci. Technol. 2013, 19, 1–18. [Google Scholar]
- Nuñez-Lillo, G.; Cifuentes-Esquivel, A.; Troggio, M.; Micheletti, D.; Infante, R.; Campos Vargas, R.; Orellana, A.; Blanco Herrera, F.; Meneses, C. Identification of candidate genes associated with mealiness and maturity date in peach [Prunus persica (L.) Batsch] using QTL analysis and deep sequencing. Tree Genet. Genomes 2015, 11, 86. [Google Scholar] [CrossRef]
- Martínez-García, P.J.; Parfitt, D.E.; Ogundiwin, E.A.; Fass, J.; Chan, H.M.; Ahmad, R.; Lurie, S.; Dandekar, A.; Gradziel, T.M.; Crisosto, C.H. High density SNP mapping and QTL analysis for fruit quality characteristics in peach (Prunus persica L.). Tree Genet. Genomes 2013, 9, 19–36. [Google Scholar] [CrossRef]
- Cao, K.; Zhou, Z.; Wang, Q.; Guo, J.; Zhao, P.; Zhu, G.; Fang, W.; Chen, C.; Wang, X.; Wang, X.; et al. Genome-wide association study of 12 agronomic traits in peach. Nat. Commun. 2016, 7, 13246. [Google Scholar] [CrossRef] [Green Version]
- Akagi, T.; Hanada, T.; Yaegaki, H.; Gradziel, T.M.; Tao, R. Genome-wide view of genetic diversity reveals paths of selection and cultivar differentiation in peach domestication. DNA Res. 2016, 23, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Cao, K.; Deng, C.; Li, Y.; Zhu, G.; Fang, W.; Chen, C.; Wang, X.; Wu, J.; Guan, L.; et al. An integrated peach genome structural variation map uncovers genes associated with fruit traits. Genome. Biol. 2020, 21, 258. [Google Scholar] [CrossRef]
- ESTree Consortium. Development of an oligo-based microarray (μPEACH 1.0) for genomics studies in peach fruit. Acta Hort. 2005, 682, 263–268. [Google Scholar]
- González-Agüero, M.; Pavez, L.; Ibáñez, F.; Pacheco, I.; Campos-Vargas, R.; Meisel, L.A.; Orellana, A.; Retamales, J.; Silva, H.; González, M.; et al. Identification of woolliness response genes in peach fruit after post-harvest treatments. J. Exp. Bot. 2008, 59, 1973–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granell, A.; Pons, C.; Marti, C.; Forment, J.; Royo, C.; Gradziel, T.M.; Peace, C.P.; Ogundiwin, E.; Crisosto, C.H. Genomic approaches-Innovative tools to improve quality of fresh cut produce. Acta Hort. 2007, 746, 203–211. [Google Scholar] [CrossRef]
- Ogundiwin, E.A.; Martì, C.; Forment, J.; Pons, C.; Granell, A.; Gradziel, T.M.; Peace, C.P.; Crisosto, C.H. Development of ChillPeach genomic tools and identification of cold-responsive genes in peach fruit. Plant Mol. Biol. 2008, 68, 379–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirona, R.; Vecchietti, A.; Lazzari, B.; Caprera, A.; Malinverni, R.; Consolandi, C.; Severgnini, M.; De Bellis, G.; Chietera, G.; Rossini, L.; et al. Expression profiling of genes involved in the formation of aroma in two peach genotypes. Plant Biol. 2013, 15, 443–451. [Google Scholar] [CrossRef]
- Pan, H.F.; Sheng, Y.; Gao, Z.H.; Chen, H.L.; Qi, Y.J.; Yi, X.K.; Qin, G.H.; Zhang, J.Y. Transcriptome analysis of peach (Prunus persica L. Batsch) during the late stage of fruit ripening. Genet. Mol. Res. 2016, 15, 10. [Google Scholar] [CrossRef]
