
Assessing the Effect of a Major Quantitative Locus for Phosphorus Uptake (Pup1) in Rice (O. sativa L.) Grown under a Temperate Region
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Pup1 Introgression into Temperate Rice through Marker-Assisted Backcrossing
2.3. Foreground and Background Genotypic Selection
2.4. Field Trials and Experimental Design
2.5. Phenotyping and Identification of Low P and Water Tolerance Indices
2.6. Statistical Analysis of Phenotypic Data
3. Results
3.1. Introgression of the Full Pup1 Locus and RP Genome Recovery Rates in Temperate Rice
3.2. Vegetative Stage Response of Temperate Pup1 Introgression Lines to Low Inputs of P and Water
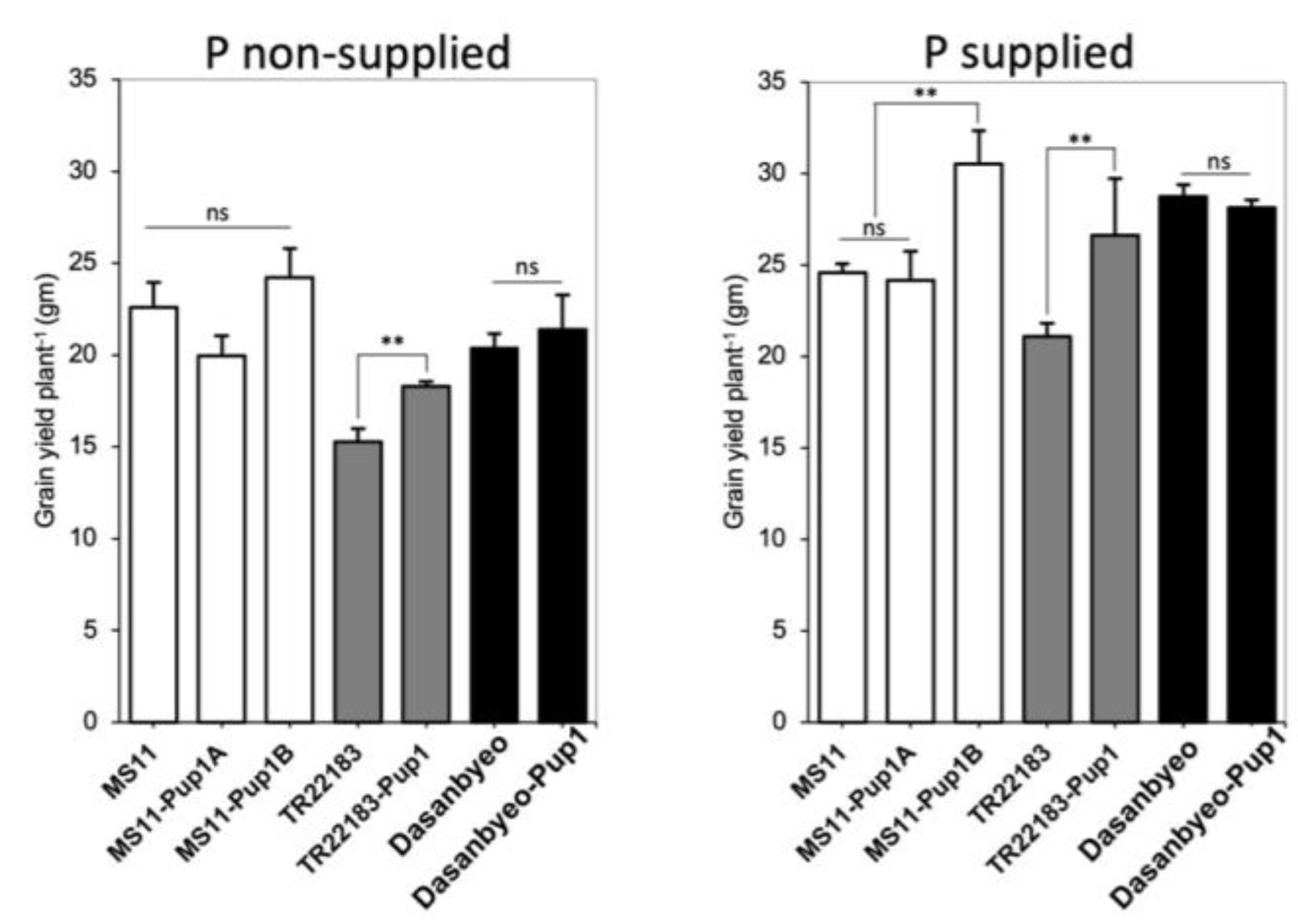
3.3. Yield Response to Low P and Rainfed Condition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [Green Version]
- Andrade, J.F.; Cassman, K.G.; Edreira, J.I.R.; Agus, F.; Bala, A.; Deng, N.; Grassini, P. Impact of urbanization trends on production of key staple crops. Ambio 2022, 51, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Haefele, S.M.; Hijmans, R.J. Soil quality in rice-based rainfed lowlands of Asia: Characterization and distribution. In Proceedings of the 26th International Rice Research Conference, Delhi, India, 9–12 October 2006; pp. 297–308. [Google Scholar]
- Fess, T.F.; Kotcon, J.B.; Benito, V.A. Crop breeding for low-input agriculture: A sustainable response to feed a growing world population. Sustainability 2011, 3, 1742–1772. [Google Scholar] [CrossRef] [Green Version]
- Beddington, J. Contributions from science to a new and greener revolution. Philos. Trans. R. Soc. B Biol. Sci. 2010, 265, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batjes, N.H. A world data set of derived soil properties by FAO- UNESCO soil unit for global modeling. Soil Use Manag. 1997, 13, 9–16. [Google Scholar] [CrossRef]
- Dobermann, A.; Fairhurst, T. Phosphorus deficiency. In Rice: Nutrient Disorders and Nutrient Management; International Rice Research Institute: Los Banos, Philippines, 2000; pp. 60–71. [Google Scholar]
- Kirk, G.J.D.; George, T.; Courtois, B.; Senadhira, D. Opportunities to improve phosphorus efficiency and soil fertility in rainfed lowland and upland rice ecosystems. Field Crops Res. 1998, 56, 73–92. [Google Scholar] [CrossRef]
