1. Introduction
Soil salinization is one of the most critical abiotic stressors that impacts crop yields globally; salinity threatens roughly 6% of the world’s total land area, including 20% of arable land and 33% of irrigated land [
1]. Land salinization is increasing, with 10 million acres of agricultural land damaged each year by salt buildup caused by human activities and other climate change-related variables [
2]. Plant growth and productivity are dramatically reduced by salinity stress, which can drastically affect production [
3]. Faba bean is one of the oldest crops in the world and its cultivation dates back to the Mediterranean region [
4]. Globally, 4.7 million tons of grain legume crop were produced over 3.4 million hectares [
5]. Despite its age and commercial importance, the fact is that faba bean is a diploid species (2n = 12), and modest progress has been made in developing an excellent genetic understanding of this crop. Faba bean has an exceptionally large genome of approximately 13.4 Gb [
6], and it considered to be the largest genome in the grain legume family.
Vicia faba L. is a cool-season legume crop and produces high-protein grains for human production and livestock in developing regions [
7,
8]. The Vicia genus belongs to the
Viceae tribe, a cool-season clade of the subfamily. Papilionoideae is part of the legume family Fabaceae [
9]. Because of the increasing demand for faba bean consumption in the middle east, there is a need to develop faba bean genotypes that are suitable for arid and semiarid regions [
8].
Salinity tolerance is a complicated physiological feature with several sub-components; the traditional view holds that salinity impacts plant performance through osmotic stress and specific ion toxicity [
10]. Salt concentration in soil severely affects faba bean yield [
11]; however, faba bean seed germination is more sensitive to growth under salinity. Characterizing salt tolerance, faba bean genotypes have shown enhanced productivity under salinity [
12]. Plants compensate for decreasing osmotic potential in the rhizosphere by minimizing water loss. It has been claimed that stomatal transpiration accounts for around 95% of plant water loss [
13]. Lower stomatal density is an essential physiological characteristic in salinity-tolerant quinoa [
14,
15], while farmed barley uses a stress-escaping strategy by lowering stomatal density to conserve water when grown in saline circumstances [
16]. Saline-induced stomatal closure would reduce CO
2 inflow, reducing leaf photosynthetic capacity and, eventually, yield [
17]. Ionic toxicity, which is induced by high sodium accumulations in the cytoplasm, is fundamental for plants under salt stress [
18]. Na
+ exclusion from the shoot is thought to be crucial for plants to overcome the adverse effects of increasing salt, and a large percentage of Na
+ exclusion (>98%) in wheat is achieved by limiting net Na
+ absorption at the soil–root interface and net xylem loading in roots [
19]. The K
+/Na
+ ratio is thought to be the fundamental trait imparting salinity stress resistance in plants, and it is frequently used as a screening tool for plant breeders [
20]. Under salt stress, Na
+ and K
+ transporters are critical for maintaining Na
+ and K
+ homeostasis in cells and plants [
21]. Enhancing the salt-tolerance capacity is one of the most efficient and viable approaches for reducing the negative impact that salinity has on crop output [
22].
Many screening techniques for salt tolerance have been conducted. The replicability of the experiments and the consistent result amongst laboratories remains challenging because of a lack of a standard growing environment [
23], and a few of these experiments were part of large-scale studies [
24,
25]. Even though salinity tolerance is a polygenic trait, several studies have treated it as a single trait and have evaluated it using visual scoring [
26]. The pyramiding of favorable morphological, physiological, and biochemical factors can enhance salt tolerance [
27]. A statistical model that incorporates morphological, physiological, and biochemical factors would be more appropriate [
28]. As a result, multivariate analysis helps to find the genetic origins of variation and to differentiate salt tolerance using several selection criteria. Methods for determining the salt tolerance of many genotypes must be economical, rapid, and easily quantifiable to achieve this goal [
29]. Because of the variability in environmental variables from season to season, morphological measures sometimes need a large amount of phenotypic data and repetitive cropping seasons for a screening assessment. Variability in agricultural soil can also harm field evaluations, increasing the coefficient of variation, leading to breeders being led away from attaining their objectives [
30]. Under salt stress, performing the morphological, physiological, and biochemical criteria would naturally differ among faba bean genotypes, with one genotype being superior in at least one feature while being poorer in others. Our research hypothesis was that the different faba bean genotypes would respond differently under different salinity conditions, modifying the cultivar tolerance ranking seen at different salt stress levels. This work aimed to investigate these discrepancies to see if these measures might be used as reliable screening criteria for genotype assessment in salinity circumstances with advanced statistical techniques.