- Sanhueza, D.; Vizoso, P.; Balic, I.; Campos-Vargas, R.; Meneses, C. Transcriptomic analysis of fruit stored under cold conditions using controlled atmosphere in Prunus persica cv. Red Pearl. Front. Plant Sci. 2015, 6, 788. [Google Scholar] [CrossRef]
- Wang, X.; Wang, B.; Chen, K.; Tian, Z.; Long, C.; Ma, B. Comparative transcriptomic analysis of peach fruits in common and high temperature conditions. Authorea 2020, 17. [Google Scholar] [CrossRef]
- Choi, H.R.; Jeong, M.J.; Baek, M.W.; Choi, J.H.; Lee, H.C.; Jeong, C.S.; Tilahun, S. Transcriptome Analysis of Pre-Storage 1-MCP and High CO2-Treated ‘Madoka’ Peach Fruit Explains the Reduction in Chilling Injury and Improvement of Storage Period by Delaying Ripening. Int. J. Mol. Sci. 2021, 22, 4437. [Google Scholar] [CrossRef]
- Zheng, B.; Zhao, L.; Jiang, X.; Cherono, S.; Liu, J.; Ogutu, C.; Ntini, C.; Zhang, X.; Han, Y. Assessment of organic acid accumulation and its related genes in peach. Food Chem. 2021, 334, 127567. [Google Scholar] [CrossRef]
- Yu, Y.; Guan, J.; Xu, Y.; Ren, F.; Zhang, Z.; Yan, J.; Fu, J.; Guo, J.; Shen, Z.; Zhao, J.; et al. Population-scale peach genome analyses unravel selection patterns and biochemical basis underlying fruit flavor. Nat. Commun. 2021, 12, 3604. [Google Scholar] [CrossRef]
- Tikunov, Y.; Lommen, A.; de Vos, C.H.; Verhoeven, H.A.; Bino, R.J.; Hall, R.D.; Bovy, A.G. A Novel Approach for Nontargeted Data Analysis for Metabolomics. Large-Scale Profiling of Tomato Fruit Volatiles. Plant Physiol. 2005, 139, 1125–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steuer, R. Review: On the analysis and interpretation of correlations in metabolomic data. Brief. Bioinform. 2006, 7, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Ursem, R.; Tikunov, Y.; Bovy, A.; van Berloo, R.; van Eeuwijk, F. A correlation network approach to metabolic data analysis for tomato fruits. Euphytica 2008, 161, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, V.A.; Osorio, S.; Borsani, J.; Lauxmann, M.A.; Bustamante, C.A.; Budde, C.O.; Andreo, C.S.; Lara, M.V.; Fernie, A.R.; Drincovich, M.F. Metabolic profiling during peach fruit development and ripening reveals the metabolic networks that underpin each developmental stage. Plant Physiol. 2011, 157, 1696–1710. [Google Scholar] [CrossRef] [Green Version]
- Xi, W.P.; Xi, W.P.; Zhang, B.O.; Liang, L.I.; Shen, J.Y.; Wei, W.W.; Xu, C.J.; Allan, A.C.; Ferguson, I.B.; Chen, K.S. Postharvest temperature influences volatile lactone production via regulation of acyl-CoA oxidases in peach fruit. Plant Cell Environ. 2012, 35, 534–545. [Google Scholar] [CrossRef]
- Monti, L.L.; Bustamante, C.A.; Osorio, S.; Gabilondo, J.; Borsani, J.; Lauxmann, M.A.; Maulión, E.; Valentini, G.; Budde, C.O.; Fernie, A.R.; et al. Metabolic profiling of a range of peach fruit varieties reveals high metabolic diversity and commonalities and differences during ripening. Food Chem. 2016, 1, 879–888. [Google Scholar] [CrossRef]