- Van Kauwenbergh, S.; Steward, M.; Mikkelsen, R. World reserves of phosphate rock: A dynamic and unfolding story. Better Crops 2013, 97, 18–20. [Google Scholar]
- Carpenter, S.R. Phosphorus control in critical to mitigating eutrophication. Proc. Natl. Acad. Sci. USA 2008, 105, 11039–11040. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.M.; Heuer, S.; Thomson, J.T.; Wissuwa, M. Genetic and genomic approaches to develop rice germplasm for problem soils. Plant Mol. Biol. 2007, 65, 547–570. [Google Scholar] [CrossRef]
- Wissuwa, M.; Ae, N. Genotypic variation for tolerance to phosphorus deficiency in rice and the potential for its exploitation in rice improvement. Plant Breed 2001, 120, 43–48. [Google Scholar] [CrossRef]
- Shin, N.-H.; Lee, O.N.; Han, J.-H.; Song, K.; Koh, H.-J.; Yoo, S.-C.; Chin, J.H. The Effect of Water Level in Rice Cropping System on Phosphorus Uptake Activity of Pup1 in a Pup1+Sub1 Breeding Line. Plants 2021, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Ha, W.G.; Torollo, G.V.; Kang, K.H.; Lapiz, M.; Padolina, T.F. Breeding temperate japonica rice MS11 for the tropics. Philipp. J. Crop Sci. 2011, 36, 111. [Google Scholar]
- Kim, B.; Kim, D.G.; Lee, G.; Seo, J.; Choi, I.Y.; Choi, B.S.; Yang, T.J.; Kim, K.S.; Lee, J.; Chin, J.H.; et al. Defining the genome structure of “Tongil” rice, an important cultivar in the Korean “Green Revolution”. Rice 2014, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, J.H.; Kim, J.H.; Jiang, W.Z.; Chu, S.H.; Woo, M.O.; Han, L.Z.; Brar, D.S.; Koh, H.J. Identificaiton of subspecies-specific STS markers and their association with segregation dis- tortion in rice (Oryza sativa L.). J. Crop Sci. Biotechnol. 2007, 10, 175–184. [Google Scholar]
- Cho, Y.I.; Jiang, W.Z.; Chin, J.H.; Piao, Z.Z.; Cho, Y.I.; McCouch, S.R.M.; Koh, H.J. Identification of QTLs associated with physiological nitrogen use efficiency in rice. Mol. Cells 2007, 23, 72–79. [Google Scholar]
- Jiang, W.; Lee, J.; Jin, Y.M.; Qiao, Y.; Piao, R.; Jang, S.M.; Woo, M.O.; Kwon, S.W.; Liu, X.; Pan, H.Y.; et al. Identification of QTLs for seed germination capability after various storage periods using two RIL populations in rice. Mol. Cells 2011, 31, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Chin, J.H.; Lu, X.; Haefele, S.M.; Gamuyao, R.; Ismail, A.; Wissuwa, M.; Heuer, S. Development and application of gene-based markers for the major rice QTL Phosphorus uptake 1. Theor. Appl. Genet. 