
The Comprehensive Effects of Rhizophagus intraradices and P on Root System Architecture and P Transportation in Citrus limon L.



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Culture
2.2. Experimental Design
2.3. Determination of Physiological Parameters
2.4. qRT-PCR for LR and Phosphatase Genes
2.5. Statistical Analysis
3. Results
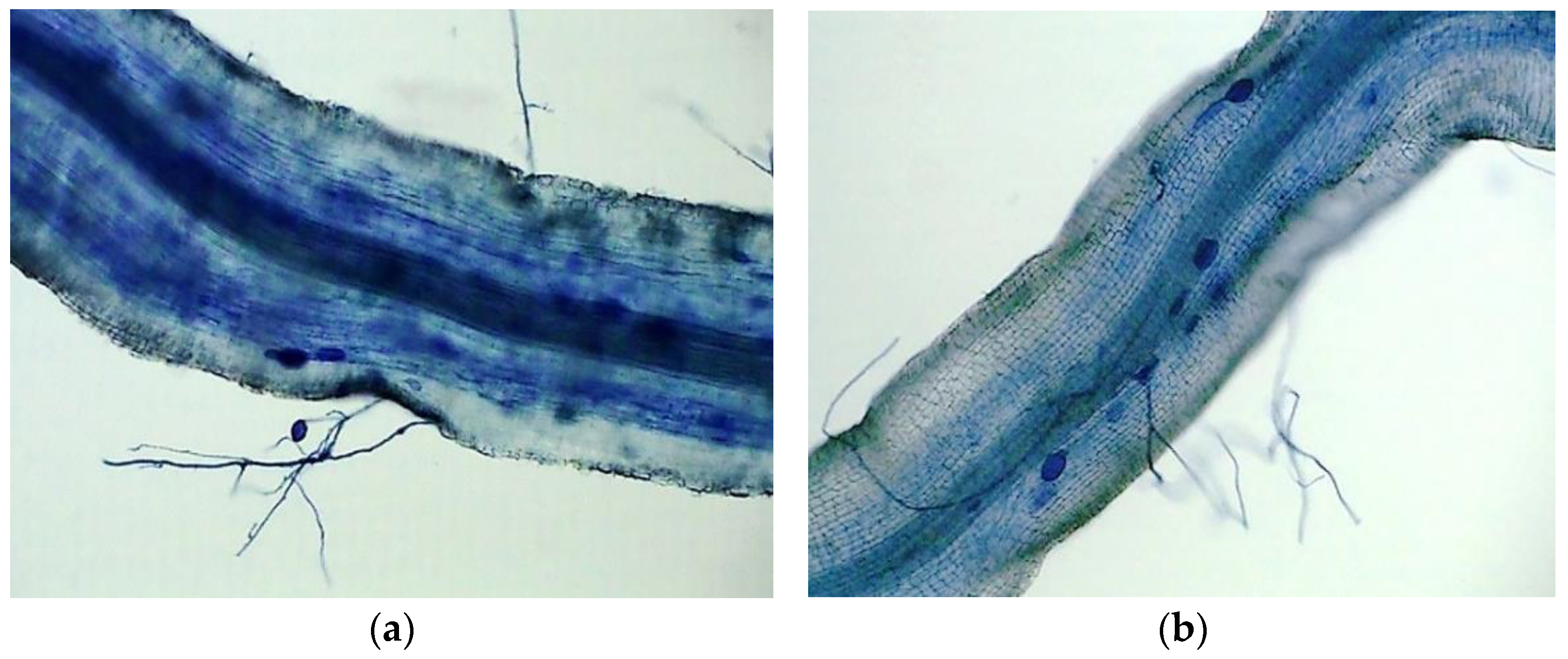
3.1. Changes in Mycorrhizal Development and Plant Growth Performance
3.2. Changes in Root Morphology
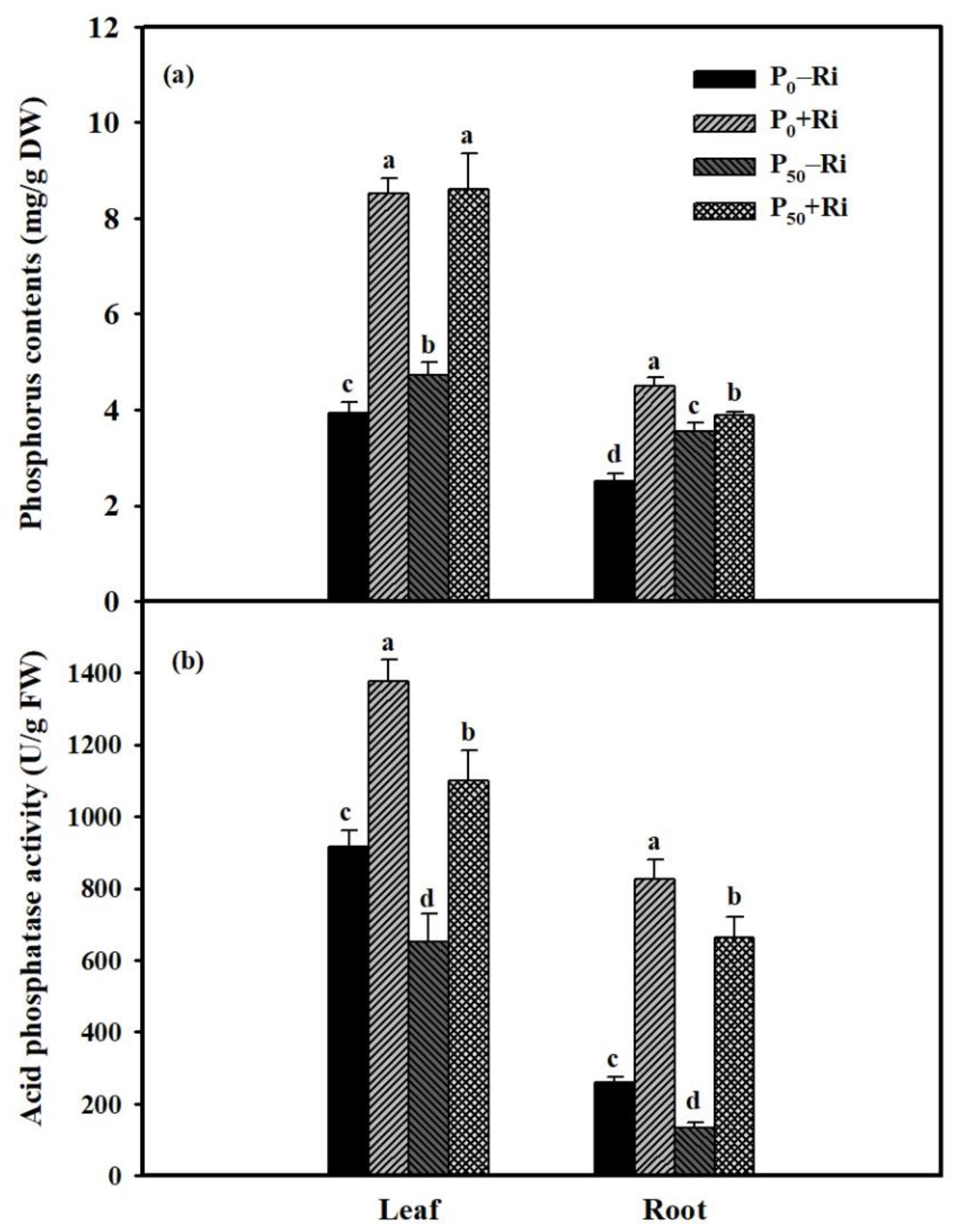
3.3. Changes in Root P Content and Phosphatase Activity
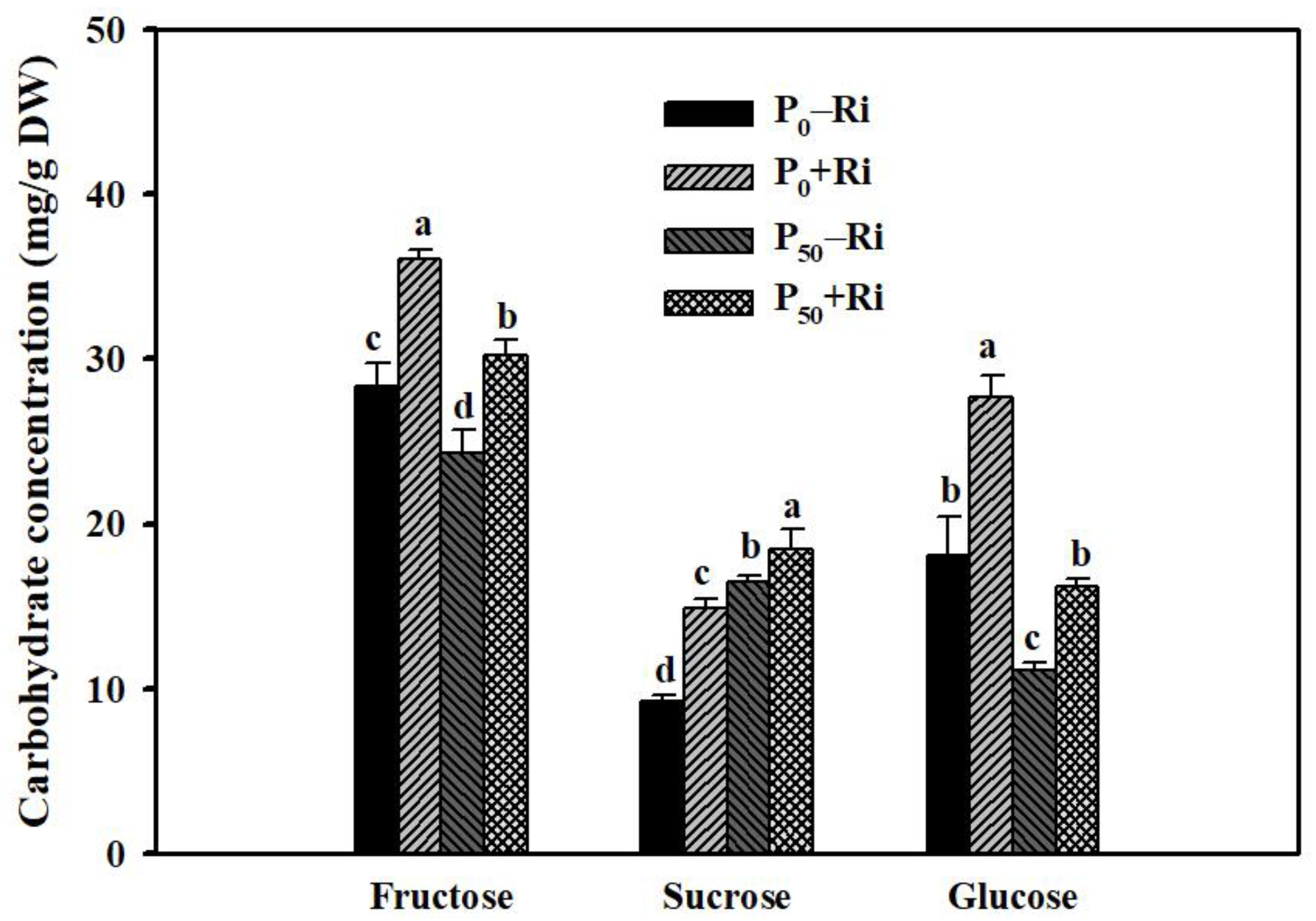
3.4. Changes in Root Carbohydrate Concentrations
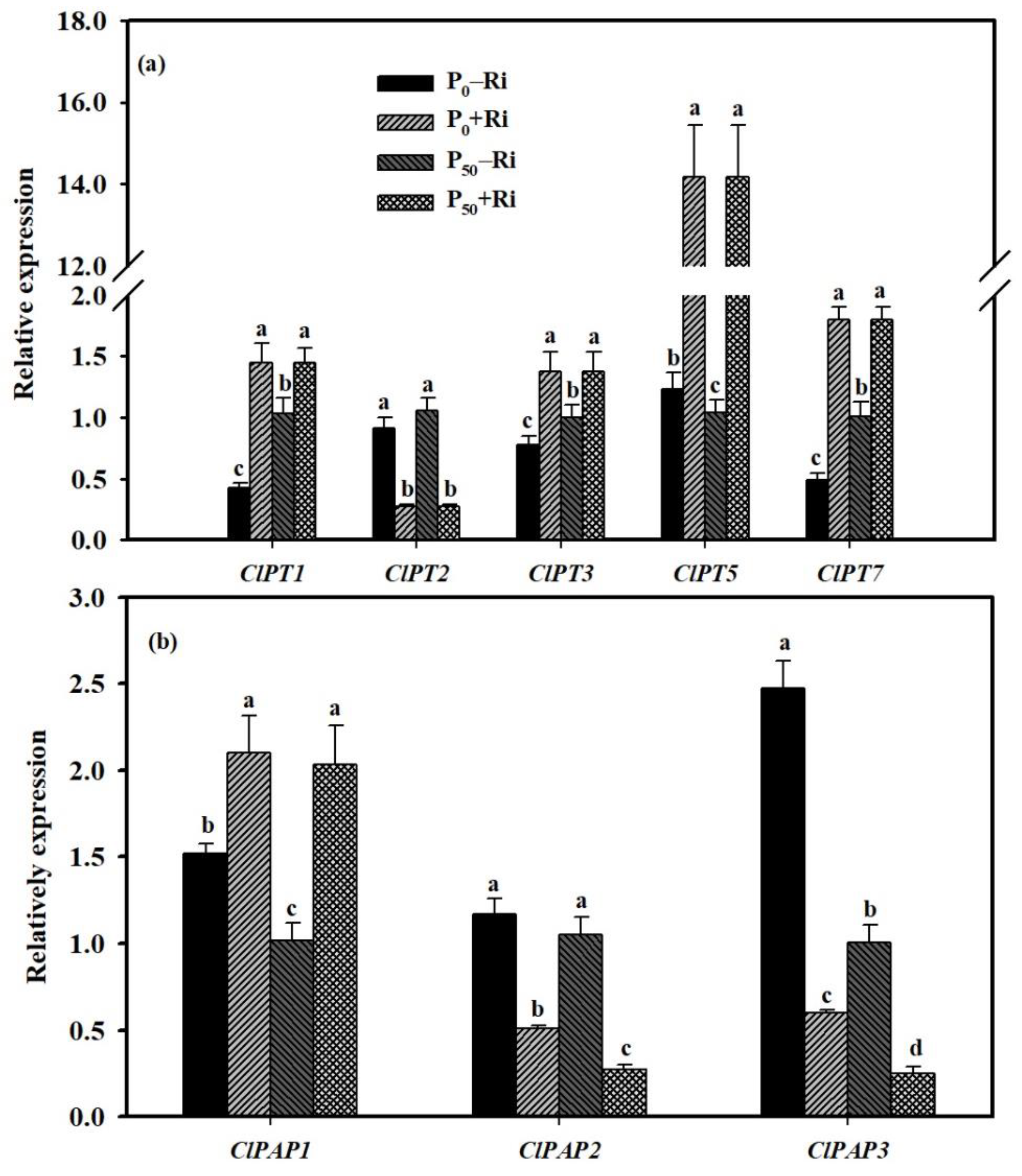
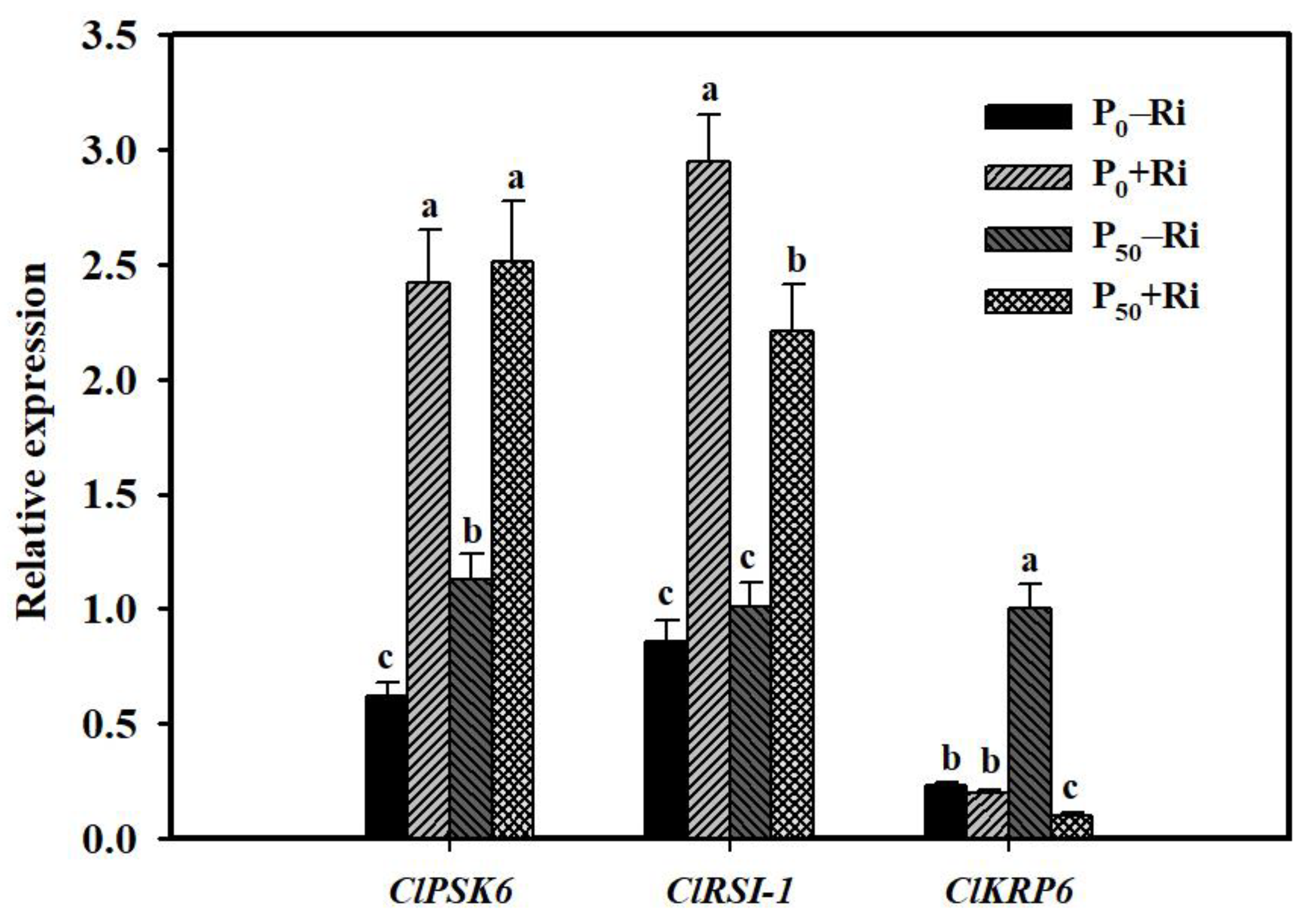
3.5. Changes in Relative Expression of Root P-Related Genes
3.6. Expression of LR-Related Genes
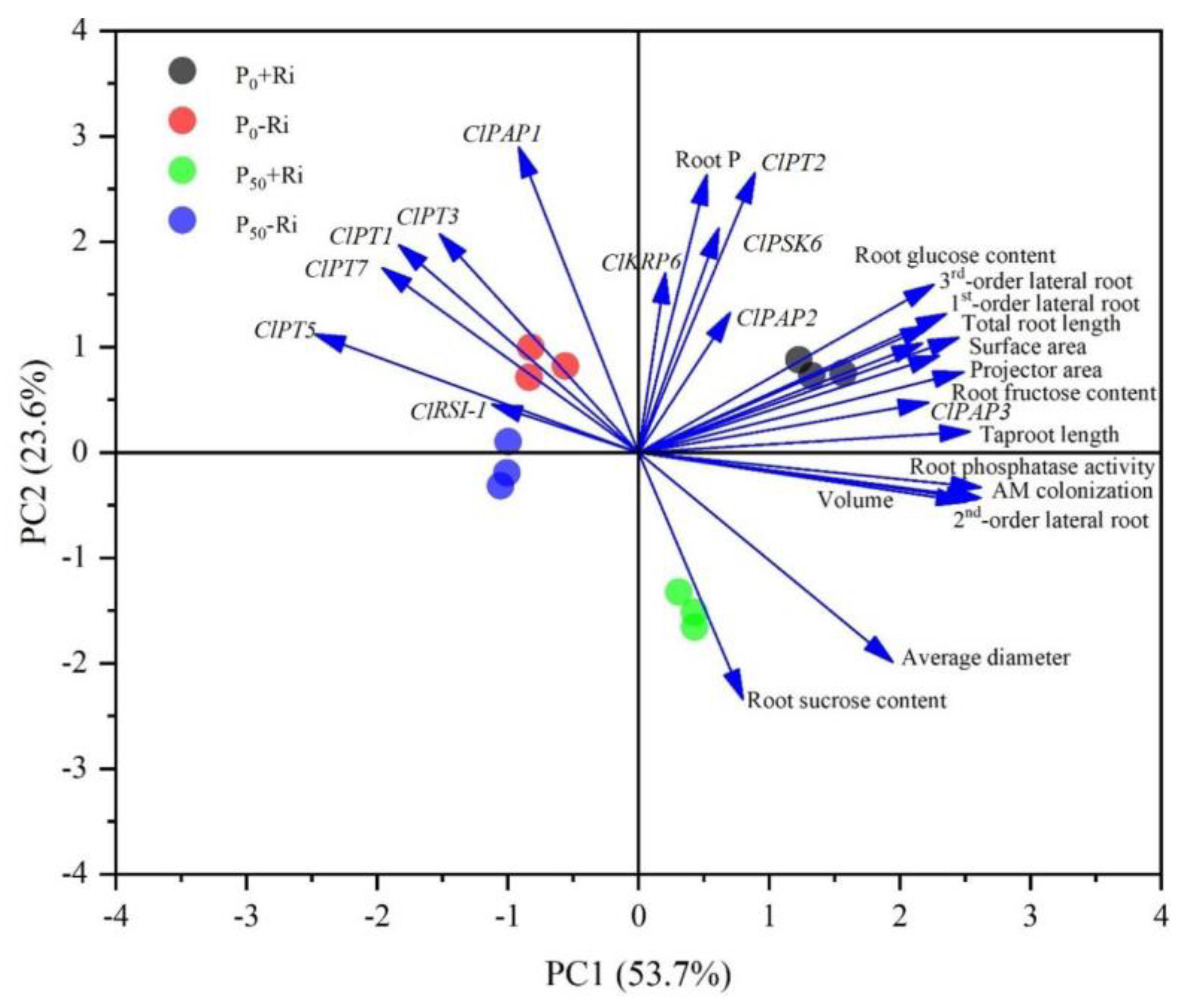
3.7. Principal Component Analysis
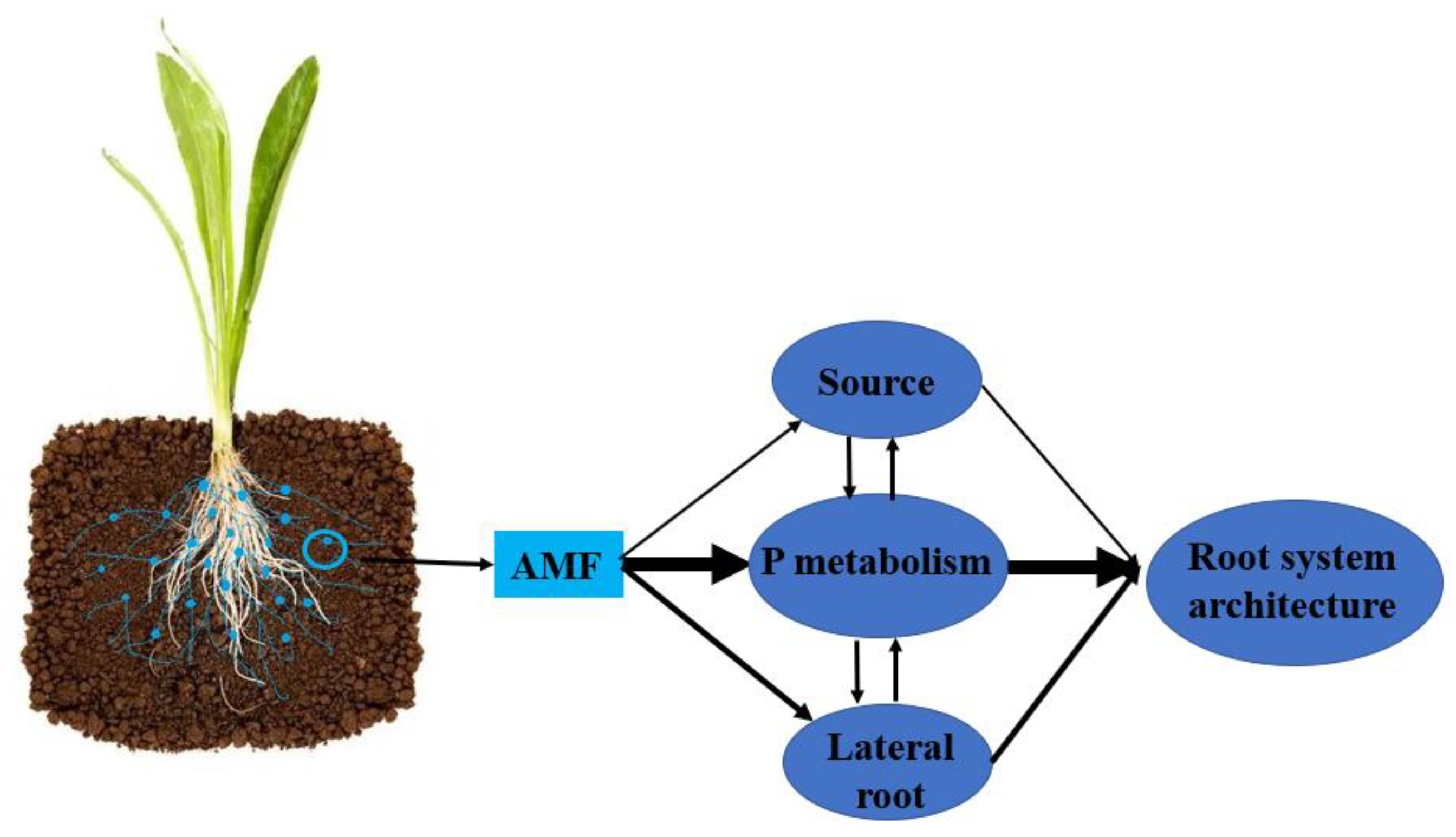
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, W.-L.; Li, J.; Zhu, H.-H.; Xu, P.-Y.; Chen, J.-Z.; Yao, Q. The differential and interactive effects of arbuscular mycorrhizal fungus and phosphorus on the lateral root formation in Poncirus trifoliata (L.). Sci. Horti. 2017, 217, 258–265. [Google Scholar] [CrossRef]
- Flavel, R.-J.; Guppy, C.-N.; Tighe, M.-K.; Watt, M.; Young, I.-M. Quantifying the response of wheat (Triticum aestivum L.) root system architecture to phosphorus in an Oxisol. Plant Soil 2014, 385, 303–310. [Google Scholar] [CrossRef]
- Funakoshi, Y.; Daimon, H.; Matsumura, A. Morphological and physiological studies on densely branched lateral roots triggered by localized phosphate in Sesbania cannabina. J. Plant Nutr. Soil Sci. 2018, 181, 336–344. [Google Scholar] [CrossRef]
- Abel, S. Phosphate scouting by root tips. Curr. Opin. Plant Biol. 2017, 39, 168–177. [Google Scholar] [CrossRef]
- Johnston, A.-E.; Poulton, P.-R.; Fixen, P.-E.; Curtin, D. Phosphorus: Its efficient use in agriculture. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 177–228. [Google Scholar] [CrossRef]
