
Morphological, Cytological, and Molecular-Based Genetic Stability Analysis of In Vitro-Propagated Plants from Newly Induced Aneuploids in Caladium

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. In Vitro Culture of the Aneuploid Variants
2.3. Plant Acclimatization and Establishment
2.4. Visual Observation and Morphological Characterization
2.5. Stomatal Assessment
2.6. Flow Cytometry Analysis
2.7. Chromosome Counting
2.8. SSR Analysis of Nuclear Genome
2.9. Data Analysis
3. Results
3.1. Plant Regeneration and Establishment
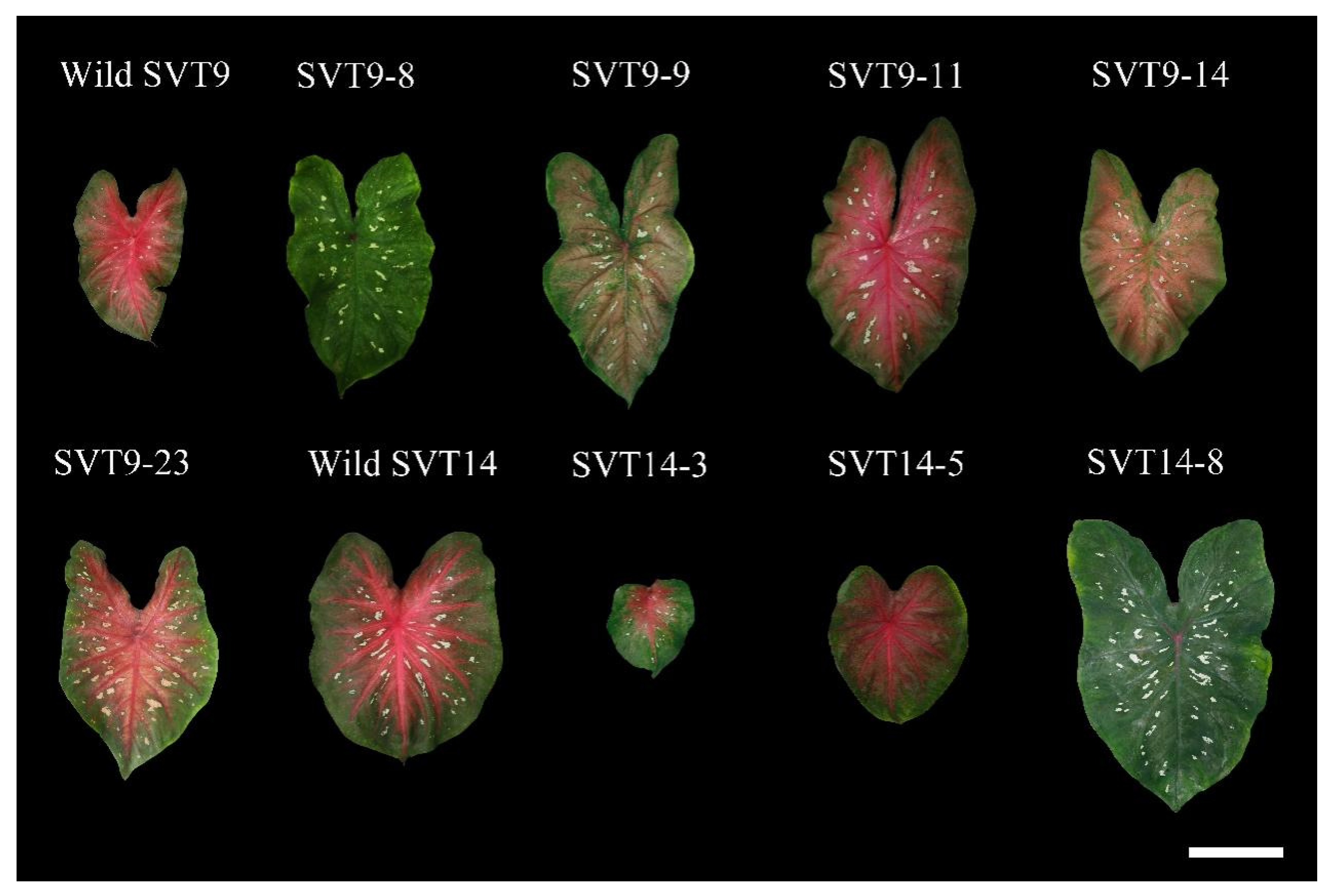
3.2. Comparisons of the Leaf Color Patterns and the Plant Performance among the Regenerated Plants
3.3. Morphological and Stomatal Characteristics among Morphological Variants
3.4. Relative DNA Content Variation
3.5. Chromosome Number Changes
3.6. SSR Identification of the Resulting Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Henry, I.M.; Dilkes, B.P.; Miller, E.S.; Burkart-Waco, D.; Comai, L. Phenotypic consequences of aneuploidy in Arabidopsis thaliana. Genetics 2010, 186, 1231–1245. [Google Scholar] [CrossRef] [Green Version]
- Birchler, J.A. Aneuploidy in plants and flies: The origin of studies of genomic imbalance. Semin. Cell Dev. Biol. 2013, 24, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.; Wu, T.; Liang, G.; Wu, D.; Guo, Q. Identification and characterization of a loquat aneuploid with novel leaf phenotypes. HortScience 2019, 54, 804–808. [Google Scholar] [CrossRef] [Green Version]
- Siegel, J.J.; Amon, A. New insights into the troubles of aneuploidy. Annu. Rev. Cell Dev. Biol. 2012, 28, 189–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Wang, H.; Dong, B.; Yang, X.; Chen, S.; Jiang, J.; Chen, F. Morphological, genome and gene expression changes in newly induced autopolyploid Chrysanthemum lavandulifolium (Fisch. ex Trautv.) Makino. Int. J. Mol. Sci. 2016, 17, 1690. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.; Wang, T.; Yue, F.; Harun, A.; Zhu, B.; Qian, W.; Li, Z. Production and cytology of Brassica autoallohexaploids with two and four copies of two subgenomes. Theor. Appl. Genet. 2022, 135, 2641–2653. [Google Scholar] [CrossRef]