2010, 120, 1073–1086. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Thomson, M.J.; Singh, N.; Dwiyanti, M.S. Large-scale deployment of a rice 6 K SNP array for genetics and breeding applications. Rice 2017, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.W.; Nam, J.; Chu, S.H. Development of KNU Axiom Oryza 580K Genotyping Array. In Proceedings of the KSBS & SABRAO International Conference on Plant Breeding for Sustainable Development, Gwangju, Republic of Korea, 2–5 July 2019; PCS02-58. p. 58. [Google Scholar]
- Chin, J.H.; Gamuyao, R.; Dalid, C.; Bustamam, J.; Prasetiyono, J.; Moeljopawiro, S.; Wissuwa, M.; Heuer, S. Developing Rice with High Yield under Phosphorus Deficiency: Pup1 Sequence to Application. Plant Physiol. 2011, 156, 1202–1216. [Google Scholar] [CrossRef] [Green Version]
- Gamuyao, R.; Chin, J.H.; Tanaka, J.P.; Pesaresi, P.; Catausan, S.; Dalid, C.; Heuer, S. The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature 2012, 488, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Parfitt, R.L. Phosphate reactions with natural allophone, ferrihydrite and goethite. Soil Sci. 1989, 40, 359–369. [Google Scholar] [CrossRef]
- Redel, Y.; Cartes, P.; Demanet, R.; Velasquez, G.; Poblete-Grant, P.R.; Bol, R.; Mora, M.L. Assessment of phosphorus status influenced by Al and Fe compounds in volcanic grassland soils. J. Soil Sci. Plant Nutr. 2016, 16, 490–506. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.S.; Yang, X.B.; Wang, C.L.; Hao, X.J.; Hong, J.P.; Lin, X.Y. Dynamics of available and enzymatically hydrolysable soil phosphorus fractions during repeated freeze-thaw cycles. Geoderma 2019, 345, 1–4. [Google Scholar] [CrossRef]
- Wissuwa, M.; Yano, M.; Ae, N. Mapping of QTLs for phosphorus deficiency tolerance in rice (Oryza sativa L.). Theor. Appl. Genet. 1998, 97, 777–783. [Google Scholar] [CrossRef]
- Navea, I.P.; Dwiyanti, M.S.; Park, J.; Kim, B.; Lee, S.; Huang, X.; Koh, H.; Chin, J.H. Identification of quantitative trait loci for panicle length and yield related traits under different water and P application conditions in tropical region in rice. Euphytica 2017, 213, 37. [Google Scholar] [CrossRef]