3. Results
The descriptive statistics of the studied characters are presented in
Table 2. The table includes minimum, maximum, mean, and standard deviation (SD) values. All parameters recorded a high range of difference among the studied characteristics, demonstrating that salinity affects the agronomic, physiological, and biochemical characteristics in faba bean genotypes (
Table 2). The mean average trait values of all the faba bean genotypes are presented in the
Table S1 in the Supplementary Materials.
The analysis of variance data is presented in
Table 3. All of the studied characteristics recorded significant differences among the control and salinity treatments.
The principal components analysis of all of the studied traits based on the combined (control, S1 = 100 mM and S2 = 200 mM) treatments were analyzed (
Table 4). The first three PCs represent 81.45% of the total variance among the studied traits.
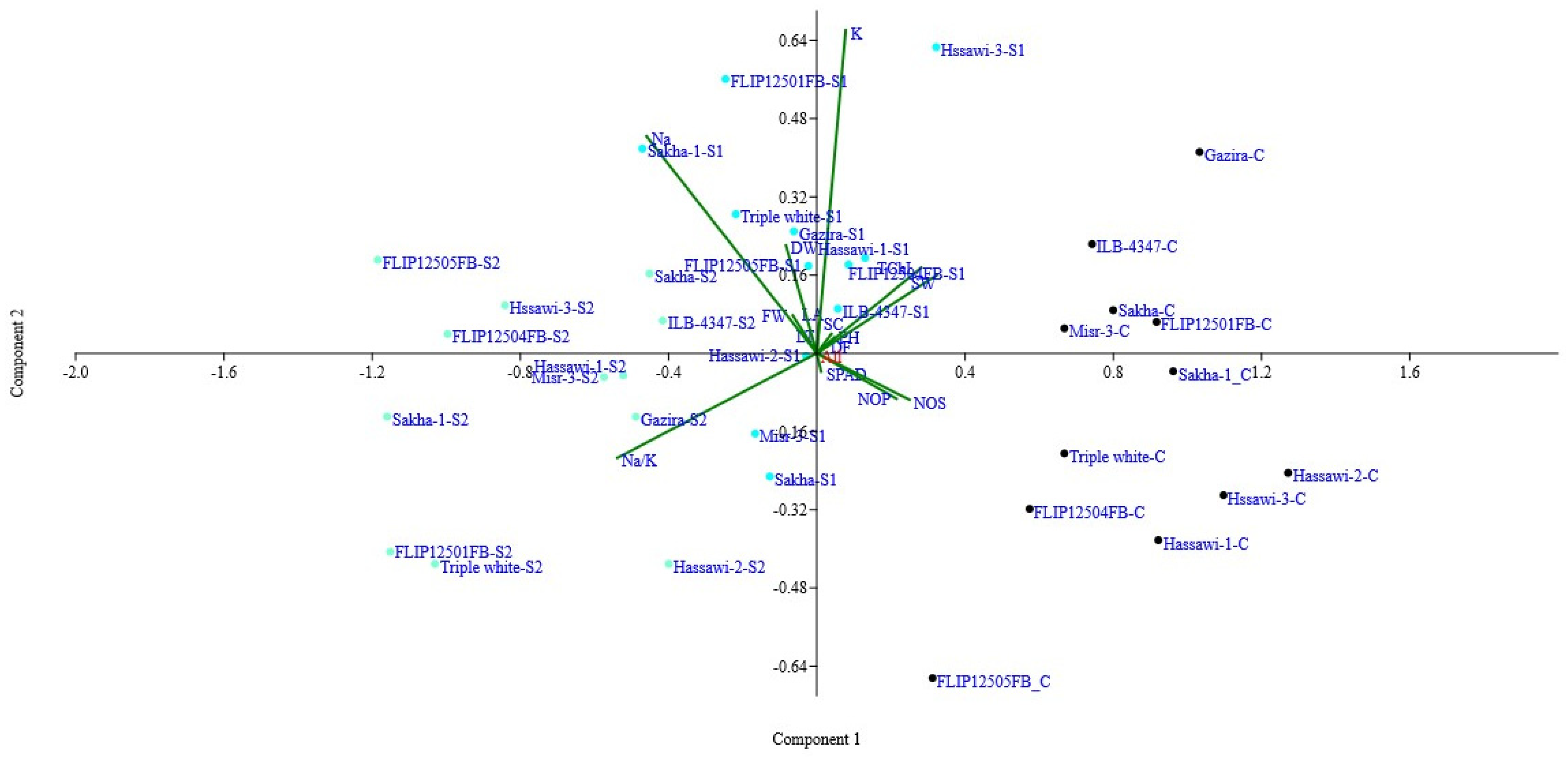
The biplot analysis (PC1 and PC2) was presented in
Figure 1. The results suggested that the genotypes (Hassawi-2, Sakha, ILB-4347, Misr-3, FLIP12501FB) were present in the positive region of PC1 (first quadrant) under control