- Evtushenko, E.V.; Lipikhina, Y.A.; Stepochkin, P.I.; Vershinin, A.V. Cytogenetic and molecular characteristics of rye genome in octoploid triticale (× Triticosecale Wittmack). Comp. Cytogenet. 2019, 13, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Cao, Z.; Xu, S.; Deng, Z. Induction, regeneration and characterization of tetraploids and variants in ‘Tapestry’ caladium. Plant Cell Tissue Organ Cult. 2015, 120, 689–700. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Chen, J.J.; Cao, Y.M.; Duan, J.X.; Cai, X.D. Induction of tetraploids in ‘Red Flash’ caladium using colchicine and oryzalin: Morphological, cytological, photosynthetic and chilling tolerance analysis. Sci. Hortic. 2020, 272, 109524. [Google Scholar] [CrossRef]
- Bairu, M.W.; Aremu, A.O.; Van Staden, J. Somaclonal variation in plants: Causes and detection methods. Plant. Growth. Regul. 2011, 63, 147–173. [Google Scholar] [CrossRef]
- Chen, J.J.; Zhang, Y.S.; Duan, J.X.; Cao, Y.M.; Cai, X.D. Morphological, cytological, and pigment analysis of leaf color variants regenerated from long-term subcultured caladium callus. In Vitro Cell Dev. Biol. Plant 2021, 57, 60–71. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, Y.; Sun, S.; Li, G.; Wang, J.; Wang, B.; Lin, X.; Huang, M.; Gong, Z.; Sanguinet, K.A.; et al. Aneuploidization under segmental allotetraploidy in rice and its phenotypic manifestation. Theor. Appl. Genet. 2018, 131, 1273–1285. [Google Scholar] [CrossRef] [Green Version]
- Husband, B.C. Chromosomal variation in plant evolution. Am. J. Bot. 2004, 91, 621–625. [Google Scholar] [CrossRef]
- Henry, I.M.; Dilkes, B.P.; Young, K.; Watson, B.; Wu, H.; Comai, L. Aneuploidy and genetic variation in the Arabidopsis thaliana triploid response. Genetics 2005, 170, 1979–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrish, S.B.; Deng, Z. Discovery and characterization of novel fertile triploids and a new chromosome number in Caladium (Caladium × hortulanum). HortScience 2022, 57, 1078–1085. [Google Scholar] [CrossRef]
- Ehdaie, B.; Waines, J.G. Chromosomal location of genes influencing plant characters and evapotranspiration efficiency in bread wheat. Euphytica 1997, 96, 363–375. [Google Scholar] [CrossRef]
- Bhatia, R.; Sharma, K.; Parkash, C.; Pramanik, A.; Singh, D.; Singh, S.; Dey, S.S. Microspore derived population developed from an inter-specific hybrid (Brassica oleracea × B. carinata) through a modified protocol provides insight into B genome derived black rot resistance and inter-genomic interaction. Plant Cell Tissue Organ Cult. 2021, 145, 417–434. [Google Scholar]
- Meghwal, P.R.; Singh, S.K.; Sharma, H.C. Micropropagation of aneuploid guava. Indian J. Hortic. 2003, 60, 29–33. [Google Scholar]