- Ros, M.-B.-H.; de Deyn, G.-B.; Koopmans, G.-F.; Oenema, O.; van Groenigen, J.-W. What root traits determine grass resistance to phosphorus deficiency in production grassland? J. Plant Nutr. Soil Sci. 2018, 181, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Wissuwa, M.; Gonzalez, D.; Watts-Williams, S.-J. The contribution of plant traits and soil microbes to phosphorus uptake from low-phosphorus soil in upland rice varieties. Plant Soil 2020, 448, 523–537. [Google Scholar] [CrossRef]
- Ma, X.-M.; Li, X.-L.; Ludewig, U. Arbuscular mycorrhizal colonisation outcompetes root hairs in maize under low phosphorus availability. Ann. Bot. 2020, 127, 155–166. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Wu, Q.-S. Responses of plant growth, root morphology, chlorophyll and indoleacetic acid to phosphorus stress in trifoliate orange. Biotechnology 2017, 16, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Habibzadeh, Y. The effects of arbuscular mycorrhizal fungi and phosphorus levels on dry matter production and root traits in cucumber (Cucumis sativus L.). Afr. J. Environ. Sci. Technol. 2015, 9, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-Y.; Wang, P.; Zhang, D.-J.; Zou, Y.-N.; Kuča, K.; Wu, Q.S. Mycorrhiza-induced change in root hair growth is associated with IAA accumulation and expression of EXPs in trifoliate orange under two P levels. Sci. Horti. 2018, 234, 227–235. [Google Scholar] [CrossRef]
- Shao, Y.-D.; Hu, X.-C.; Wu, Q.-S.; Yang, T.-Y.; Srivastava, A.-K.; Zhang, D.-J.; Gao, X.-B.; Kuča, K. Mycorrhizas promote P acquisition of tea plants through changes in root morphology and P transporter gene expression. S. Afr. Bot. 2021, 137, 455–462. [Google Scholar] [CrossRef]
