
Assessing Nitrogen Availability in Biobased Fertilizers: Effect of Vegetation on Mineralization Patterns



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Setup
2.2. Soil Collection and Analyses
2.3. Biobased Fertilizer Collection and Analyses
2.4. Laboratory Incubation and Sampling
2.5. Maize-Growing Experiment and Sampling
2.6. Calculation on N Release and Mineralization
2.7. Mass Balance Calculation
2.8. Statistical Analyses
3. Results
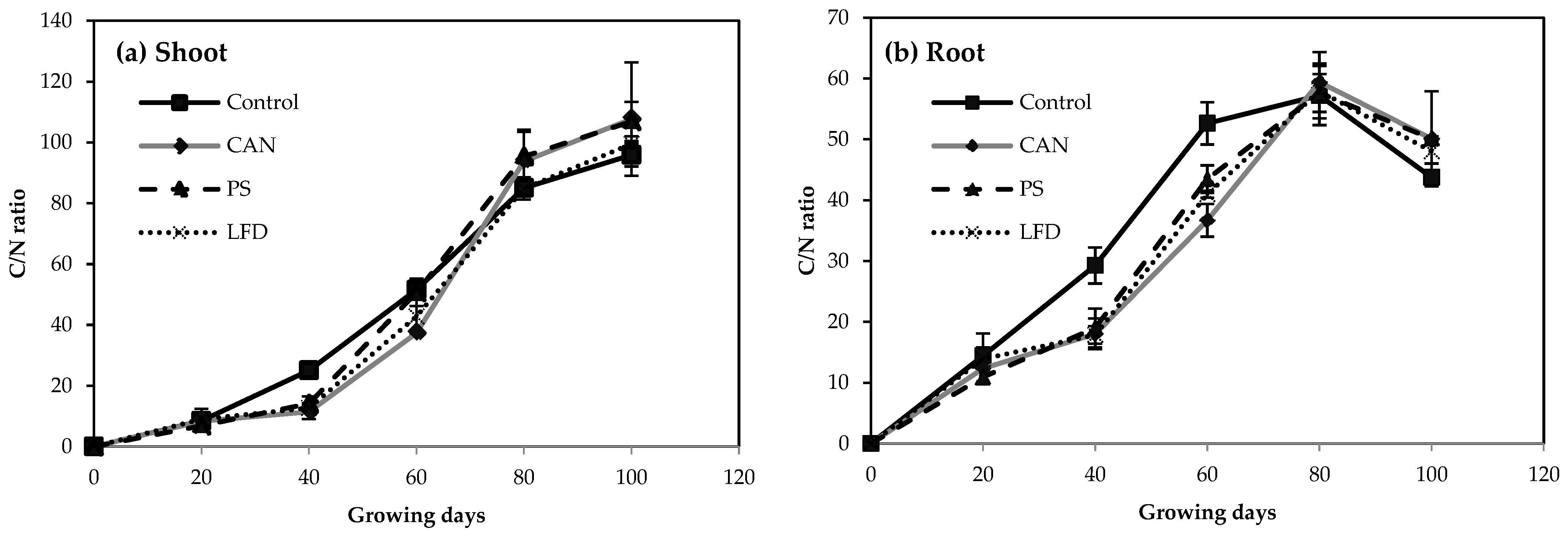
3.1. Root and Shoot Development of Plants
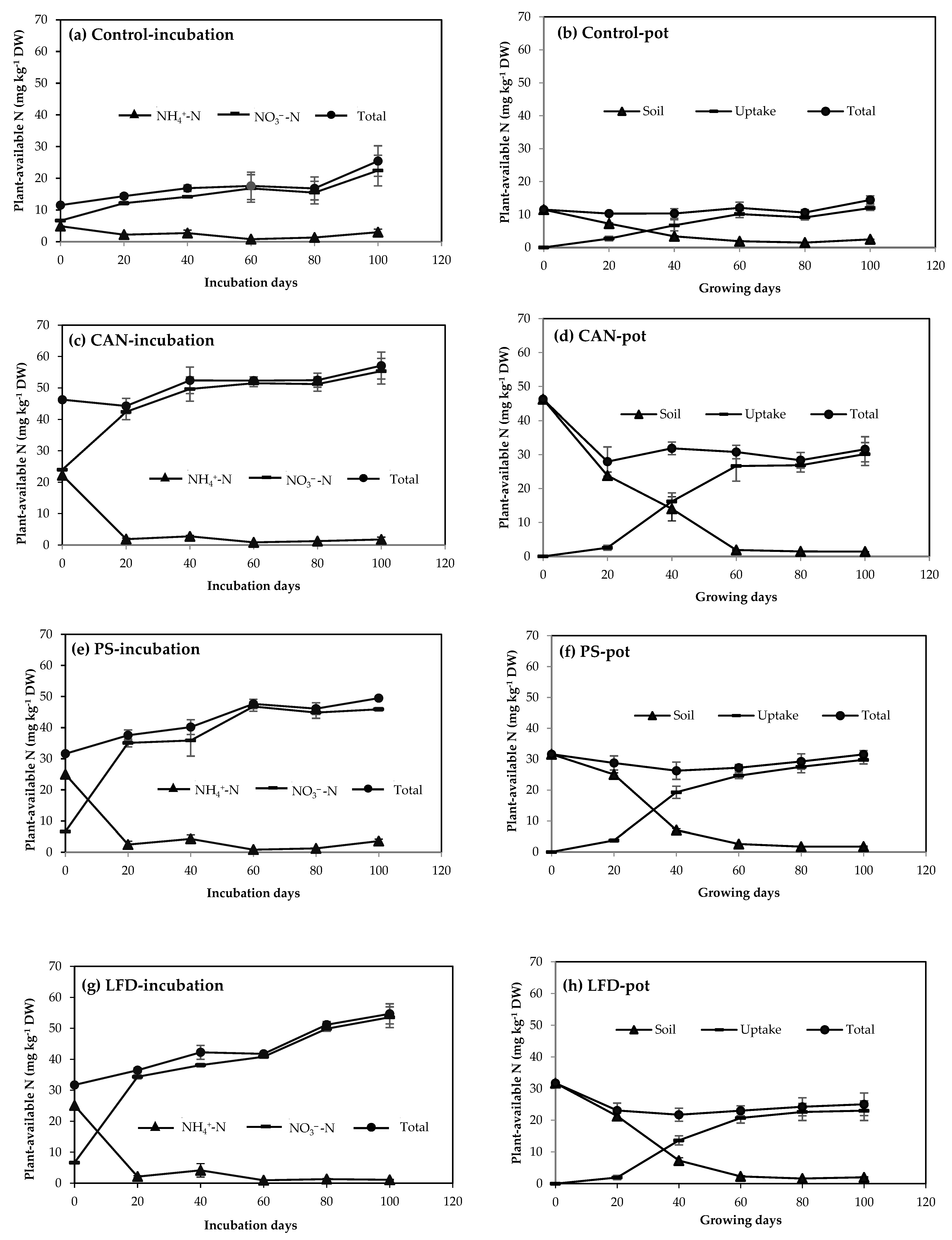
3.2. N Mineralization in Soil with and without Vegetation
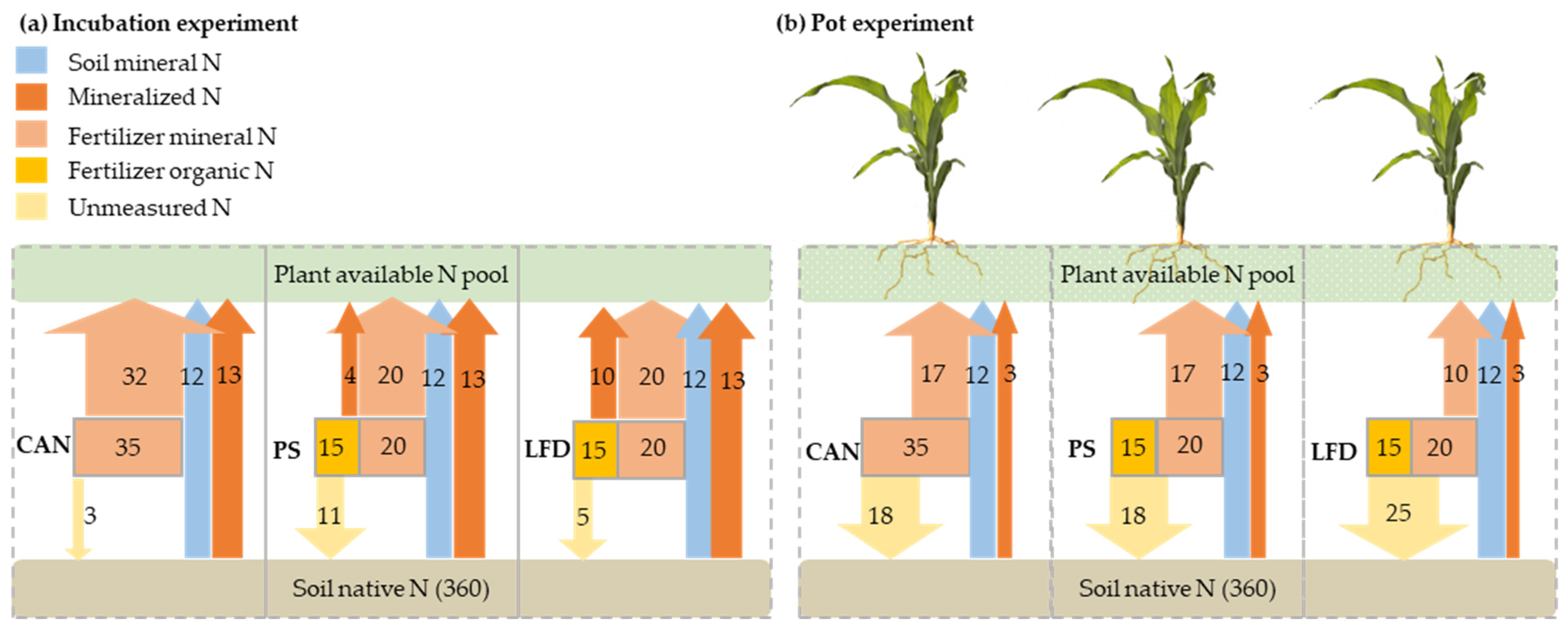
3.3. Nitrogen Mass Balance
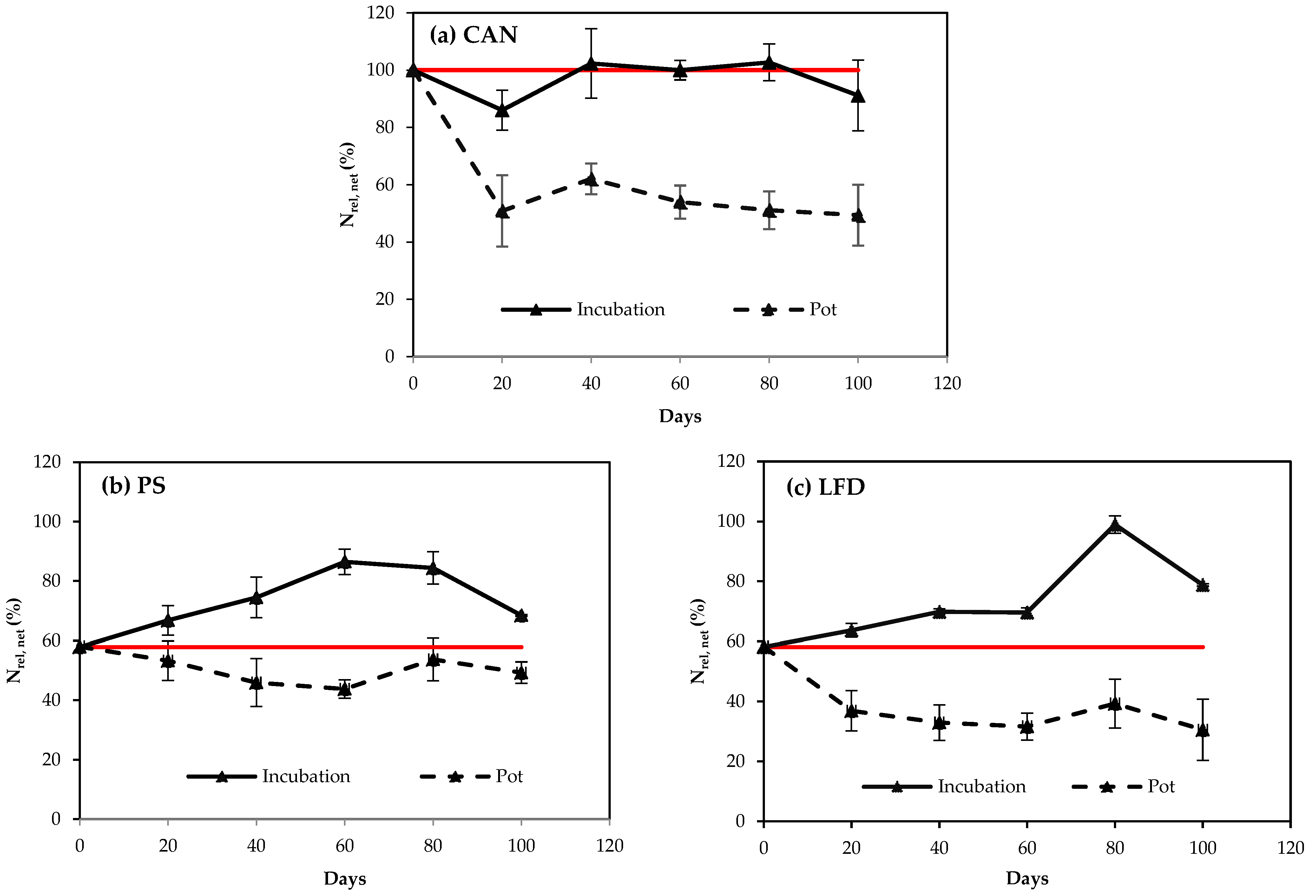
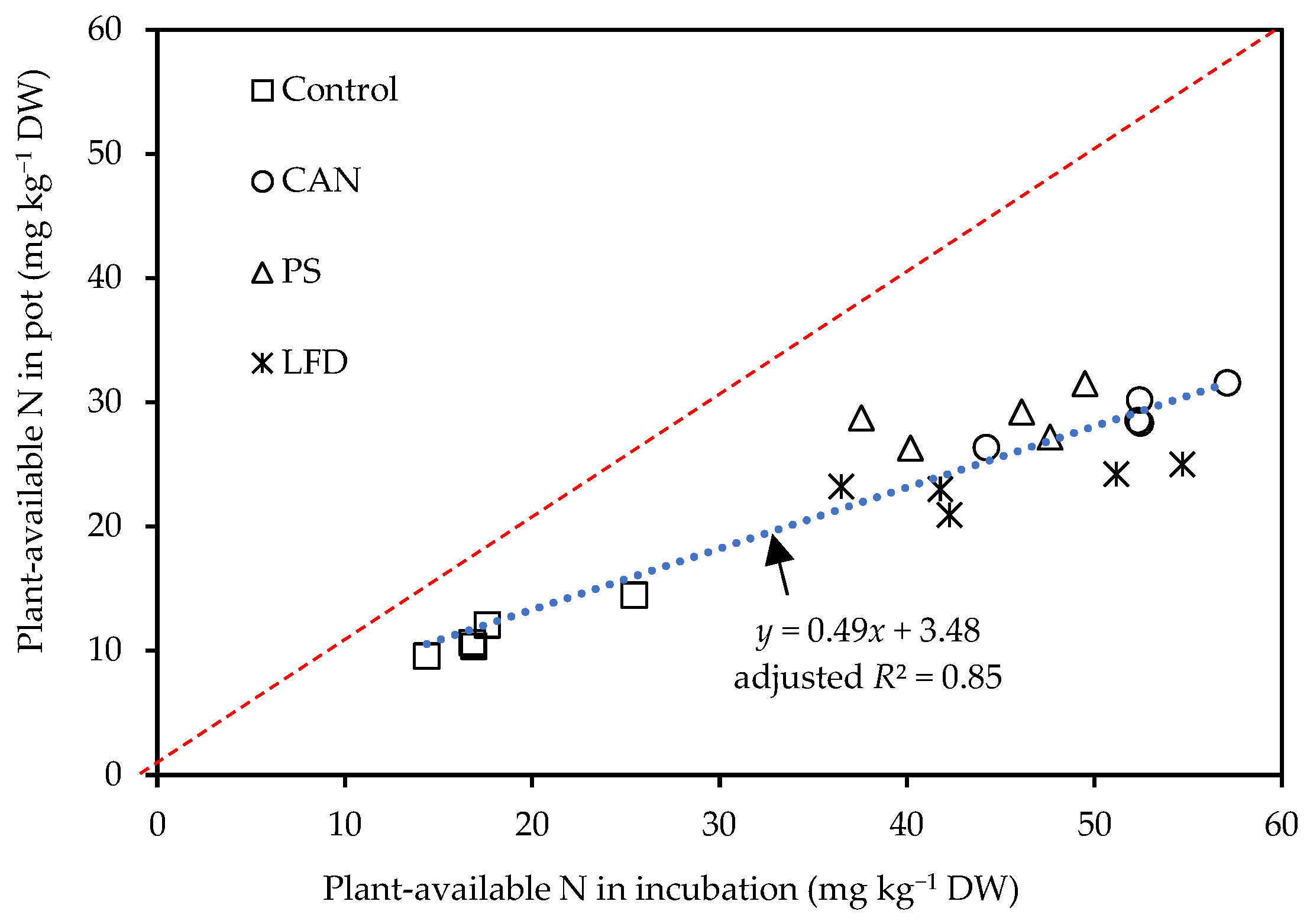
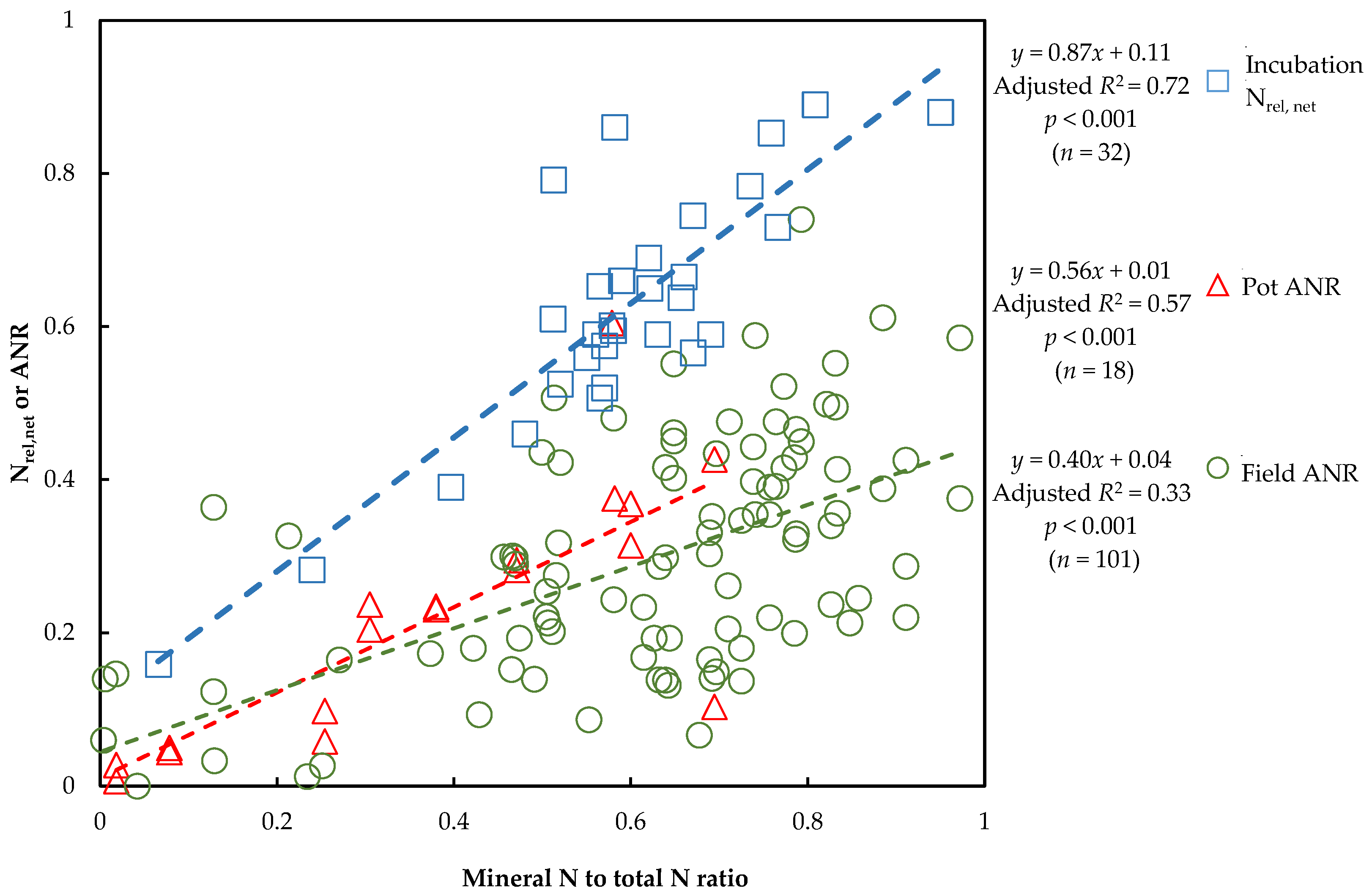
