1. Introduction
Time of bud break and flowering in deciduous fruit trees is controlled by the number of chill days accumulated over winter, with the warming weather of spring triggering the transition from endormancy to ecodormany, and ultimately bud growth [
1]. With a warming climate there is a growing concern that current growing regions may become marginal for fruit production [
2,
3], and in many growing regions it has become routine to apply chemicals to synchronise floral bud break, ensuring a commercially viable flowering and fruit set. These treatments can be particularly important if winter chilling and/or spring conditions have been suboptimal [
4]. The use of dormancy breakers is becoming increasingly common in sweet cherry (
Prunus avium) production. With continually increasing high premium prices compared with other horticultural products, sweet cherry is rapidly becoming one of the highest value tree crops in Australia.
According to Petri et al. [
5] and Rademacher [
6], hydrogen cyanamide (HC) is the leading plant bioregulator used to substitute for chilling. However, the toxicity of HC [
7] and its ban in Europe [
8] have led to the exploration of other less toxic chemicals as alternatives [
4,
5,
9], many of which are based on thiourea, nitrate salts and mineral oils [
6]. Emulsified vegetable oil compounds (EVOC) have also been shown to be an effective trigger of floral bud break in apples (
Malus domestica) [
9] and sweet cherry (
Prunus avium) [
10]. In addition to synchronising bud development, the application time of dormancy breakers can impact flowering time; Bound and Jones [
11] observed advanced flower development following early application of HC to apples, while late applications delayed flower development. A similar effect was reported by Bound and Miller [
10] following the application of the EVOC Waiken
®.
There is strong evidence that plant hormones play key roles in maintaining floral bud dormancy and inducing bud break in deciduous fruit trees, including prominent roles for auxin, abscisic acid, cytokinins and gibberellins and some evidence of roles for ethylene [
12].
Some of the evidence of the involvement of these hormones in floral bud dormancy and/or break is based on observation of the expression of hormone related genes [
13,
14,
15,
16]. However, as many hormone pathways are under strong feedback control, these gene expressions may not necessarily reflect hormone level. More compelling evidence comes from studies that quantify endogenous hormone levels in buds over the course of floral bud break [
17,
18]. Application studies, including the application of hormones or hormone precursors and substances that modify hormone level or action are also informative.
In cherries, several hormone groups have been considered as potentially important regulators of floral bud break from studies using both field-collected and detached branches. Hormone quantification in buds transitioning from dormant to growing has indicated abscisic acid may maintain endodormancy [
18], while HC-induced bud burst was associated with elevated cytokinin in floral buds [
17] and gibberellin levels in whole branches [
19]. However, there are some inconsistencies. For example, a reduction in ABA content prior to bud break has been found in some but not all studies of cherries, and in apples, HC-induced bud burst has been associated with elevated auxin [
20], but this was not observed in HC-treated cherries [
17]. The role of gibberellin as a promoter of bud burst is also unclear, as the application of gibberellin has been found to both suppress or promote bud break depending on the species and time of application [
8,
21,
22,
23].
Some studies have included an examination of the effect of dormancy-breaking agents on fruit set, yield and maturity in a range of crops [
5,
24,
25,
26,
27,
28], but most studies are restricted to the impacts of dormancy-breaking agents on bud break and flowering. Hence there is limited information available on fruit growth and quality at harvest on crops treated with dormancy-breaking agents.
This study examined the effect of two chemically distinct dormancy-breaking agents (HC and EVOC) in sweet cherry to determine (1) their influence on bud break and the endogenous hormone content of buds to ascertain whether they act via modification of the same hormone triggers, (2) the effect of treatments on bud development through to shuck fall and (3) effects on fruit growth and quality at harvest.
2. Materials and Methods
A field trial was established on mature ‘Lapins’ sweet cherry trees grown on F12/1 rootstock in a commercial orchard in the Derwent Valley, Tasmania (42.7548° S, 146.9853° E) in late winter 2018. Trial design was a randomised complete block design with three treatments (HC, EVOC, and untreated control), each with four replicates and a plot size of three trees. Three representative branches were tagged on each tree for bud-stage counts during the season and for obtaining fruit samples for quality assessments at harvest.
The two dormancy-breaking chemicals applied in this study were HC in the form of Dormex
® (520 g/L cyanamide, NuFarm Australia Limited, Melbourne, VIC, Australia) and EVOC applied as Waiken
® (388 g/L methyl esters of fatty acids, SST Australia Pty Ltd., Dandenong South, VIC, Australia). Treatments were applied in late winter with the application rate and time based on label recommendations and aimed to avoid either delay or advancement of flowering. Hydrogen cyanamide was applied at 2.5%
v/v and EVOC at 4%
v/v; application times are shown in
Table 1. All sprays were applied to the whole tree to runoff at a water volume of 1000 L/ha. At the time of spray application, buds on all trees were dormant or at the early bud swell stage.