- Gutjahr, C.; Casieri, L.; Paszkowski, U. Glomus intraradices induces changes in root system architecture of rice independently of common symbiosis signaling. New Phytol. 2009, 182, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; De Smet, I.; Ding, Z. Shaping a root system: Regulating lateral versus primary root growth. Trends Plant Sci. 2014, 19, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.-L.; Liang, S.-M.; Chu, X.-N.; Yang, Y.-L.; Wu, Q.-S. Mycorrhizal fungi enhance flooding tolerance of peach through inducing proline accumulation and improving root architecture. Plant Soil Environ. 2020, 66, 624–631. [Google Scholar] [CrossRef]
- Cozzolino, V.; Pigna, M.; Di Meo, V.; Caporale, A.-G.; Violante, A. Effects of arbuscular mycorrhizal inoculation and phosphorus supply on the growth of Lactuca sativa L. and arsenic and phosphorus availability in an arsenic polluted soil under non-sterile conditions. Appl. Soil Ecol. 2010, 45, 262–268. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.-Y.; Weng, B.-S.; Cai, B.-P.; Dong, Y.-R.; Yan, C.-L. Effects of arbuscular mycorrhizal inoculation and phosphorus supply on the growth and nutrient uptake of Kandelia obovata (Sheue, Liu & Yong) seedlings in autoclaved soil. Appl. Soil Ecol. 2014, 75, 162–171. [Google Scholar] [CrossRef]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Santner, A.; Pozo, J.-C.-D.; Murray, J.-A.; Estelle, M. Degradation of the cyclin-dependent kinase inhibitor KRP1 is regulated by two different ubiquitin E3 ligases. Plant J. 2008, 53, 705–716. [Google Scholar] [CrossRef]
- Zhang, Y.; Anja, P.; Caroline, M.; Liu, S.; Tai, H.; Josefine, N.; Yeh, C.-T.; Nina, O.; Christa, L.; Schnable, P.-S. The Aux/IAA gene rum1 involved in seminal and lateral root formation controls vascular patterning in maize (Zea mays L.) primary roots. J. Exp. Bot. 2015, 65, 4919–4930. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.-S.; Srivastava, A.-K.; Li, Y. Effects of mycorrhizal symbiosis on growth behavior and carbohydrate metabolism of trifoliate orange under different substrate P levels. J. Plant Growth Regul. 2015, 34, 499–508. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Liu, C.-Y.; Zhang, D.-J.; Zou, Y.-N.; He, X.-H.; Wu, Q.