- Seo, J.; Lee, S.-M.; Han, J.-H.; Shin, N.-H.; Lee, Y.K.; Kim, B.; Chin, J.H.; Koh, H.-J. Characterization of the Common Japonica-Originated Genomic Regions in the High-Yielding Varieties Developed from Inter-Subspecific Crosses in Temperate Rice (Oryza sativa L.). Genes 2020, 11, 562. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Pedigree | Generation | Genomic Similarity to RP |
|---|---|---|---|
| MS11 | RP | RP | N/A |
| MS11-Pup1A | (MS11*3/IR64-Pup1)-8-24-11-5-2-1-1-1-1-1-4 | BC2F12 | 71.68% |
| MS11-Pup1B | (MS11*3/IR64-Pup1)-17-15-8-3-5-4-1-1-1-1-4 | BC2F12 | 66.50% |
| TR22183 | RP | RP | N/A |
| TR22183-Pup1 | (TR22183*3/IR64-Pup1)-1-2-24-2-4-1-1-1-1-1-4 | BC2F12 | 99.75% |
| Dasanbyeo | RP | RP | N/A |
| Dasanbyeo-Pup1 | (Dasanbyeo*3/IR64-Pup1)-2-6-5-3-3-3-1-1-1-1-4 | BC2F12 | 92.33% |
| Line | Treatment | Plant Height (cm) | Tiller Number | SPAD Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 30 DAT | 44 DAT | 54 DAT | 30 DAT | 44 DAT | 54 DAT | 30 DAT | 44 DAT | 54 DAT | ||
| MS11 | P non-supplied | 39.53 a ± 0.21 | 55.50 a ± 1.08 | 53.50 b ± 1.14 | 3.50 ± 0.43 | 4.00 b ± 0.76 | 5.67 ab ± 1.15 | 38.45 b ± 0.66 | 35.18 b ± 0.93 | 30.95 b ± 1.36 |
| MS11-Pup1A | 38.00 a ± 0.58 | 50.67 b ± 2.52 | 61.67 a ± 1.53 | 3.33 ± 1.00 | 5.00 ab ± 0.58 | 5.00 b ± 0.67 | 36.27 b ± 1.95 | 31.67 b ± 2.99 | 34.19 ab ± 2.17 | |
| MS11-Pup1B | 34.83 b ± 1.67 | 44.67 c ± 2.03 | 50.33 b ± 1.67 | 3.00 ± 0.88 | 5.33 a ± 1.53 | 6.33 a ± 0.88 | 46.30 a ± 1.21 | 39.50 a ± 2.70 | 36.77 a ± 1.27 | |
| TR22183 | 38.00 a ± 1.00 | 58.00 ± 5.13 | 72.33 ± 1.86 | 1.67 b ± 0.33 | 2.33 ± 0.33 | 3.00 ± 0.58 | 44.87 ± 0.81 | 50.97 a ± 0.98 | 41.53 a ± 0.78 | |
| TR22183-Pup1 | 34.00 b ± 1.00 | 55.00 ± 1.15 | 69.33 ± 1.20 | 2.00 a ± 0.00 | 2.67 ± 0.33 | 3.00 ± 0.00 | 44.77 ± 1.92 | 42.80 b ± 2.25 | 37.77 b ± 1.72 | |
| Dasanbyeo | 31.67 ± 0.71 | 43.50 a ± 2.26 | 51.00 a ± 2.45 | 3.17 ± 0.17 | 4.67 ± 0.33 | 5.50 ± 0.43 | 44.20 ± 1.22 | 42.18 b ± 1.06 | 39.17 ± 1.46 | |
| Dasanbyeo-Pup1 | 30.00 ± 1.15 | 38.67 b ± 0.88 | 46.33 b ± 0.67 | 2.33 ± 0.67 | 4.33 ± 0.67 | 5.00 ± 0.58 | 45.43 ± 2.13 | 44.50 a ± 1.10 | 37.57 ± 0.93 | |
| IR64 | 34.33 ± 1.34 | 41.33 a ± 0.34 | 42.33 a ± 1.15 | 3.50 ± 0.50 | 4.00 ± 0.26 | 6.83 ± 0.75 | 43.43 a ± 0.55 | 41.92 ± 0.81 | 38.35 ± 1.79 | |
| IR64-Pup1 | 32.17 ± 0.67 | 35.67 b ± 2.67 | 37.00 b ± 0.67 | 3.00 ± 0.00 | 3.67 ± 0.67 | 7.00 ± 0.58 | 39.90 b ± 0.61 | 42.27 ± 0.67 | 39.03 ± 3.52 | |