2.1. Bud Sampling for Phenological Assessment and Hormone Analysis
Flower buds were collected twice weekly over the course of six weeks, commencing on 2 August 2018. Buds were selected at random from the centre tree in each replicate, avoiding the tagged branches. Six to eight buds were collected for determination of bud stage. For hormone analysis a similar number of buds were harvested and placed directly into pre-weighed tubes containing 80% methanol with 250 mg L−1 butylated hydroxytoluene. All samples were placed in a portable cooler and returned to the laboratory for bud stage assessment and hormone extraction.
2.1.1. Bud Stage Assessment
Buds were inspected under a Leica M80 stereo microscope fitted with a Leica DFC 295 camera and Leica software application suite version 3.8.0 and rated for bud phenological stage based on the BBCH growth stage scale for stone fruit [
33,
34] (
Figure 1).
2.1.2. Hormone Extraction
At each harvest date, tubes were weighed and the initial weight was subtracted to determine tissue weight. Buds were then processed to extract endogenous hormones from the excised flower buds using the following procedure. On average, six buds (~300 mg fresh weight) were included per sample. Hormones were extracted according to Großkinsky et al. [
35] with minor adjustments. Samples were pulverised with a physcotron homogeniser (Microtech), standards added and incubated overnight at 4 °C. Stable isotope-labelled internal standards were [
13C
6]-indole-3-acetic acid, [
2H
6] abscisic acid (Cambridge Isotope Laboratories, Tewksbury, MA, USA) and cytokinins: [
2H
5] zeatin (Z), [
2H
5] zeatin riboside (ZR), [
2H
3] di hydrozeatin (DiHZ), [
2H
3] di-hydrozeatin riboside (DiHZR), [
2H
6] isopentenyl adenine (Isop adenine) and [
2H
6] isopentenyl adenosine (Isop adenosine) (OlChemIm, Olomouc, Czech Republic). Samples were spun and supernatants loaded onto preconditioned C18 500 mg SepPak (Waters, Rydalmere, Australia) with methanol. Eluates were collected, taken to dryness, resuspended in 20% methanol, sonicated for eight minutes and transferred to auto-sampling vials for analysis by ultra-performance liquid chromatography tandem mass spectrometry using a Waters Acquity H-Class UPLC instrument coupled to a Waters Xevo triple quadrupole mass spectrometer.
A Waters Acquity C18 BEH column (100 mm × 2.1 mm × 1.7 um) coupled in series to a Waters PFP BEH column (100 mm × 2.1 mm × 1.7 um) was used for analyte analysis. The UPLC programme used to separate the metabolites of interest consisted of Mobile Phase A (89.5:10:0.5,
v/
v/
v) water:methanol:acetic acid and Mobile Phase B (99.5:0.5,
v/
v) methanol:acetic acid. A gradient elution was used starting at 10% B and moving to 55% B at 12.0 min, with a 1 min hold, followed by an increase to 95% B at 13.5 min with a 4 min hold. Re-equilibration to starting conditions occurred at 17.6 min for 3 min. The flow rate was 0.20 mL min
−1 with the column held at 35 °C, and the sample compartment was at 6 °C. The mass spectrometer was operated in positive and negative ion electrospray mode with a needle voltage of 2.8 kV, and MRM was used to detect all analytes (
Appendix A,
Table A1). The ion source temperature was 130 °C, the desolvation gas was N
2 at 950 L per hour
−1, the cone gas flow was 100 L per hour
−1, and the desolvation temperature was 450 °C. Data were processed using MassLynx software. The endogenous hormone level contained within each sample was calculated by comparing the endogenous hormone transition peak area and corresponding internal standard peak area.
2.2. Field Assessment of Bud Development and Phenological Stage
The total number of floral buds was recorded on each tagged branch on 16 August 2018, and each bud was rated for phenological stage (
Figure 1). Phenological stage counts were then completed at regular intervals until 18 October 2019, when flowering was complete and all set fruit had reached shuck fall stage. For each assessment date the percentage of buds at each phenological stage was calculated.