conditions. Similarly, Hassawi-1, Hassawi-2, Sakha, Gazira, and FLIP12504FB were present in the same group. The most notable characteristics that contributed to the variance under control and salinity conditions were the K
+, seed weight (g), and days to 50% flowering. The genotypes (Sakha-1, triple white, Hassawi-3, Gazira, Hassawi-3) were present in PC1 (second quadrant). The significant characters contributed more in PC1 (2nd quadrate), SPAD, seed number, and pods/plant (
Figure 1). However, at higher salinities (S2 = 200 mM), the more significant genotypes are (Hassawi-2, Hassawi-1, ILB-4347) in one group; however, FLIP12501FB, Sakha-1, and triple white are away from the center (sensitive) but are present in PC2 (second quadrant) (
Figure 1). The most important characteristics contributed more in terms of PC2 leaf fresh weight (g), leaf dry weight (g), leaf area (cm
2), Na
+, K
+, and Na
+/K
+).
We generated the regression analysis to determine the prediction of the variables studied (
Table 5). The table describes the slope, error, intercept, correlation coefficient, and probability values. The regression results suggested that the characteristics, i.e., the number of pods/plant, number of seeds/pod, seed weight, and Na
+/K
+ ratio, were more affected by an increase in salinity. The leaf area (cm
2) remained unaffected as the salinity increased (
Table 5).
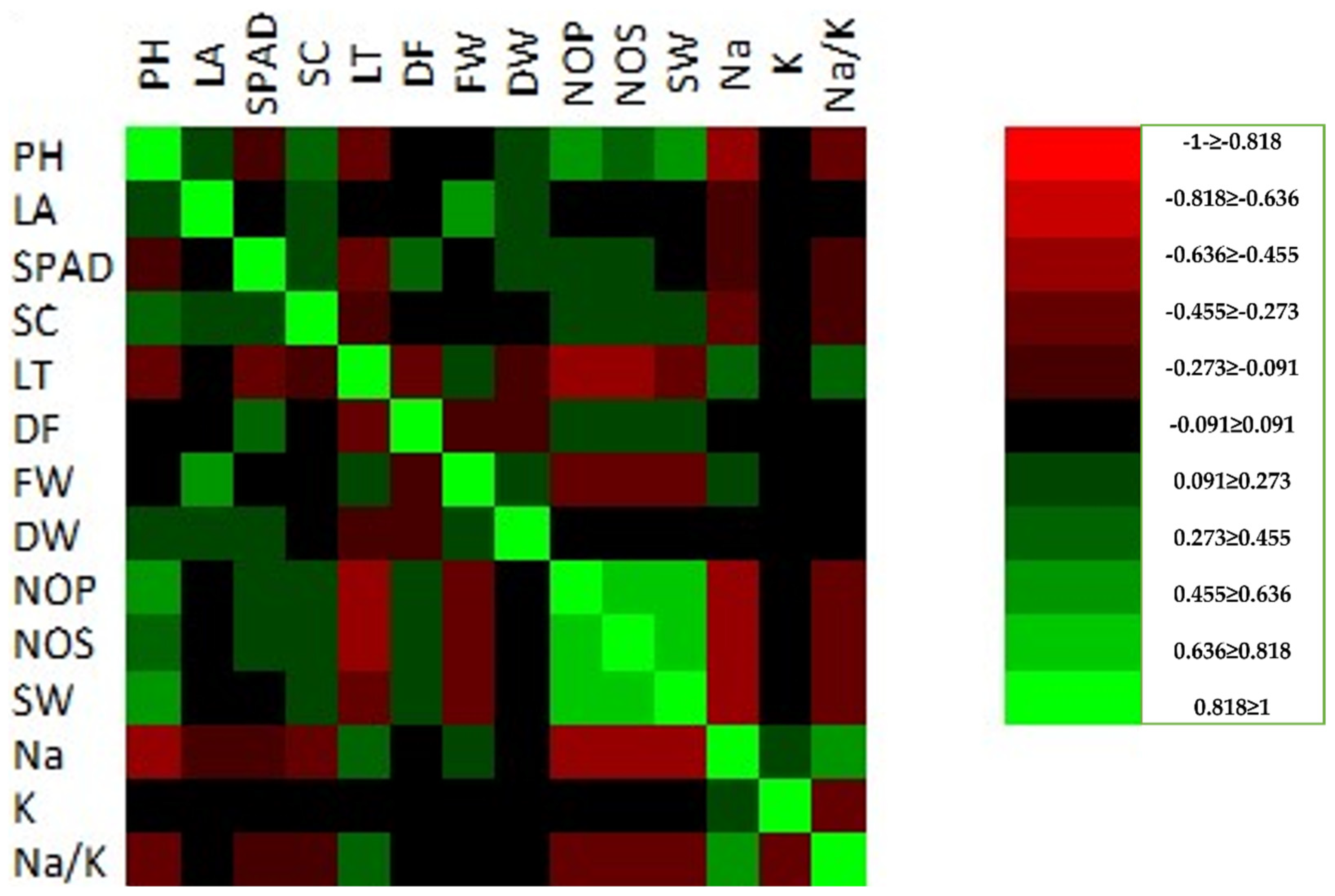
The Pearson’s correlation matrix was also determined for the studied characteristics using the combined mean data of the control, S1 = 100 mM, and S2 = 200 mM (
Figure 2).
A significant correlation was recorded for plant height (cm) with leaf area (cm
2), stomatal conductance mmol (m
−2 s
−1), dry weight (g), number of pods, number of seeds, and seeds weight (g), while a negative correlation was recorded for SPAD, leaf temperature, Na
+, K
+, and Na
+/K
+. The fresh weight (FW) and dry weight (DW) were the characteristics that were the most significantly affected by the different salinity conditions. The FW was strong negatively correlated to the PH, SPAD, SC, DF, NOP, NOS, SW, K, and Na
+/K
+. The absorbance of the Na
+, K
+, and Na
+/K
+ indicates the salinity tolerance under the field conditions. These three parameters (Na
+, K
+, and Na
+/K
+) were recorded as being insignificant for all of the studied parameters; however, a significant correlation was recorded for LT and FW (