| MS11 | P supplied | 38.67 b ± 1.33 | 56.17 b ± 3.21 | 62.83 a ± 2.52 | 5.50 c ± 0.33 | 7.67 a ± 0.58 | 7.50 b ± 0.33 | 40.45 ± 1.46 | 33.57 b ± 1.43 | 30.57 b ± 2.20 |
| MS11-Pup1A | 40.00 a ± 1.33 | 63.00 a ± 4.26 | 66.00 a ± 2.03 | 7.00 a ± 0.00 | 10.00 b ± 0.67 | 10.67 a ± 0.58 | 42.17 ± 1.73 | 34.33 b ± 1.65 | 31.03 b ± 1.48 | |
| MS11-Pup1B | 35.67 b ± 3.06 | 51.33 b ± 0.33 | 56.67 b ± 1.67 | 6.33 b ± 0.33 | 11.00 b ± 1.15 | 11.33 a ± 0.58 | 43.70 ± 2.22 | 39.57 a ± 1.99 | 38.17 a ± 1.74 | |
| TR22183 | 38.33 ± 0.88 | 60.33 ± 0.67 | 71.67 ± 3.18 | 3.00 a ± 0.00 | 4.00 a ± 0.00 | 3.67 b ± 1.20 | 48.53 ± 1.41 | 46.67 a ± 8.16 | 45.20 a ± 1.42 | |
| TR22183-Pup1 | 39.67 ± 1.20 | 61.67 ± 2.03 | 70.67 ± 1.45 | 4.33 b ± 0.33 | 5.33 b ± 0.33 | 5.67 a ± 1.20 | 49.57 ± 2.08 | 42.20 b ± 0.68 | 38.73 b ± 3.69 | |
| Dasanbyeo | 34.10 a ± 0.43 | 49.08 a ± 1.20 | 53.33 ± 0.95 | 4.17 b ± 0.17 | 8.17 ± 0.60 | 7.83 b ± 0.65 | 44.93 ± 1.13 | 40.88 b ± 1.33 | 34.83 ± 1.62 | |
| Dasanbyeo-Pup1 | 30.67 b ± 0.33 | 46.33 b ± 0.33 | 52.83 ± 4.11 | 5.00 a ± 0.00 | 8.33 ± 1.20 | 10.00 a ± 0.58 | 46.31 ± 0.93 | 44.39 a ± 0.83 | 37.50 ± 0.71 | |
| IR64 | 37.67 ± 0.52 | 47.33 b ± 0.98 | 47.33 ± 1.56 | 4.33 b ± 0.42 | 8.67 b ± 1.67 | 10.50 b ± 0.43 | 43.47 ± 1.50 | 43.85 ± 0.98 | 37.57 a ± 0.93 | |
| IR64-Pup1 | 36.33 ± 2.33 | 50.67 a ± 0.88 | 48.67 ± 1.76 | 5.00 a ± 0.58 | 12.00 a ± 0.00 | 12.67 a ± 0.67 | 41.97 ± 3.56 | 42.27 ± 0.73 | 34.77 b ± 0.19 | |
| Line | Rainfed | Irrigated | ||||||
|---|---|---|---|---|---|---|---|---|
| Plant Height (cm) | Tiller Number | Plant Height (cm) | Tiller Number | |||||
| 40 DAT | 60 DAT | 40 DAT | 60 DAT | 40 DAT | 60 DAT | 40 DAT | 60 DAT | |
| MS11 | 69.90 a ± 0.67 | 88.73 a ± 1.25 | 11.73 b ± 0.52 | 12.73 c ± 0.46 | 78.00 ± 1.22 | 99.10 a ± 0.62 | 14.80 b ± 0.58 | 12.80 ± 1.02 |
| MS11-Pup1A | 69.13 a ± 1.55 | 83.33 b ± 0.99 | 17.33 c ± 0.80 | 17.27 b ± 0.56 | 77.00 ± 0.45 | 100.00 a ± 1.38 | 19.60 a ± 1.75 | 13.40 ± 1.03 |
| MS11-Pup1B | 51.87 b ± 0.96 | 69.13 c ± 1.58 | 25.93 a ± 0.63 | 25.27 a ± 0.86 | 76.00 ± 0.71 | 88.20 b ± 2.85 | 18.80 a ± 0.86 | 14.00 ± 0.84 |
| TR22183 | 70.63 ± 1.95 | 86.67 ± 2.76 | 5.33 b ± 0.36 | 6.87 b ± 0.32 | 87.80 a ± 1.36 | 106.70 ± 3.28 | 9.20 ± 0.49 | 9.60 ± 0.40 |