-H. Mycorrhiza alters the profile of root hairs in trifoliate orange. Mycorrhiza 2016, 26, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-Y.; Zhang, F.; Zhang, D.-J.; Zou, Y.-N.; Shu, B.; Wu, Q.-S. Transcriptome analysis reveals improved root hair growth in trifoliate orange seedlings by arbuscular mycorrhizal fungi. Plant Growth Regul. 2020, 95, 195–203. [Google Scholar] [CrossRef]
- Phillips, J.-M.; Hayman, D.-S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Mc Lachlan, K.-D.; Elliot, D.-E.; De Marco, D.-G.; Garran, J.-H. Leaf acid phosphatase isozymes in the diagnosis of phosphorus status in field-grown wheat. Aust. J. Agric. Res. 1987, 38, 1–13. [Google Scholar] [CrossRef]
- Yan, J.-W.; Yuan, F.-R.; Long, G.-Y.; Qin, L.; Deng, Z.-N. Selection of reference genes for quantitative real-time RT-PCR analysis in citrus. Mol. Biol. Rep. 2012, 39, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.-J.; Schmittgen, T.-D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Srivastava, A.-K.; Wu, Q.-S. Mycorrhizal fungi regulate root responses and leaf physiological activities in trifoliate orange. Not. Bot. Horti Agrobot. 2017, 45, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Pan, Q.; Chen, F.; Yan, X.; Liao, H. Effects of co-inoculation with arbuscular mycorrhizal fungi and rhizobia on soybean growth as related to root architecture and availability of N and P. Mycorrhiza 2010, 21, 173–181. [Google Scholar] [CrossRef]
- Norman, J.-R.; Atkinson, D.; Hooker, J.-E. Arbuscular mycorrhizal fungal-induced alteration to root architecture in strawberry and induced resistance to the root pathogen Phytophthora fragariae. Plant Soil 1996, 185, 191–198. [Google Scholar] [CrossRef]
- López-Bucio, J.; Hernández-Abreu, E.; Sánchez-Calderón, L.; Nieto-Jacobo, M.-F.; Simpson, J.; Herrera-Estrella, L. Phosphate availability alters architecture and causes changes in hormone sensitivity in the Arabidopsis root system. Plant Physiol. 2002, 129, 244–256. [Google Scholar] [CrossRef] [Green Version]