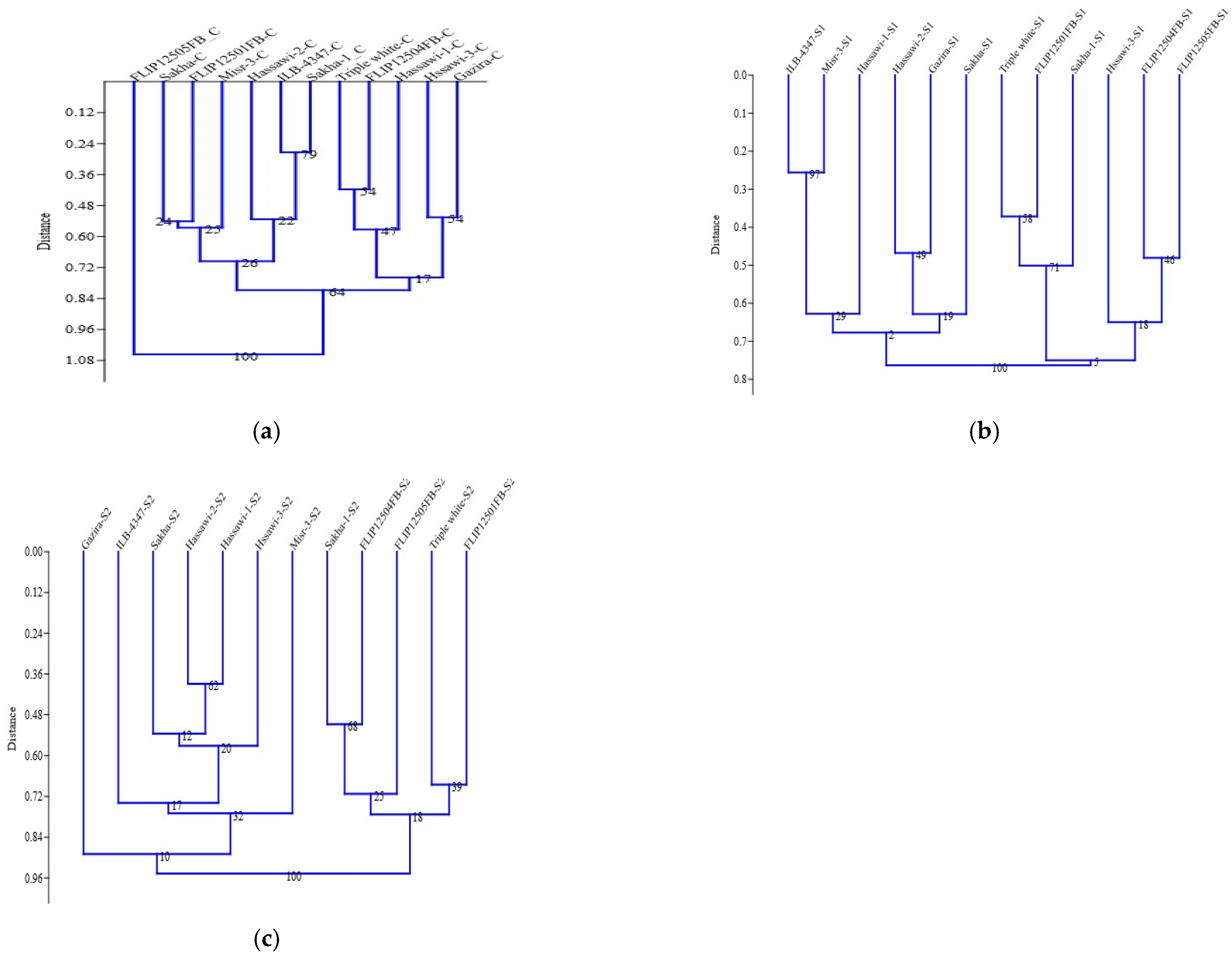
Figure 2). The cluster analysis was used to group the faba bean genotypes based on the control, 100 mM, and 200 mM salt stress conditions (
Figure 3a–c). The cluster analysis for control the treatments was divided into two groups. The first group was further subdivided into two subgroups. The first group compassed the 11 genotypes, while the FLIP12505FB genotype was individually separated (
Figure 3c). Under the 100 mM condition, the genotypes were equally distributed between the two main groups. Each main group was subdivided into two subgroups under the 100 mM salt stress condition. Sakha, Misr-3, Hassawi-1, Gazira, Hassawi-2, and ILB-4347 were allocated to group I (
Figure 3b); however, triple white was grouped with Sakha-1, Hassawi-3, and three genotypes of FLIP1250FB. Similarly, under the 200 mM salt stress condition, the genotypes were clustered into two main groups, and the Gazira genotype failed to be grouped and was separated into a group of its own. Group 1 comprised five genotypes (FLIP12505FB, triple white, FLIP12504FB, FLIP12501FB, and Sakha-1), while group II contained the Hassawi-1, Hassawi-2, Hassawi-3, Sakha, and Misr-3 genotypes (
Figure 3c).