- Gong, Z.; Xue, C.; Zhou, Y.; Zhang, M.; Liu, X.; Shi, G.; Gu, M. Molecular cytological characterization of somatic variation in rice aneuploids. Plant Mol. Biol. Rep. 2013, 31, 1242–1248. [Google Scholar] [CrossRef]
- Lin, W.; Xiao, X.; Sun, W.; Liu, S.; Wu, Q.; Yao, Y.; Zhang, H.; Zhang, X. Genome-wide identification and expression analysis of cytosine DNA methyltransferase genes related to somaclonal variation in Pineapple (Ananas comosus L.). Agronomy 2022, 12, 1039. [Google Scholar] [CrossRef]
- Deng, Z. Caladium genetics and breeding: Recent advances. Floric. Ornam. Biotechnol. 2012, 6, 53–61. [Google Scholar]
- Wilfret, G.J. Caladium. In The Physiology of Flower Bulbs; Elsevier: Amsterdam, The Netherlands, 1993; pp. 239–247. [Google Scholar]
- Cao, Z.; Deng, Z.; Mclaughlin, M. Interspecific genome size and chromosome number variation shed new light on species classification and evolution in Caladium. J. Am. Soc. Hortic. Sci. 2014, 139, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, E.U.; Hayashi, T.; Yazawa, S. Auxins increase the occurrence of leaf-colour variants in Caladium regenerated from leaf explants. Sci. Hortic. 2004, 100, 153–159. [Google Scholar] [CrossRef]
- Cao, Z.; Sui, S.; Cai, X.; Yang, Q.; Deng, Z. Somaclonal variation in ‘Red Flash’ caladium: Morphological, cytological and molecular characterization. Plant Cell Tissue Organ Cult. 2016, 126, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Deng, Z. Morphological, cytological and molecular marker analyses of ‘Tapestry’ caladium variants reveal diverse genetic changes and enable association of leaf coloration pattern loci with molecular markers. Plant Cell Tissue Organ Cult. 2020, 143, 363–375. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–479. [Google Scholar] [CrossRef]
- Loureiro, J.; Rodriguez, E.; Doležel, J.; Santos, C. Two new nuclear isolation buffers for plant DNA flow cytometry: A test with 37 species. Ann. Bot. 2007, 100, 875–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, L.; Deng, Z. Development and characterization of microsatellite markers for caladiums (Caladium Vent.). Plant Breed. 2011, 130, 591–595. [Google Scholar] [CrossRef]
- Fulton, T.M.; Chunzoongse, J.; Tanksley, S.D. Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep. 1995, 13, 207–209. [Google Scholar] [CrossRef]