| TR22183-Pup1 | 69.53 ± 1.22 | 84.00 ± 0.93 | 9.20 a ± 0.54 | 10.53 a ± 0.79 | 75.20 b ± 1.32 | 109.00 ± 1.01 | 9.40 ± 0.51 | 8.20 ± 0.58 |
| Line | Treatment | Days to Heading (DAT) | Culm Length (cm) | Productive Tiller Number | Panicle Length (mm) | Hundred Grain Weight (g) | Fertility (%) |
|---|---|---|---|---|---|---|---|
| MS11 |
P non-supplied | 78 | 70.80 b ± 2.27 | 7.00 a ± 0.63 | 223.00 b ± 7.00 | 2.86 ± 0.05 | 94.04 ± 2.18 |
| MS11-Pup1A | 76 | 77.50 a ± 0.92 | 4.60 b ± 0.24 | 251.12 a ± 5.10 | 2.42 ± 0.13 | 94.01 ± 2.21 | |
| MS11-Pup1B | 82 | 66.00 c ± 0.45 | 5.40 b ± 0.24 | 251.61 a ± 5.20 | 2.62 ± 0.02 | 94.60 ± 3.46 | |
| TR22183 | 51 | 51.50 b ± 0.39 | 3.20 ± 0.20 | 254.21 b ± 4.20 | 2.82 ± 0.11 | 92.42 b ± 4.59 | |
| TR22183-Pup1 | 58 | 56.00 a ± 0.84 | 3.40 ± 0.51 | 267.10 a ± 3.30 | 2.91 ± 0.04 | 100.00 a ± 0.00 | |
| Dasanbyeo | 84 | 56.50 ± 1.20 | 4.20 ± 0.20 | 260.02 ± 3.51 | 2.70 ± 0.09 | 90.33 ± 2.94 | |
| Dasanbyeo-Pup1 | 91 | 53.20 ± 0.20 | 3.80 ± 0.37 | 261.61 ± 6.72 | 2.79 ± 0.08 | 95.43 ± 2.16 | |
| MS11 |
P supplied | 68 | 68.40 ± 2.07 | 9.20 b ± 0.86 | 209.02 c ± 4.60 | 2.57 ± 0.12 | 85.61 ab ± 2.75 |
| MS11-Pup1A | 72 | 73.30 ± 1.84 | 7.20 a ± 0.20 | 234.05 b ± 9.01 | 2.49 ± 0.04 | 90.53 a ± 4.17 | |
| MS11-Pup1B | 78 | 70.40 ± 1.21 | 9.20 b ± 0.58 | 261.02 a ± 8.05 | 2.66 ± 0.06 | 80.91 b ± 4.60 | |
| TR22183 | 56 | 59.40 b ± 1.71 | 4.80 ± 0.37 | 243.00 b ± 13.32 | 2.89 ± 0.04 | 99.17 a ± 0.83 | |
| TR22183-Pup1 | 65 | 64.90 a ± 1.25 | 5.20 ± 0.66 | 266.00 a ± 8.31 | 2.81 ± 0.05 | 95.19 b ± 1.54 | |
| Dasanbyeo | 82 | 69.30 a ± 0.86 | 7.00 ± 0.71 | 251.02 ± 5.88 | 2.63 ± 0.07 | 96.86 ± 1.59 | |
| Dasanbyeo-Pup1 | 82 | 61.50 b ± 0.32 | 6.20 ± 0.20 | 245.10 ± 3.87 | 2.83 ± 0.05 | 94.01 ± 1.84 |
| Line | Treatment | Culm Length (cm) | Productive Tiller Number | Panicle Length (mm) | Hundred Grain Weight (g) | Fertility (%) | Grain Yield per Plant (g) |
|---|---|---|---|---|---|---|---|
| MS11 | Rainfed | 47.43 c ± 0.60 | 14.60 ± 0.51 | 183.20 b ± 4.72 | 1.70 ± 0.16 | 20.06 c ± 2.44 | 13.29 b ± 1.20 |
| MS11-Pup1A | 64.14 a ± 1.45 | 12.60 ± 0.75 | 208.20 a ± 4.44 | 1.75 ± 0.82 | 69.98 a ± 1.75 | 27.31 a ± 1.29 | |
| MS11-Pup1B | 54.70 b ± 1.20 | 15.20 ± 1.02 | 204.40 a ± 4.12 | 1.66 ± 0.77 | 58.72 b ± 3.64 | 26.56 a ± 2.20 | |
| TR22183 | 43.44 ± 4.93 | 18.10 ± 2.20 | 236.00 ± 9.92 | 2.46 ± 0.24 | 20.04 b ± 5.21 | 18.10 b ± 3.19 | |