Some salt tolerance indexes (YI, SSI, STI, TOL, MPI, MRP, REI) were also calculated based on seed weight/plant to determine the susceptibility and tolerance level of each faba bean genotype (
Table 6).
The maximum YI was reported in Gazira, followed by Hassawi-2 under the 100 mM salinity condition. The minimum YI was recorded in the FLIP12505FB genotype. Similarly, the maximum YI was observed in Sakha followed by in ILB-4347 (1.25), Hassawi-3 (1.23), and Hassawi-2 (1.16 under the 200 mM salt stress condition. The minimum YI (0.54) was recorded (FLIP12504FB) genotype under a 200 mM salt stress level. The maximum SSI (0.800) was recorded in Hassawi-1 (100 mM), ILB-4347 (0.646; 200 mM), while the minimum SSI (0.016) was recorded for Hassawi-2 under the 100 mM salt stress condition. The maximum (1.678) salinity tolerance index was recorded in Hassawi-2, and the minimum was recorded in the FLIP12504FB faba bean genotype under the 100 mM salinity conditions, while for the 200 mM salinity conditions, the maximum was recorded for ILB-4347 and Hassawi-2, respectively. The mean productivity index was recorded maximum for Hassawi-2 under both the 100 mM and 200 mM salt stress conditions, while the minimum was recorded for the FLIP12505FB genotype under both stress conditions. The maximum relative efficiency index was recorded in Hassawi-2 followed by in ILB-4347 under both of the salt stress (100 mM and 200 mM) conditions. The correlation matrix was also determined among the salt stress indices under moderate and high salt (100 mM and 200 mM) stress conditions (
Table 7). The YI recorded a negative correlation with SSI at both stress levels; however, all of the other index (SSI, STI, TOL, MPI, REI) values recorded positive and significant correlations.
4. Discussion
The present study was conducted to determine the tolerance of different faba bean genotypes using different agro-morphological, physiological, and biochemical characteristics under different salinity levels. The descriptive and analysis of variance values recorded significant differences among the studied characteristics. Similar results [
42] suggested that significant differences were recorded in the studied characteristics under different abiotic stress conditions in barley varieties. Among different types of legumes, Faba bean is salt-sensitive a [
43]. In the faba bean genotypes studied here, the salt treatments lowered biomass production and water intake [
44]. Ahmad et al. [
43] also found that NaCl inhibited faba bean growth and biomass yield. Cell division inhibition and cell elongation are the result of the loss in growth and biomass yield caused by salinity [
45]. NaCl reduces growth and biomass yield by reducing mineral intake, generating reactive oxygen species, inhibiting enzyme activity, and causing hormonal imbalances [
46]. Ahmad et al. [
43] suggested that sensitive cultivars are more vulnerable to damage relative to tolerance in pea plants and mulberry seedlings [
43]. Similar results were reported in
Solanum lycopersicum and
Vicia faba [
42]. Sodium and potassium ions share similar physiochemical structures and compete for Na and K ion uptake in the soil [
47]. Other plant species, such as mustard [
48] and strawberry, have increased Na
+ buildup with lower K
+ and Ca
+ absorption when exposed to NaCl [
49]. The inhibition of these mineral elements, the primary cause of stunted plant growth and development, is caused by high saline levels [
47].