- Brizzolara, S.; Tonutti, P. The effect of cold storage on volatile organic compounds (VOCs) emitted from intact peach fruit. Acta Hortic. 2019, 1256, 151–156. [Google Scholar] [CrossRef]
- Pott, D.M.; Vallarino, J.G.; Osorio, S. Metabolite Changes during Postharvest Storage: Effects on Fruit Quality Traits. Metabolites 2020, 10, 187. [Google Scholar] [CrossRef]
- Lauxmann, M.A.; Borsani, J.; Osorio, S.; Lombardo, V.A.; Budde, C.O.; Bustamante, C.A.; Monti, L.L.; Andreo, C.S.; Fernie, A.R.; Drincovich, M.F.; et al. Deciphering the metabolic pathways influencing heat and cold responses during post-harvest physiology of peach fruit. Plant Cell Environ. 2014, 37, 601–616. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Liu, H.R.; Gao, J.; Huang, Y.J.; Zhang, B.; Chen, K.S. Two ω-3 FADs Are Associated with Peach Fruit Volatile Formation. Int. J. Mol. Sci. 2016, 17, 464. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Shen, W.; Zheng, Q.; Fowler, D.B.; Zou, J. Adjustments of lipid pathways in plant adaptation to temperature stress. Plant Signal Behav. 2016, 11, e1058461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, B.; Yu, M.; Zhang, B.; Xu, J.; Ma, R. Differences in PpAAT1 Activity in High- and Low-Aroma Peach Varieties Affect γ-Decalactone Production. Plant Physiol. 2020, 182, 2065–2080. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.Z.; Peng, B.; Gu, Z.X.; Tang, M.L.; Li, B.; Liang, M.X.; Wang, L.M.; Guo, X.T.; Wang, J.P.; Sha, Y.F.; et al. Site-directed mutagenesis identified the key active site residues of alcohol acyltransferase PpAAT1 responsible for aroma biosynthesis in peach fruits. Hortic. Res. 2021, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Xie, K.; Duan, W.; Zhu, Y.; Liu, M.; Chen, K.; Klee, H.; Zhang, B. Peach Carboxylesterase PpCXE1 Is Associated with Catabolism of Volatile Esters. J. Agric. Food Chem. 2019, 67, 5189–5196. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cao, X.; Liu, X.; Xin, R.; Wang, J.; Gao, J.; Wu, B.; Gao, L.; Xu, C.; Zhang, B.; et al. UV-B irradiation differentially regulates terpene synthases and terpene content of peach. Plant Cell Environ. 2017, 40, 2261–2275. [Google Scholar] [CrossRef]
- Carrari, F.; Baxter, C.; Usadel, B.; Urbanczyk-Wochniak, E.; Zanor, M.I.; Nunes-Nesi, A.; Nikiforova, V.; Centero, D.; Ratzka, A.; Pauly, M.; et al. Integrated analysis of metabolite and transcript levels reveals the metabolic shifts that underlie tomato fruit development and highlight regulatory aspects of metabolic network behavior. Plant Physiol. 2006, 142, 1380–1396. [Google Scholar] [CrossRef] [Green Version]
- Mounet, F.; Moing, A.; Garcia, V.; Petit, J.; Maucourt, M.; Deborde, C.; Bernillon, S.; Le Gall, G.; Colquhoun, I.; Defernez, M.; et al. Gene and metabolite regulatory network analysis of early developing fruit tissues highlights new candidate genes for the control of tomato fruit composition and development. Plant Physiol. 2009, 149, 1505–1528. [Google Scholar] [CrossRef] [Green Version]