2.3. Fruit Growth and Quality Sampling and Assessment
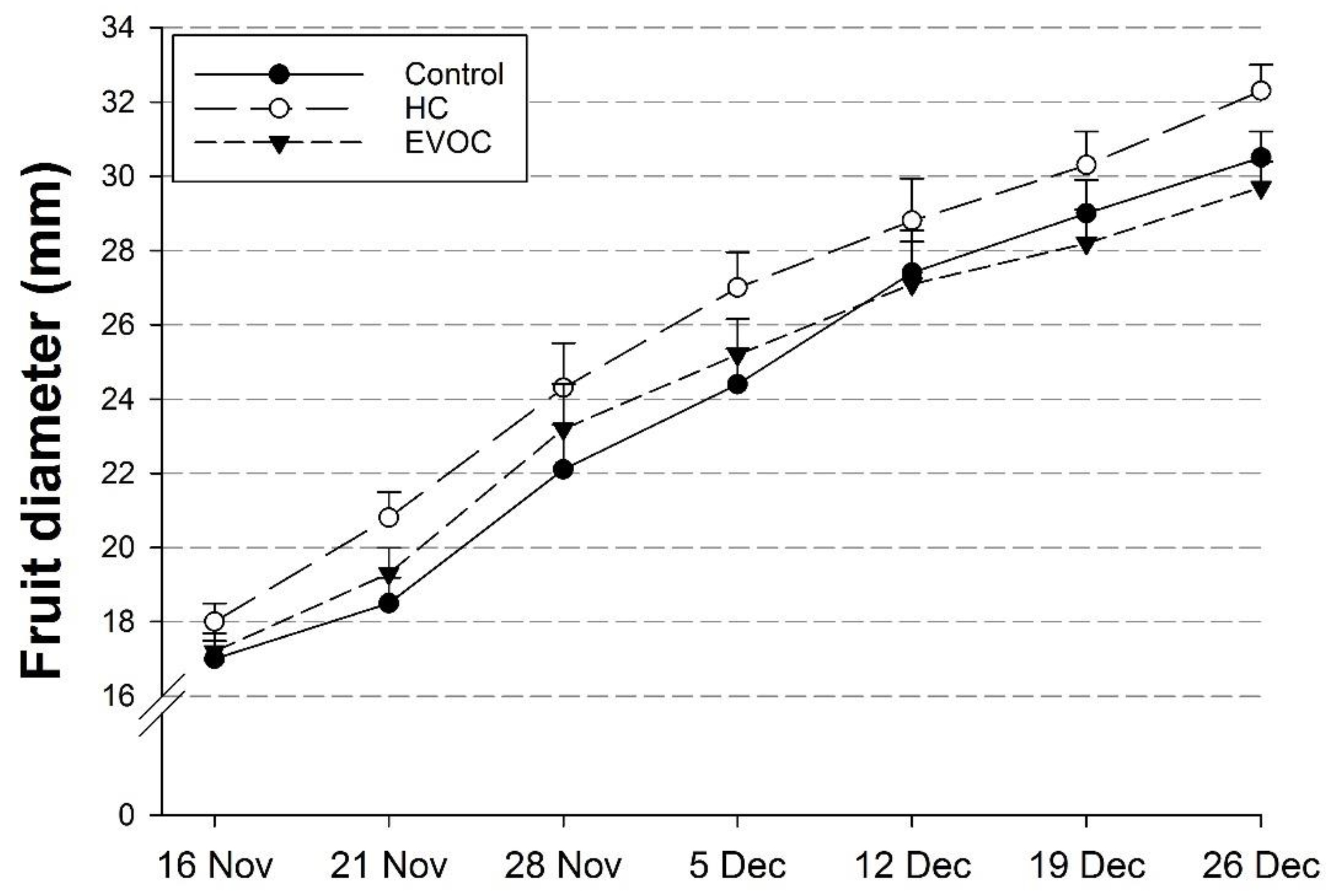
Fruit samples were collected at weekly intervals from 16 November 2018 when control fruit reached straw stage through to 27 December 2018 (commercial harvest). At each sampling date, random samples of four fruits were taken from each of the three trees in each plot to give 12 fruits per replicate, avoiding the tagged branches. At commercial harvest on 27 December, all of the fruit from each tagged branch was harvested. Harvested fruits were weighed, counted and sorted into A grade, B grade and reject. For quality assessment, two sets of samples of 10 fruits were randomly selected from the A grade fruit for each branch to give 30 fruits per tree and a total of 90 fruits per replicate based on three trees per plot. One sample set was assessed immediately, and the second sample was stored at 0 °C for 6 weeks for post-harvest quality assessment. Quality assessments included fruit weight, diameter, skin colour, compression firmness, flesh firmness, skin puncture force, flesh colour, stem pull force, dry matter content (DMC), total soluble solids (TSS) content, malic acid (MA) concentration, and juice pH.