- Bassam, B.J.; Caetano-Anollés, G.; Gresshoff, P.M. Fast and sensitive silver staining of DNA in polyacrylamide gels. Anal. Biochem. 1991, 196, 80–83. [Google Scholar] [CrossRef]
- Marfil, C.F.; Duarte, P.F.; Masuelli, R.W. Phenotypic and epigenetic variation induced in newly synthesized allopolyploids and autopolyploids of potato. Sci. Hortic. 2018, 234, 101–109. [Google Scholar] [CrossRef]
- Gao, L.; Diarso, M.; Zhang, A.; Zhang, H.; Dong, Y.; Liu, L.; Liu, B. Heritable alteration of DNA methylation induced by whole-chromosome aneuploidy in wheat. New Phytol. 2016, 209, 364–375. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Harbaugh, B.K. Independent inheritance of leaf shape and main vein color in caladium. J. Am. Soc. Hortic. Sci. 2006, 131, 53–58. [Google Scholar] [CrossRef]
- Deng, Z.; Goktepe, F.; Harbaugh, B.K. Inheritance of leaf spots and their genetic relationships with leaf shape and vein color in caladium. J. Am. Soc. Hortic. Sci. 2008, 133, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Syeed, R.; Mujib, A.; Malik, M.Q.; Gulzar, B.; Zafar, N.; Mamgain, J.; Ejaz, B. Direct somatic embryogenesis and flow cytometric assessment of ploidy stability in regenerants of Caladium × hortulanum ‘Fancy’. J. Appl. Genet. 2022, 63, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Sabooni, N.; Gharaghani, A. Induced polyploidy deeply influences reproductive life cycles, related phytochemical features, and phytohormonal activities in blackberry species. Front. Plant. Sci. 2022, 13, 938284. [Google Scholar] [CrossRef]
- Huettel, B.; Kreil, D.P.; Matzke, M.; Matzke, A.J.M. Effects of aneuploidy on genome structure, expression, and interphase organization in Arabidopsis thaliana. PLoS Genet. 2008, 4, e1000226. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Code | Leaf Shape | Main Veins | Interveinal Areas | Leaf Margins | Leaf Spots |
|---|---|---|---|---|---|
| Wild SVT9 | Fancy | Red | Red | Green | White |
| SVT9-8 | Fancy | Green | Green | Light green | White |
| SVT9-9 | Fancy | Green | Purple-red | Light green | White |
| SVT9-11 | Fancy | Red | Red | Green | White and pinkish-red |
| SVT9-14 | Fancy | Green | Pinkish-red | Green | Pinkish-red |
| SVT9-23 | Fancy | Red | Red | Green | Pinkish-red |
| Wild SVT14 | Fancy | Red | Red | Green | White |
| SVT14-3 | Fancy | Red | Red | Light green | White |
| SVT14-5 | Fancy | Red | Red | Green | No spots |
| SVT14-8 | Fancy | Green | Green | Light green | White |