- Osorio, S.; Alba, R.; Damasceno, C.M.B.; Lopez-Casado, G.; Lohse, M.; Zanor, M.I.; Tohge, T.; Usadel, B.; Rose, J.K.; Fei, Z.; et al. Systems biology of tomato fruit development: Combined transcript, protein, and metabolite analysis of tomato transcription factor (nor, rin) and ethylene receptor (Nr) mutants reveals novel regulatory interactions. Plant Physiol. 2011, 157, 405–425. [Google Scholar] [CrossRef] [Green Version]
- Zamboni, A.; Carli, M.D.; Guzzo, F.; Stocchero, M.; Zenoni, S.; Ferrarini, A.; Tononi, P.; Toffali, K.; Desiderio, A.; Lilley, K.S.; et al. Identification of putative stage-specific grapevine berry biomarkers and omics data integration into networks. Plant Physiol. 2010, 154, 1439–1459. [Google Scholar] [CrossRef] [Green Version]
- Savoi, S.; Wong, D.C.J.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Transcriptome and metabolite profiling reveals that prolonged drought modulates the phenylpropanoid and terpenoid pathway in white grapes (Vitis vinifera L.). BMC Plant Biol. 2016, 16, 67. [Google Scholar] [CrossRef] [Green Version]
- Savoi, S.; Wong, D.C.J.; Degu, A.; Herrera, J.C.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Multi-Omics and Integrated Network Analyses Reveal New Insights into the Systems Relationships between Metabolites, Structural Genes, and Transcriptional Regulators in Developing Grape Berries (Vitis vinifera L.) Exposed to Water Deficit. Front. Plant Sci. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eduardo, I.; Chietera, G.; Raul Pirona, R.; Pacheco, I.; Troggio, M.; Banchi, E.; Bassi, D.; Rossini, L.; Vecchietti, A.; Pozzi, C. Genetic dissection of aroma volatile compounds from the essential oil of peach fruit: QTL analysis and identification of candidate genes using dense SNP maps. Tree Genet. Genomes 2013, 9, 89–204. [Google Scholar] [CrossRef]
- Salazar, J.A.; Ruiz, D.; Campoy, J.A.; Sánchez-Pérez, R.; Crisosto, C.H.; Martínez-García, P.J.; Blenda, A.; Jung, S.; Main, D.; Martínez-Gómez, P.; et al. Quantitative Trait Loci (QTL) and Mendelian Trait Loci (MTL) analysis in Prunus: A breeding perspective and beyond. Plant Mol. Biol. Rep. 2014, 32, 1–18. [Google Scholar] [CrossRef]
- Sánchez, G.; Martínez, J.; Romeu, J.; García, J.; Monforte, A.J.; Badenes, M.L.; Granell, A. The peach volatilome modularity is reflected at the genetic and environmental response levels in a QTL mapping population. BMC Plant Biol. 2014, 14, 137. [Google Scholar] [CrossRef] [Green Version]
- Fresnedo-Ramírez, J.; Bink, M.C.A.M.; van de Weg, E.; Famula, T.R.; Crisosto, C.H.; Frett, T.J.; Gasic, K.; Peace, C.P.; Gradzie, T.M. QTL mapping of pomological traits in peach and related species breeding germplasm. Mol. Breed. 2015, 35, 166. [Google Scholar] [CrossRef]