- Gahoonia, T.-S.; Nielsen, N.-E. Phosphorus (P) uptake and growth of a root hairless barley mutant (bald root barley, brb) and wild type in low- and high-P soils. Plant Cell Environ. 2003, 26, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Vysotskaya, L.-B.; Trekozova, A.-W.; Kudoyarova, G.-R. Effect of phosphorus starvation on hormone content and growth of barley plants. Acta Physiol. Plant. 2016, 38, 1–6. [Google Scholar] [CrossRef]
- Zhang, C.; Simpson, R.-J.; Kim, C.-M.; Warthmann, N.; Delhaize, E.; Dolan, L.; Byrne, M.-E.; Wu, Y.; Ryan, P.-R. Do longer root hairs improve phosphorus uptake? Testing the hypothesis with transgenic Brachypodium distachyon lines overexpressing endogenous RSL genes. New Phytol. 2018, 217, 1654–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Li, J.; Zhu, H.-H.; Xu, P.-Y.; Chen, J.-X.; Yao, Q. Arbuscular mycorrhizal fungus enhances lateral root formation in Poncirus trifoliata (L.) as revealed by RNA-Seq analysis. Front. Plant Sci. 2017, 8, 2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javot, H.; Pumplin, N.; Harrison, M.-J. Phosphate in the arbuscular mycorrhizal symbiosis: Transport properties and regulatory roles. Plant Cell Environ. 2007, 30, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-Y.; Wu, Q.-S.; Zou, Y.-N. Effects of arbuscular mycorrhizal fungi on phosphorous uptake and phosphatase release in trifoliate orange seedlings. Mycosystema 2017, 36, 942–949. [Google Scholar] [CrossRef]
- Pumplin, N.; Harrison, M.-J. Live-cell imaging reveals peri arbuscular membrane domains and organelle location in Medicago truncatula roots during arbuscular mycorrhizal symbiosis. Plant Physiol. 2009, 151, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-C.; Gui, S.-H.; Yang, T.; Walk, T.; Wang, X.-R.; Liao, H. Identification of soybean purple acid phosphatase genes and their expression responses to phosphorus availability and symbiosis. Ann. Bot. 2012, 109, 275–285. [Google Scholar] [CrossRef]
- Shu, B.; Wang, P.; Xia, R.-X. Effects of mycorrhizal fungi on phytate phosphorus utilization in trifoliate orange (Poncirus trifoliata L. Raf) seedlings. Acta Physiol. Plant 2014, 36, 1023–1032. [Google Scholar] [CrossRef]