| Plant Code | Plant Height (cm) | Leaf Number per Plant | Leaf Length (cm) | Leaf Width (cm) | Leaf Length/Width Ratio | Leaf Thickness (mm) |
|---|---|---|---|---|---|---|
| Wild SVT9 | 26.2 | 6 | 18.67 ± 0.15 d | 11.23 ± 0.42 f | 1.66 ± 0.05 b | 0.26 ± 0.00 b |
| SVT9-8 | 34.2 | 5 | 25.40 ± 5.99 bc | 12.90 ± 0.26 e | 1.96 ± 0.42 a | 0.26 ± 0.02 b |
| SVT9-9 | 45.6 | 5 | 27.77 ± 1.33 ab | 16.23 ± 0.72 c | 1.71 ± 0.12 ab | 0.27 ± 0.02 b |
| SVT9-11 | 37.4 | 6 | 27.00 ± 1.08 bc | 16.73 ± 0.91 c | 1.61 ± 0.12 b | 0.25 ± 0.02 b |
| SVT9-14 | 30.5 | 6 | 23.63 ± 0.91 bc | 14.30 ± 0.85 d | 1.65 ± 0.04 b | 0.25 ± 0.02 b |
| SVT9-23 | 30.2 | 4 | 24.87 ± 1.05 bc | 16.70 ± 0.53 c | 1.49 ± 0.04 bc | 0.27 ± 0.02 b |
| Wild SVT14 | 33.4 | 5 | 23.43 ± 0.55 c | 19.00 ± 0.36 b | 1.23 ± 0.02 cd | 0.35 ± 0.02 a |
| SVT14-3 | 18.6 | 2 | 9.90 ± 1.98 e | 8.01 ± 1.56 g | 1.24 ± 0.01 cd | 0.35 ± 0.01 a |
| SVT14-5 | 22.7 | 4 | 17.07 ± 0.64 d | 14.70 ± 0.95 d | 1.16 ± 0.03 d | 0.33 ± 0.01 a |
| SVT14-8 | 51.5 | 3 | 31.50 ± 1.01 a | 21.13 ± 1.05 a | 1.47 ± 0.12 bc | 0.27 ± 0.02 b |
| Plant Code | Stomatal Guard Cell Length (μm) | Stomatal Guard Cell Width (μm) | Stomatal Density (no./mm2) |
|---|---|---|---|
| Wild SVT9 | 27.88 ± 2.69 cd | 19.08 ± 1.82 bc | 112.12 ± 4.34 b |
| SVT9-8 | 28.40 ± 2.33 bcd | 17.98 ± 2.05 bc | 96.21 ± 9.56 c |
| SVT9-9 | 27.02 ± 0.94 d | 18.57 ± 1.65 bc | 129.49 ± 10.92 a |
| SVT9-11 | 29.16 ± 1.77 bc | 17.07 ± 1.97 c | 122.97 ± 5.01 ab |
| SVT9-14 | 24.51 ± 0.87 e | 17.48 ± 2.19 c | 88.25 ± 3.74 c |
| SVT9-23 | 33.51 ± 2.85 a | 18.91 ± 4.69 bc | 122.97 ± 13.67 ab |
| Wild SVT14 | 33.67 ± 2.86 a | 20.65 ± 4.05 b | 68.10 ± 13.05 d |
| SVT14-3 | 30.24 ± 1.76 b | 20.74 ± 1.21 b | 68.72 ± 7.23 d |
| SVT14-5 | 34.90 ± 1.62 a | 23.81 ± 4.17 a | 67.27 ± 6.42 d |
| SVT14-8 | 34.65 ± 1.99 a | 20.77 ± 1.65 b | 70.17 ± 12.36 d |
| Plant Code | Mean Fluorescence Intensity (MFI) | MFI Change Compared to the Corresponding Wild Type (%) | Chromosome Number Observed in Somatic Cells (2n) |
|---|---|---|---|
| Wild SVT9 | 361,547.4 ± 275.9 | – | 2x − 2 = 28 |
| SVT9-1 | 369,554.8 ± 3243.8 ns | +2.21 | 2x − 2 = 28 |
| SVT9-2 | 360,779.3 ± 5688.5 ns | −0.21 | 2x − 2 = 28 |
| SVT9-3 | 363,844.3 ± 3293.5 ns | +0.64 | 2x − 2 = 28 |
| SVT9-4 | 401,378.1 ± 2886.3 * | +11.02 | 2x = 30 |
| SVT9-5 | 364,728.6 ± 4364.3 ns | +0.88 | 2x − 2 = 28 |
| SVT9-6 | 366,356.2 ± 1425.4 ns | +1.33 | 2x − 2 = 28 |
| SVT9-7 | 356,318.6 ± 5712.4 ns | −1.45 | 2x − 2 = 28 |