| TR22183-Pup1 | 52.76 ± 1.49 | 14.50 ± 1.57 | 235.20 ± 13.03 | 2.64 ± 0.99 | 66.94 a ± 2.74 | 25.71 a ± 0.87 | |
| MS11 | Irrigated | 79.76 ± 2.36 | 15.80 b ± 1.57 | 266.40 a ± 7.22 | 2.20 b ± 1.19 | 87.56 a ± 2.46 | 28.42 b ± 1.65 |
| MS11-Pup1A | 79.76 ± 2.36 | 14.50 b ± 0.58 | 222.00 b ± 2.83 | 2.18 b ± 0.80 | 78.64 b ± 3.45 | 29.72 b ± 1.87 | |
| MS11-Pup1B | 77.56 ± 2.27 | 20.60 a ± 1.17 | 259.20 a ± 8.33 | 2.78 a ± 1.44 | 88.58 a ± 1.50 | 52.02 a ± 2.23 | |
| TR22183 | 68.52 b ± 1.87 | 24.40 a ± 0.68 | 265.60 ± 9.50 | 2.84 ± 0.86 | 92.74 ± 1.97 | 45.64 ± 2.08 | |
| TR22183-Pup1 | 76.24 a ± 1.72 | 21.20 b ± 0.58 | 281.20 ± 2.94 | 2.83 ± 0.51 | 92.62 ± 0.97 | 42.32 ± 2.90 |
| Line | Days to Heading (DAT) | Culm Length (cm) | Productive Tiller Number | Panicle Length (cm) | Hundred Grain Weight (g) | Fertility (%) | Grain Yield per Plant (g) |
|---|---|---|---|---|---|---|---|
| MS11 | −0.13 | −0.03 | 0.31 | −0.06 | −0.10 | −0.09 | 0.09 |
| MS11-Pup1A | −0.05 | −0.05 | 0.57 | −0.07 | 0.03 | −0.04 | 0.21 |
| MS11-Pup1B | −0.05 | 0.07 | 0.70 | 0.04 | 0.02 | −0.14 | 0.26 |
| TR22183 | 0.10 | 0.15 | 0.50 | 0.05 | 0.02 | 0.07 | 0.38 |
| TR22183-Pup1 | 0.12 | 0.16 | 0.53 | −0.09 | −0.03 | −0.05 | 0.46 |
| Dasanbyeo | −0.02 | 0.23 | 0.67 | −0.03 | −0.03 | 0.07 | 0.41 |
| Dasanbyeo-Pup1 | −0.10 | 0.16 | 0.63 | −0.06 | 0.01 | −0.01 | 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navea, I.P.; Han, J.-H.; Shin, N.-H.; Lee, O.N.; Kwon, S.-W.; Choi, I.-R.; Chin, J.H. Assessing the Effect of a Major Quantitative Locus for Phosphorus Uptake (Pup1) in Rice (O. sativa L.) Grown under a Temperate Region. Agriculture 2022, 12, 2056. https://doi.org/10.3390/agriculture12122056
Navea IP, Han J-H, Shin N-H, Lee ON, Kwon S-W, Choi I-R, Chin JH. Assessing the Effect of a Major Quantitative Locus for Phosphorus Uptake (Pup1) in Rice (O. sativa L.) Grown under a Temperate Region. Agriculture. 2022; 12(12):2056. https://doi.org/10.3390/agriculture12122056
Chicago/Turabian StyleNavea, Ian Paul, Jae-Hyuk Han, Na-Hyun Shin, O New Lee, Soon-Wook Kwon, Il-Ryong Choi, and Joong Hyoun Chin. 2022. "Assessing the Effect of a Major Quantitative Locus for Phosphorus Uptake (Pup1) in Rice (O. sativa L.) Grown under a Temperate Region" Agriculture 12, no. 12: 2056. https://doi.org/10.3390/agriculture12122056