- Veneklaas, E.-J.; Lambers, H.; Bragg, J.; Finnegan, P.-M.; Lovelock, C.-E.; Plaxton, W.-C.; Price, C.-A.; Scheible, W.-R.; Shane, M.-W.; White, P.-J.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef]
- Wasaki, J.; Yamamura, T.; Shinano, T.; Osaki, M. Secreted acid phosphatase is expressed in cluster roots of lupin in response to phosphorus deficiency. Plant Soil 2003, 248, 129–136. [Google Scholar] [CrossRef]
- Wu, H.-H.; Zou, Y.-N.; Rahman, M.-M.; Ni, Q.-D.; Wu, Q.-S. Mycorrhizas alter sucrose and proline metabolism in trifoliate orange exposed to drought stress. Sci. Rep. 2017, 7, 42389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; Li, Y.; Wu, Q.-S. Exogenous carbon magnifies mycorrhizal effects on growth behaviour and sucrose metabolism in trifoliate orange. Not. Bot. Horti Agrobot. 2018, 46, 365. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.-Q.; Yamagishi, M.; Osaki, M.; Masuda, K. Sugar signaling mediates cluster root formation and phosphorus starvation-induced gene expression in white lupin. J. Exp. Bot. 2008, 59, 2749–2756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, S.-K.; Arthikala, M.-K.; Nanjareddy, K.; Lara, M. Plant-symbiont interactions: The functional role of expansins. Symbiosis 2017, 74, 1–10. [Google Scholar] [CrossRef]
- Mackowska, K.; Jarosz, A.; Grzebelus, E. Plant regeneration from leaf-derived protoplasts within the Daucus genus: Effect of different conditions in alginate embedding and phytosulfokine application. Plant Cell Tiss. Org. 2014, 117, 241–252. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Sequence of Forward Primer (5′→3′) | Sequence of Reverse Primer (5′→3′) |
|---|---|---|---|
| Cl18SrRNA | FJ356261.1 | TCGGGTGTTTTCACGTCTCA | CGAAGGGTCGCCGTAGGT |
| ClRSI-1 | Cs6g17190 | GCTCCGGTCATCAGGTAATT | CCGAACACCGATAAGAACA |
| ClPSK6 | Cs1g21390.1 | GTGCTTTTCTCCTCCTCATC | CAGGCTCATAGCTGAAACATG |
| ClKRP6 | Cs1g14980.2 | AGGATCAGACCTCAATGGACAA | CAAACGGAGGACTTACTTCTCA |