- Carrasco-Valenzuela, T.; Muñoz-Espinoza, C.; Riveros, A.; Pedreschi, R.; Arús, P.; Campos-Vargas, R.; Meneses, C. Expression QTL (eQTLs) Analyses Reveal Candidate Genes Associated with Fruit Flesh Softening Rate in Peach [Prunus persica (L.) Batsch]. Front. Plant Sci. 2019, 10, 1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Cao, X.; Liu, H.; Zhu, C.; Klee, H.; Zhang, B.; Chen, K. UDP-glucosyltransferase PpUGT85A2 controls volatile glycosylation in peach. J. Exp Bot. 2019, 70, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Han, S.; Jiang, L.; Yu, M.; Ma, R.; Yu, Z. 1-MCP treatment affects peach fruit aroma metabolism as revealed by transcriptomics and metabolite analyses. Food Res. Int. 2019, 122, 573–584. [Google Scholar] [CrossRef]
- Scossa, F.; Alseekh, S.; Fernie, A.R. Integrating multi-omics data for crop improvement. J. Plant Physiol. 2021, 257, 153352. [Google Scholar] [CrossRef]
- Salazar, J.A.; Pacheco, I.; Shinya, P.; Zapata, P.; Silva, C.; Ruiz, D.; Martínez-Gómez, P.; Infante, R. Genotyping by Sequencing for SNP-Based Linkage Analysis and Identification of QTLs Linked to Fruit Quality Traits in Japanese Plum (Prunus salicina Lindl.). Front. Plant Sci. 2017, 8, 476. [Google Scholar] [CrossRef] [Green Version]


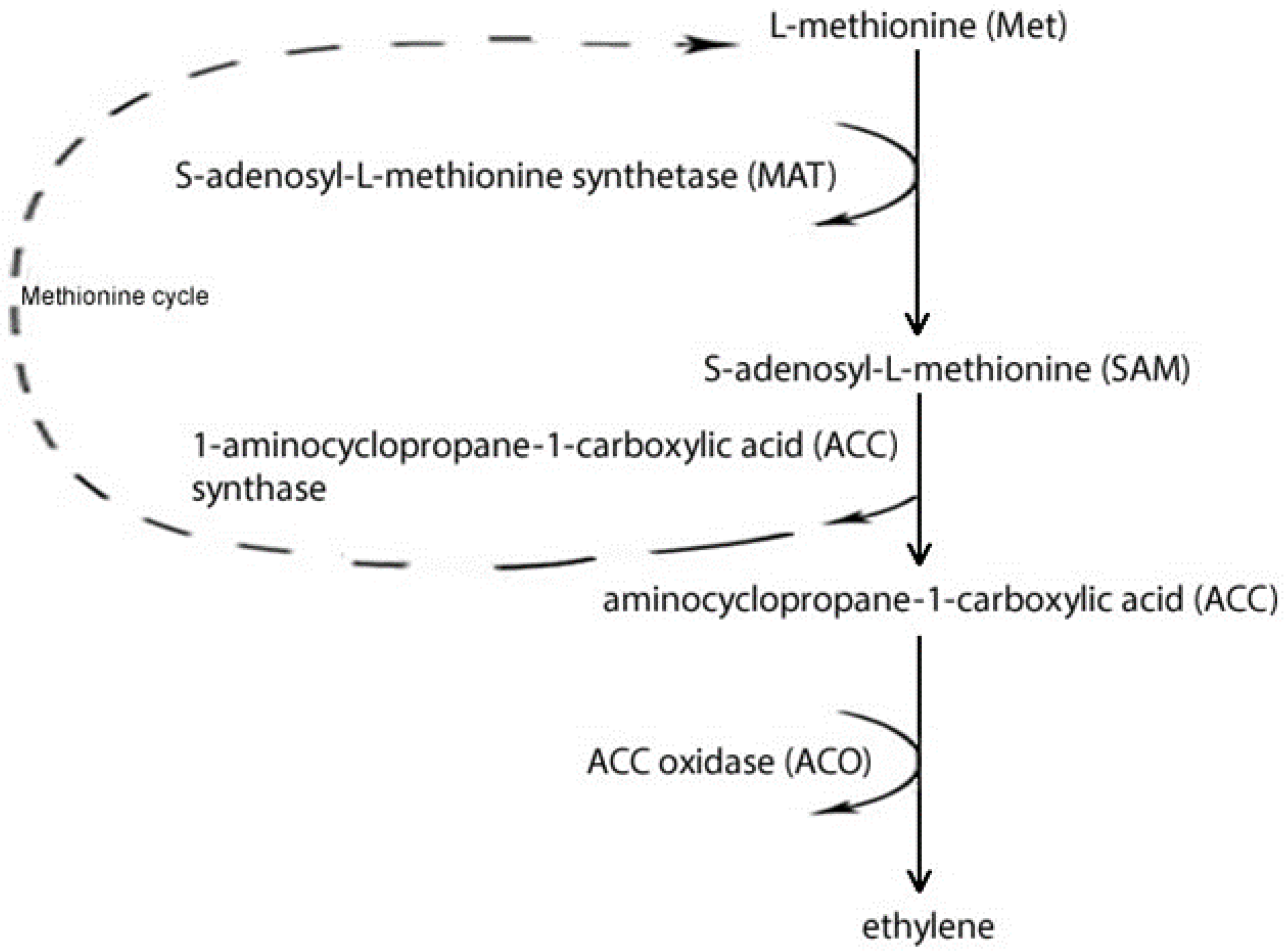{kind=link}
| Abbreviations | Definition |
|---|---|
| 1-MCP | 1-Methylcyclopropene |
| AATs | Alcohol acyltransferases |
| ABA | Abscisic Acid |
| ACC | Aminocyclopropane-1-carboxylic acid |
| ADHs | Alcohol Dehydrogenases |
| AVG | Aminoethoxyvinylglycine |
| BAC | Bacterial Artificial Chromosome |
| CA | Controlled Atmosphere |
| CBF | C-Repeat Binding Factor |
| cDNA | complementary DNA |
| CI | Chilling Injury |
| CKs | Cytokinins |
| CNA | Correlation Networks Analysis |
| CRISP/Cas9 | Clustered Regularly Interspaced Short Palindromic Repeats/CRISP associated protein 9 |
| CS | Cold Storage |
| DEG | Differentially Expressed Gene |
| EST | Expressed Sequence Tag |
| GAs | Gibberellins |
| GC-FPD | Gas Chromatography-Flame Photometric Detection |
| GC-MS | Gas Chromatography–Mass Spectrometry Techniques |
| GC-O | Gas Chromatography-Olfactometry |
| GWAS | Genome-Wide Association Study |
| HPLs | Hydroperoxide Lyases |
| HS-SPME | Headspace Solid-Phase Microextraction |
| Jas | Jasmonates |
| LC-MS | Liquid Chromatography–Mass Spectrometry Techniques |
| LOXs | Lipoxygenases |
| MET | L-methionine |
| MJA | Methyl Jasmonate |
| NGS | Next Genetation Sequencing |
| NMR | Nuclear Magnetic Resonance |
| QTL | Quantitative Trait Loci |
| qRT-PCR | Real Time or Quantitative PCR–Polymerase Chain Reaction |
| SAM | S-Adenosyl Methionine |
| SMRT | Single Molecule Real-Time |
| SNPs | Single Nucleotide Polymorphisms |
| SVs | Structural Variants |
| TD | Thermal Desorption |
| UV | Ultraviolet |
| VOC | Volatile Organic Compounds |
| Omic Sciences and Techniques | Post-Harvest Study * | Description | References |
|---|---|---|---|
| transcriptomics/microarray | μPEACH1.0 definition | [107] | |
| transcriptomics/microarray | Analysis of auxin and ethylene in ripening | [34] | |