| SVT9-8 | 348,594.2 ± 993.8 * | −3.58 | 2x − 3 = 27 |
| SVT9-9 | 378,007.6 ± 4449.3 * | +4.55 | 2x − 1 = 29 |
| SVT9-10 | 364,349.8 ± 2577.7 ns | +0.78 | 2x − 2 = 28 |
| SVT9-11 | 380,383.7 ± 2924.5 * | +5.21 | 2x − 1 = 29 |
| SVT9-12 | 369,176.3 ± 8997.0 ns | +2.11 | 2x − 2 = 28 |
| SVT9-13 | 371,075.8 ± 4186.6 ns | +2.64 | 2x − 2 = 28 |
| SVT9-14 | 350,182.6 ± 555.9 * | −3.14 | 2x − 3 = 27 |
| SVT9-15 | 368,263.1 ± 2776.1 ns | +1.86 | 2x − 2 = 28 |
| SVT9-16 | 370,699.7 ± 2459.0 ns | +2.53 | 2x − 2 = 28 |
| SVT9-17 | 364,147.3 ± 6699.7 ns | +0.72 | 2x − 2 = 28 |
| SVT9-18 | 359,826.5 ± 1295.5 ns | −0.48 | 2x − 2 = 28 |
| SVT9-19 | 372,395.5 ± 532.5 ns | +3.00 | 2x − 2 = 28 |
| SVT9-20 | 370,988.0 ± 3242.0 ns | +2.61 | 2x − 2 = 28 |
| SVT9-21 | 371,636.7 ± 2274.3 ns | +2.79 | 2x − 2 = 28 |
| SVT9-22 | 362,934.1 ± 16,053.1 ns | +0.38 | 2x − 2 = 28 |
| SVT9-23 | 378,928.2 ± 1686.4 * | +4.81 | 2x = 30 |
| Wild SVT14 | 678,715.4 ± 8091.9 | – | 4x − 6 = 54 |
| SVT14-1 | 726,124.5 ± 2163.8 * | +6.99 | 4x − 4 = 56 |
| SVT14-2 | 689,727.1 ± 15,919.7 ns | +1.62 | 4x − 6 = 54 |
| SVT14-3 | 624,862.1 ± 2061.8 * | −7.93 | 4x − 10 = 50 |
| SVT14-4 | 673,317.1 ± 10,184.6 ns | −0.80 | 4x − 6 = 54 |
| SVT14-5 | 615,617.3 ± 5015.4 * | −9.30 | 4x − 10 = 50 |
| SVT14-6 | 684,457.9 ± 8992.9 ns | +0.85 | 4x − 6 = 54 |
| SVT14-7 | 668,038.6 ± 2779.5 ns | −1.57 | 4x − 6 = 54 |
| SVT14-8 | 640,112.3 ± 3472.5 * | −5.69 | 4x − 10 = 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Zhao, X.; Wang, Y.; Jiang, D.; Zhang, Y.; Hu, L.; Liu, Y.; Cai, X. Morphological, Cytological, and Molecular-Based Genetic Stability Analysis of In Vitro-Propagated Plants from Newly Induced Aneuploids in Caladium. Agriculture 2022, 12, 1708. https://doi.org/10.3390/agriculture12101708
Yu S, Zhao X, Wang Y, Jiang D, Zhang Y, Hu L, Liu Y, Cai X. Morphological, Cytological, and Molecular-Based Genetic Stability Analysis of In Vitro-Propagated Plants from Newly Induced Aneuploids in Caladium. Agriculture. 2022; 12(10):1708. https://doi.org/10.3390/agriculture12101708
Chicago/Turabian StyleYu, Shuangying, Xiaoqin Zhao, Yida Wang, Dongzhu Jiang, Yiming Zhang, Liu Hu, Yiqing Liu, and Xiaodong Cai. 2022. "Morphological, Cytological, and Molecular-Based Genetic Stability Analysis of In Vitro-Propagated Plants from Newly Induced Aneuploids in Caladium" Agriculture 12, no. 10: 1708. https://doi.org/10.3390/agriculture12101708