Fruit weight was measured on digital scales (Mettler-Toledo Model: TLE3002) and diameter and length was measured with digital calipers (DigiMax, Wiha-41101, Buchs, Switzerland). Skin colour was assessed with both the Australian Cherry Colour Guide (Cherry Growers Australia Inc., Hobart, Australia) and a Konica Minolta Colourimeter—Chroma Meter CR-400. Colour space parameters obtained from the colourimeter included the L*a*b* colour space. Fruit compression firmness was determined with a FirmTech 2 (Bioworks Inc., Wamego, KS, USA) and flesh firmness and skin puncture force were both measured with a fruit texture analyser (Güss model GS-20, Strand, South Africa). Stem pull force was measured using a stand mounted Mark-10 Series 5 force gauge (Mark-10, Copiague, NY, USA). Seeds were then removed from all of the fruit, and fifteen fruits from each replicate were placed into pre-weighed bags and oven dried to determine DMC; the remaining fifteen fruits were juiced collectively, and duplicate samples were taken for measurement of TSS, pH, and MA concentration. TSS content, expressed as °Brix, was measured with an Atago PR-1 digital refractometer (Atago Co., Ltd., Tokyo, Japan). Juice pH was the initial pH value of 10 mL juice samples measured using a Mettler Toledo G20 compact titrator (Mettler Toledo, Melbourne, Australia). MA concentration was calculated as g L−1.
Australian sweet cherry industry standards classifying fruit into domestic and export finest were used as a reference for fruit quality (
Table 2).
Climatic data for the bud development period was obtained from the nearest Bureau of Meteorology station at Bushy Park, 4.5 km from the trial site (
Figure 2).
2.4. Statistical Analysis
For the bud data, one-way analysis of variance (ANOVA) was carried out to compare treatments at each time point using the RStudio statistical software version 3.6.0 (RStudio, Boston, MA, USA). Statistical significance was assumed at p ≤ 0.05. Fruit growth and quality data were subjected to ANOVA using Genstat release 17.1 (VSN International Ltd., Hemel Hempstead, Hertfordshire, UK). Data are presented as mean values for each treatment. Significance was calculated at p = 0.05 and least significant difference (LSD) was used for the comparison of mean values in the tables and figures.
4. Discussion
4.1. Bud Break and Hormone Analysis
In this study, cytokinin was identified as a potentially important marker of bud break in sweet cherry, with a clear elevation of cytokinin prior to bud break across treatments. Specifically, the results indicate that dihydrozeatin and dihydrozeatin riboside are key regulators of bud break as they were significantly elevated by HC treatments prior to bud burst compared to control plants, suggesting that these hormones were clear predictors of the transition from dormancy to growth in floral buds. Dihydrozeatin and dihydrozeatin riboside are major forms of active cytokinin found in many plant tissues [
38], and this key role is consistent with the findings of Ionescu et al. [
17] who reported that HC induced a more rapid increase in the level of these compounds prior to and during bud release. This is also consistent with studies in grape and mango that found bud break was associated with elevated cytokinin content in buds, although it should be noted these studies used suboptimal methods for hormone quantification such as ELISA and antibodies [
39,
40].
The decrease in abscisic acid observed in this study prior to bud burst across all treatments was consistent with previous observations in untreated cherry trees across multiple seasons [
18]. However, more rapid reduction to abscisic acid level of buds did not appear to underpin the hastened bud break observed in HC-treated plants. This is consistent with the findings of Wang et al. [
19] and Ionescu et al. [
17] and suggests that although reduction in abscisic acid may be important for release from endodormancy it is not the mechanism through which HC induced bud break.
Recent studies have revealed central roles for gibberellin in HC-induced bud break of cherry, apple and grape [
19,
20,
41]. Although this method was not optimised for gibberellin, future studies will examine the role of gibberellin in bud break induced by HC and other less toxic dormancy breakers including EVOC.
Although EVOC treatment led to a small but significant hastening of bud break compared to control plants, this was not reflected in any significant change in hormone profile. While the recommended application windows for the two chemicals differ, applying EVOC one week later than HC may have contributed to the relatively mild effect of EVOC compared to previous trials and such a mild effect may be difficult to distinguish at the level of hormone action. Future trials with more significant effects of EVOC on bud break will seek to explore the hormonal underpinning of less toxic regulators and examine whether it also influences cytokinin content.