| transcriptomics/microarray | CS | Identification of woolliness response | [108] |
| transcriptomics/microarray | CS | Identification of cold-responsive genes | [110] |
| metabolomics | CS | Analysis of post-harvest temperature influence on lactone production via acyl-CoA oxidases | [122] |
| metabolomics | CNA to study the effect of pre- and post-harvest treatments | [2] | |
| transcriptomics/microarray | Transcriptomic changes at different ripening stages | [111] | |
| genomics | Peach v2.0.a1 definition | [99] | |
| transcriptomics/RNA-SEQ | RNA-Seq and the peach transcriptome landscape | [95,96] | |
| genomics/QTL | Analysis of genes responsible for fruit quality traits ** | [100] | |
| genomics/QTL/SNP | CS | High resolution SNP mapping and QTL analysis for fruit quality traits | [103] |
| metabolomics | CS | Metabolic pathways involved in heat and cold responses in post-harvest conditions | [126] |
| genomics/QTL | CS | Identification of putative genes associated with mealiness and ripeness in peach | [102] |
| genomics/QTL | QTL mapping of peach traits | [142] | |
| transcriptomics/RNA-SEQ | CA, CS | Study of gene expression in post-harvest conditions | [113] |
| metabolomics | Analysis of compounds important for organoleptic properties during post-harvest ripening | [123] | |
| transcriptomics/RNA-SEQ | Transcriptome analysis during the late stage of ripening | [112] | |
| metabolomics | Correlation among FADs and VOCs | [127] | |
| metabolomics | CS | Metabolic responses to low temperature of different cultivars | [76] |
| metabolomics | CS | Study of effect of CS on intact peach VOCs | [124] |
| metabolomics | CS | Fruit volatilome profiling and gene expression analyses amongst peach cultivars | [7] |
| metabolomics | Identification of key metabolites involved in organoleptic and health-benefit traits ** | [125] | |
| genomics/SVs | Genes associated with fruit traits and VOCs | [106] | |
| transcriptomics/RNA-SEQ | Analysis at high temperature conditions during fruit ripening and post-harvest | [114] | |
| transcriptomics/RNA-SEQ | CS, 1-MCP | Analysis of gene expression changes in pre-storage treatments during CS | [115] |
| transcriptomics/RNA-SEQ | UV-C | UV-C treatment in post-harvest | [28] |
| genomics/mutagenesis | Identification of the key gene regions of PpAAT1 responsible for peach aroma | [130] |
| Omic Science and Techniques | Post-Harvest Study * | Description | References |
|---|---|---|---|
| metabolomics, transcriptomics/microarray | Integrated omics analysis to study tomato development and ripening ** | [133] | |
| metabolomics, transcriptomics/microarray | Genetic and Metabolic Regulatory Network Analysis in Tomato development ** | [134] | |
| genomics, EST resources, metabolomics, qRT-PCR | Post-harvest ripening at 20 °C | Study about aroma and fatty acids pathway | [5] |
| metabolomics, transcriptomics | Survey of molecular regulation in fruit ripening ** | [135] | |