The analysis of variance results recorded high levels of significant differences among the studied traits under control and salinity. Similar results were reported by Filipović et al. [
50] and suggested that increasing the salinity (100 mM) of the irrigation water significantly reduced morphological characteristics. The biplot loadings for PC1 and PC2 were performed related to salinity in order to group the faba bean genotypes. Similar results were recorded when phenotypical data were analyzed, and the genotypes were grouped in the hierarchical cluster pattern [
51]. Multivariate analytical techniques such as PCA and cluster analysis were used [
52] and was significant in determining the best genotypes under stress conditions. The findings refer to Karadavut [
53] and Mekonnen et al. [
54]. Similarly, the cluster analysis results reported that the faba bean genotypes were divided into two groups under the control and 100 mM salinity levels; however, the 200 mM salinity l level recorded three groups of faba bean genotypes, showing that salinity stress may limit phenotypic variability among faba bean genotypes. The genotypic variation was decreased, and the genotype distribution in the group was also different because of the increased salt stress level. Similar results were in line with those recorded by Saed-Moucheshi et al. [
55], who suggested that stress reduced the phenotypic variation among the studied variables and limited the diversity potential in crop plants, such as in triticale [
56,
57].
Some essential multivariate strategies have been adequately employed to determine tolerant and susceptible genotypes in response to varied environmental situations. In-plant breeding and screening programs, clustering analysis, regression approaches, and principal component analysis are popular methodologies [
58]. There are a variety of univariate strategies for detecting tolerant genotypes that work in different ways. Other studies have identified a variety of univariate methodologies that are suitable for determining the optimum genotype for various environmental situations. The susceptibility and tolerant indices are powerful tools that help to screen genotypes under abiotic stress conditions [
55]; similarly, STI is also a vital tolerance index to determine the relationship among the other stress indices [
59]. A positive correlation was found with TOL under control conditions in our study. However, a negative correlation was reported under stress conditions. Similar results were reported by Talebi et al. [
60]. Similar results were also reported in another study, suggesting that there is positive association between TOL under control conditions and a negative association under stress conditions suggests that TOL selection would reduce yield under control conditions [
61,
62]. As a result, integrating multivariate approaches with tolerance indices is an efficient way to use all of the computed indices in genotype screening systems. Besides our study, Fernandez [
63] used the multivariate analysis technique in different crop plants such as mung bean and wheat [
60,
64,
65], and used a similar type of analysis [
62] in lentil to introduce tolerant and susceptible genotypes. This tolerance could be because of genetic variances in salinity tolerance rather than differences in ROS detoxification capabilities. Differences in antioxidant enzyme expression or activity are linked to more tolerant genotypes, but they can also be linked to more sensitive genotypes. The differences among agro-morphological, physiological, and biochemical characteristics among faba bean genotypes could be because of genotypic differences in stomatal closure or in other responses that affect the rate of CO
2 fixation [
10].
5. Conclusions
The study confirms the significance of the studied traits, salinity index, and their association to determine the salinity tolerance of each genotype. The presentation of the data using multivariate analyses (descriptive, ANOVA, PCA, cluster analysis, regression analysis, correlation analysis, and salinity indices) also forces the establishment and definition of tolerance levels. Four genotypes, Hassawi-2, ILB-4347, Sakha, Misr-3, and Flip12501FB, showed significant tolerance levels; however, FLIP12504FB represents a sensitive genotype based on its final grain yield. Significant index correlations with grain yield supports the use of these indices as tools for screening faba bean genotypes for salinity stress evaluation. In short, these findings show that faba bean genotypes can perform in a semiarid environment and can be grown under salt stress conditions where high temperatures and salinity are the constraints. The salinity tolerance trait is complicated and can be easily improved through breeding selection. However, the best results could be achieved through molecular genetics and genomics techniques by unraveling the molecular mechanism involved when crops are under salinity stress conditions. Furthermore, characterizing new salt tolerance genes could help researches to improve the faba bean genome under abiotic stress conditions in the future, especially under high salinity and high temperature conditions. Consider these tolerant genotypes and utilize them for breeding and genetic programs to improve salinity sensitivity in faba bean.


 ,
, 



{kind=link}
{kind=link}
{kind=link}