4.2. Phenological Development and Fruit Growth
The application timing of treatments was aimed at concentrating flowering rather than advancing or delaying bud break, but the earlier bud break and flowering observed with HC but not EVOC in this study demonstrates that, in practice, this is difficult to achieve. Godini et al. [
27] state that climatic conditions can have a critical effect on the efficient use of HC. In a study of EVOC on apples, Bound and Miller [
9] observed that higher concentrations held trees in dormancy, resulting in delayed bud break, and Brunt et al. [
42] confirmed that this dormancy breaker induces a period of dormancy following application. Although EVOC was applied later than HC in this study, the responses observed by Bound and Miller [
9] and Brunt et al. [
42] may partially explain the delay in bud break in the EVOC-treated trees compared to HC. Sheard et al. [
43] concluded that application of dormancy-breaking agents should be based not only on calendar date but on a combination of chill unit accumulation and visual bud development stage. It has been reported that the efficacy of the EVOC Waiken
® was improved by 20–30% when combined with potassium nitrate [
26].
Winter chill accumulation influences bloom time [
27], and the chilling requirement is cultivar specific [
44]. Chill requirements for the cultivar ‘Sweetheart’ have been calculated as 1066 chill hours (CH) (54 chill portions [CP]) and for ‘Kordia 1307 CH (67 CP) [
44]. The chill rating for ‘Lapins’, used in this study, is described by Brunt et al. [
42] as low–moderate to moderate–high with the chill requirement ranging from 45–62 CP. Chemical dormancy-breaking agents are more effective when 50% or more of the chilling requirement of the plant has been satisfied [
5]. In this study, 100% of the chill requirement had already been met prior to the application of both dormancy-breaking agents (75.8 CP for HC and 82.1 for EVOC), and this may explain the similar development observed between the untreated control and the EVOC treatments.
Reporting on a three-year study on the impact of HC on bud break and fruit ripening in several sweet cherry cultivars, Godini et al. [
27] suggested that the effects of HC do not extend beyond the induction of advanced bud break, and that climatic factors influence fruit growth from bloom onward, generally decreasing any initial gains. However, in our study fruit growth in HC-treated trees remained more advanced throughout the growing season.
4.3. Harvest Packout and Fruit Quality
Despite an advancement in bud break following application of HC, no advancement in maturity was observed in treated trees in this study. This conflicts with the findings of Raffoa et al. [
28] who reported an advancement in ripening of 7–9 days in ‘Burlat’ and ‘Ferrovia’ in Argentina.
There is limited information available on the impact of dormancy-breaking agents on fruit quality, but in this study different effects on fruit size and pack-out were observed between the two chemicals used. The 60% increase in cracking incidence in HC-treated trees is most likely linked to the larger fruit size in these trees, as larger fruits have been shown to be more susceptible to cracking [
45]. This high incidence of cracking also contributed to the reduction in pack-out of A-grade fruit as cracked fruit are downgraded or rejected depending on the severity of the cracking.
The increased fruit size observed in HC-treated trees was most likely due to the earlier budburst and flowering in these trees which resulted in a longer growing period. Petri [
46] suggested that an advancement in flowering should lead to an increase in fruit weight. Ardiles and Ayala [
7] suggested that greater photo assimilate availability as a result of higher leaf area early in the season optimises cell division during fruit development, thus contributing to increased fruit size. Cherry fruits continue to increase in size and weight the longer that they are left on the tree (Bound and Hölzel, unpublished data).
Stem retention is an important quality attribute for consumers, with a minimum stem pull force of 500 g being the Australian benchmark for both domestic and export fruit. Minimum fruit compression firmness standards are also listed in the industry standards, with a value of 250 g mm−2 and 300 g mm−2 for domestic and export standards, respectively. Although fruit compression firmness and stem pull force at harvest were reduced slightly by EVOC, these two quality parameters were still above the threshold required to meet export quality standard.
In addition to size, firmness and stem retention, the other important quality parameter is TSS. There are no reports of the impact of EVOC on TSS in sweet cherries, but this study has shown that EVOC has no effect on TSS in ‘Lapins’. In relation to the impact of HC on TSS, the results of both Raffoa et al. [
28] and Ardiles and Ayala [
7] are consistent with the findings of this study that HC has no effect.
Although there were some small effects, particularly of EVOC, on some fruit quality parameters, this study has demonstrated that the impact was not sufficient to downgrade fruit from export finest to domestic. However, the increase in fruit cracking and reduction in pack-out of A grade fruit in HC-treated fruit will impact on grower returns, so caution should be exercised in application of this dormancy-breaking agent.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