| genomics, transcriptomics, metabolomics | Survey on ‘omics’ methods to investigate fresh fruit quality ** | [92] | |
| genomics/QTL/SNP, metabolomics | Identification of candidate genes and QTL analysis through SNP maps | [139] | |
| metabolomics, transcriptomics/microarray | Identification of new candidate genes to impact aroma volatiles | [70] | |
| genomics/QTL, metabolomics | QTL analysis in Prunus: a breeding perspective | [140] | |
| genomics/QTL, metabolomics | QTL analysis and individuation of new loci | [141] | |
| metabolomics, transcriptomics/qRT-PCR | GWAS to study 12 peach agronomic traits | [104] | |
| metabolomics, transcriptomics/RNA-SEQ | Relationships among metabolites, genes, and transcription factors/regulators in developing grape berries ** | [137,138] | |
| metabolomics, transcriptomics/qRT-PCR | CS | Exploring priming responses to cold stress | [19] |
| metabolomics, transcriptomics/RNA-SEQ | UV-B | Regulation of genes encoding terpene synthases and terpene levels | [132] |
| metabolomics, transcriptomics/RNA-SEQ | 1-MCP UV-B | PpUGT85A2 regulates volatile glycosylation in peach | [144] |
| genomics/QTL, transcriptomics/RNA-SEQ | Expression QTL analyses and individuation of genes associated with fruit softening rate | [143] | |
| metabolomics, transcriptomics/RNA-SEQ | MJA, UV-B | Correlation between PpCXE1 and catabolism of volatile esters | [131] |
| metabolomics, transcriptomics/RNA-SEQ | 1-MCP | 1-MCP treatment affects peach fruit aroma | [145] |
| genomics/CRISPR/Cas9, metabolomics, transcriptomics/RNA-SEQ | Correlation between PpAAT1 activity and Aroma peach varieties | [129] | |
| genomics, metabolomics, transcriptomics | Review on multi-omics applications with a breeding perspective-Prunus species | [33] | |
| genomic, transcriptomic | CS and CI | Review about genomic and transcriptomic studies to find QTLs and genes responsible for CI symptoms | [97] |
| metabolomics, transcriptomics/RNA-SEQ | Organic acid accumulation and related genes | [116] | |
| genomics/GWAS/SNPs, transcriptomics/RNA-SEQ | Study of genes associated with aroma profile | [117] | |
| genomics, metabolomics, transcriptomics | Survey on multi-omics studies for crop improvement ** | [146] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirangelo, T.M.; Rogers, H.J.; Spadafora, N.D. Multi-Omic Approaches to Investigate Molecular Mechanisms in Peach Post-Harvest Ripening. Agriculture 2022, 12, 553. https://doi.org/10.3390/agriculture12040553
Sirangelo TM, Rogers HJ, Spadafora ND. Multi-Omic Approaches to Investigate Molecular Mechanisms in Peach Post-Harvest Ripening. Agriculture. 2022; 12(4):553. https://doi.org/10.3390/agriculture12040553
Chicago/Turabian StyleSirangelo, Tiziana M., Hilary J. Rogers, and Natasha D. Spadafora. 2022. "Multi-Omic Approaches to Investigate Molecular Mechanisms in Peach Post-Harvest Ripening" Agriculture 12, no. 4: 553. https://doi.org/10.3390/agriculture12040553