| ClPAP1 | JQ666163 | GATTTAGTCGTGGCTGGTCAT | GATAGACTGGAGCAGAAGGGT |
| ClPAP2 | JQ666164 | GTGGTGGCTGAGGGAAGA | TCGGCAAAGATGAAGTCG |
| ClPAP3 | JQ666165 | TACAACCTCAACAGTCAGTCACA | CCTTCCCCAATAATCCCAAC |
| ClPT1 | JQ666156 | GCTGCTCTTACTTACTACTGGC | TGCTACCTTGTCCTCCTGA |
| ClPT2 | JQ666157 | TAATGGTGTAGCGTTCTGTGG | ATAGTGGCGGAAAGTGGGTAG |
| ClPT3 | JQ666158 | ACTCTGTTTCTTTCGCTTCTG | TCTTCTTGTTGGCATACTCG |
| ClPT5 | JQ666160 | GGGGTTCCTCTGCTCTTT | ATGCTTTCCGTTGGTTGC |
| ClPT7 | JQ666162 | AACCACAACTCGGGACAA | GAAAGCAGCAGGAATAGA |
| Treatment | Root AMF Colonization (%) | Mycorrhizal Dependence (%) | Plant Height (cm) | Leaf Number | Biomass (g FW/Plant) | ||
|---|---|---|---|---|---|---|---|
| Leaf | Stem | Root | |||||
| P0 − Ri | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 8.18 ± 0.84 b | 8 ± 1 b | 0.27 ± 0.02 c | 0.26 ± 0.02 b | 0.53 ± 0.04 c |
| P0 + Ri | 38.96 ± 0.35 a | 68.63 ± 5.38 a | 14.1 ± 1.39 a | 16 ± 2 a | 1.73 ± 0.16 a | 0.82 ± 0.07 a | 1.16 ± 0.16 a |
| P50 − Ri | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 8.48 ± 0.58 b | 7 ± 1 b | 0.27 ± 0.02 c | 0.26 ± 0.03 b | 0.75 ± 0.06 b |
| P50 + Ri | 24.21 ± 0.24 b | 49.10 ± 5.01 b | 14.22 ± 0.59 a | 15 ± 1 a | 0.79 ± 0.04 b | 0.79 ± 0.08 a | 1.09 ± 0.11 a |
| Significance | |||||||
| Ri | 0.0082 | 0.0079 | <0.0001 | <0.0001 | <0.0001 | 0.4637 | 0.0411 |
| P | <0.0001 | <0.0001 | 0.5136 | 0.0407 | <0.0001 | <0.0001 | 0.0001 |
| Ri × P | 0.0082 | 0.0079 | 0.0358 | 0.0307 | <0.0001 | 0.5669 | 0.0438 |
| Treatment | Total Length (cm) | Project Area (cm2) | Surface Area (cm2) | Average Diameter (mm) | Volume (cm3) | Taproot Length (cm) | Number of Lateral Roots (Number/plant) | ||
|---|---|---|---|---|---|---|---|---|---|
| 1st-Order | 2nd-Order | 3rd-Order | |||||||
| P0 − Ri | 141.0 5 ± 9.90 b | 10.67 ± 0.56 b | 13.29 ± 1.01 bc | 0.662 ± 0.017 c | 0.548 ± 0.045 c | 13.50 ± 0.62 c | 39 ± 3 b | 17 ± 1 c | 3 ± 1 b |
| P0 + Ri | 202.18 ± 6.29 a | 13.31 ± 1.46 a | 15.20 ± 0.82 a | 0.853 ± 0.054 b | 0.862 ± 0.068 a | 21.33 ± 1.59 a | 60 ± 4 a | 59 ± 2 a | 14 ± 1 a |
| P50 − Ri | 118.68 ± 6.14 c | 9.49 ± 0.96 c | 12.36 ± 0.71 c | 0.655 ± 0.026 c | 0.381 ± 0.024 d | 12.27 ± 1.25 c | 40 ± 3 b | 22 ± 2 c | 0 ± 0 c |
| P50 + Ri | 147.73 ± 10.58 b | 11.09 ± 1.03 b | 13.35 ± 0.59 b | 0.978 ± 0.075 a | 0.799 ± 0.014 b | 17.17 ± 0.76 b | 41 ± 3 b | 45 ± 3 b | 4 ± 1 b |
| Significance | |||||||||
| Ri | <0.0001 | 0.0009 | 0.0177 | <0.0001 | <0.0001 | <0.0001 | 0.0004 | <0.0001 | <0.0001 |
| P | <0.0001 | 0.0034 | 0.0308 | 0.0679 | 0.0017 | 0.0032 | 0.0013 | 0.0043 | <0.0001 |
| Ri × P | 0.0112 | 0.2499 | 0.1977 | 0.0476 | 0.0713 | 0.0058 | 0.0007 | <0.0001 | 0.0016 |
| Variation Source | Ri | P | Ri × P |
|---|---|---|---|
| P content | <0.0001 | 0.0333 | <0.0001 |
| Phosphatase activity | 0.3334 | 0.2716 | 0.7298 |
| ClPAP1 | 0.006 | 0.0367 | 0.0507 |
| ClPAP2 | 0.4564 | 0.0103 | 0.3730 |
| ClPAP3 | 0.0150 | 0.1147 | 0.2643 |
| ClPT1 | 0.0001 | 0.0219 | 0.9495 |
| ClPT2 | 0.7931 | 0.0003 | 0.0003 |
| ClPT3 | 0.0011 | 0.1411 | 0. 0373 |
| ClPT5 | <0.0001 | 0.2870 | 0.6459 |
| ClPT7 | 0.0004 | 0.2712 | 0.2846 |
| ClPSK6 | 0.7902 | 0.0317 | 0.1747 |
| ClKRP6 | 0.7981 | 0.0329 | 0.2313 |
| ClRSI-1 | 0.0704 | 0.1922 | 0.0037 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-Y.; Guo, X.-N.; Wu, X.-L.; Dai, F.-J.; Wu, Q.-S. The Comprehensive Effects of Rhizophagus intraradices and P on Root System Architecture and P Transportation in Citrus limon L. Agriculture 2022, 12, 317. https://doi.org/10.3390/agriculture12030317
Liu C-Y, Guo X-N, Wu X-L, Dai F-J, Wu Q-S. The Comprehensive Effects of Rhizophagus intraradices and P on Root System Architecture and P Transportation in Citrus limon L. Agriculture. 2022; 12(3):317. https://doi.org/10.3390/agriculture12030317
Chicago/Turabian StyleLiu, Chun-Yan, Xiao-Niu Guo, Xiao-Long Wu, Feng-Jun Dai, and Qiang-Sheng Wu. 2022. "The Comprehensive Effects of Rhizophagus intraradices and P on Root System Architecture and P Transportation in Citrus limon L." Agriculture 12, no. 3: 317. https://doi.org/10.3390/agriculture12030317