3.4. Link of N Availability between Vegetation and Non-Vegetation
4. Discussion
4.1. Effect of Vegetation on N Mineralization
4.2. Effect of Fertilization on N Mineralization
4.3. Effect of N Deficiency on Plant Growth and N Dynamics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilland, B. Is a Haber-Bosch World sustainable? Population, nutrition, cereals, nitrogen and environment. J. Soc. Political Econ. Stud. 2014, 39, 166. [Google Scholar]
- Leip, A.; Weiss, F.; Lesschen, J.P.; Westhoek, H. The nitrogen footprint of food products in the European Union. J. Agric. Sci. 2014, 152, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Klop, G.; Velthof, G.L.; Van Groenigen, J.W. Application technique affects the potential of mineral concentrates from livestock manure to replace inorganic nitrogen fertilizer. Soil Use Manag. 2012, 28, 468–477. [Google Scholar] [CrossRef]
- Sigurnjak, I. Animal Manure Derivatives as Alternatives for Synthetic Nitrogen Fertilizers. Ph.D. Thesis, Faculty of Bioscience Engineering, Ghent University, Ghent, Belgium, 2017. [Google Scholar]
- Vaneeckhaute, C.; Meers, E.; Michels, E.; Buysse, J.; Tack, F.M.G. Ecological and economic benefits of the application of bio-based mineral fertilizers in modern agriculture. Biomass Bioenergy 2013, 49, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Vaneeckhaute, C.; Ghekiere, G.; Michels, E.; Vanrolleghem, P.A.; Tack, F.M.; Meers, E. Assessing nutrient use efficiency and environmental pressure of macronutrients in biobased mineral fertilizers: A review of recent advances and best practices at field scale. Adv. Agron. 2014, 128, 137–180. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Wu, L.; Wang, J.; Ma, J.; Zheng, N.; Hill, P.W.; Chadwick, D.R.; Jones, D.L. Fertilizer regime changes the competitive uptake of organic nitrogen by wheat and soil microorganisms: An in-situ uptake test using 13C, 15N labelling, and 13C-PLFA analysis. Soil Biol. Biochem. 2018, 125, 319–327. [Google Scholar] [CrossRef]
- Inselsbacher, E.; Umana, N.H.N.; Stange, F.C.; Gorfer, M.; Schüller, E.; Ripka, K.; Wanek, W. Short-term competition between crop plants and soil microbes for inorganic N fertilizer. Soil Biol. Biochem. 2010, 42, 360–372. [Google Scholar] [CrossRef]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; de la Fuente, C.; Bernal, M.P. Chemical properties of anaerobic digestates affecting C and N dynamics in amended soils. Agric. Ecosyst. Environ. 2012, 160, 15–22. [Google Scholar] [CrossRef]
- Bonanomi, G.; Sarker, T.C.; Zotti, M.; Cesarano, G.; Allevato, E.; Mazzoleni, S. Predicting nitrogen mineralization from organic amendments: Beyond C/N ratio by 13 C-CPMAS NMR approach. Plant Soil 2019, 441, 129–146. [Google Scholar] [CrossRef]
- Cheng, W. Rhizosphere priming effect: Its functional relationships with microbial turnover, evapotranspiration, and C–N budgets. Soil Biol. Biochem. 2009, 41, 1795–1801. [Google Scholar] [CrossRef]
- Grunert, O.; Robles-Aguilar, A.A.; Hernandez-Sanabria, E.; Schrey, S.D.; Reheul, D.; Van Labeke, M.C.; Vlaeminck, S.E.; Vandekerckhove, T.G.; Mysara, M.; Monsieurs, P.; et al. Tomato plants rather than fertilizers drive microbial community structure in horticultural growing media. Sci. Rep. 2019, 9, 9561. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Streeter, T.C.; Bol, R. Soil microbes compete effectively with plants for organic-nitrogen inputs to temperate grasslands. Ecology 2003, 84, 1277–1287. [Google Scholar] [CrossRef]
- Ninh, H.T.; Grandy, A.S.; Wickings, K.; Snapp, S.S.; Kirk, W.; Hao, J. Organic amendment effects on potato productivity and quality are related to soil microbial activity. Plant Soil 2015, 386, 223–236. [Google Scholar] [CrossRef]
- Vellinga, T.V.; André, G. Sixty years of Dutch nitrogen fertiliser experiments an overview of the effects of soil type fertiliser input management and of developments in time. NJAS Wagening. J. Life Sci. 1999, 47, 215–241. [Google Scholar] [CrossRef]
- De Neve, S. Organic matter mineralization as a source of nitrogen. In Advances in Research on Fertilization Management of Vegetable Crops; Springer: Cham, Switzerland, 2017; pp. 65–83. [Google Scholar] [CrossRef]
- Rigby, H.; Clarke, B.O.; Pritchard, D.L.; Meehan, B.; Beshah, F.; Smith, S.R.; Porter, N.A. A critical review of nitrogen mineralization in biosolids-amended soil, the associated fertilizer value for crop production and potential for emissions to the environment. Sci. Total Environ. 2016, 541, 1310–1338. [Google Scholar] [CrossRef] [PubMed]
- Coskun, D.; Britto, D.T.; Shi, W.; Kronzucker, H.J. How plant root exudates shape the nitrogen cycle. Trends Plant Sci. 2017, 22, 661–673. [Google Scholar] [CrossRef]
- Meier, I.C.; Finzi, A.C.; Phillips, R.P. Root exudates increase N availability by stimulating microbial turnover of fast-cycling N pools. Soil Biol. Biochem. 2017, 106, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Cesco, S.; Mimmo, T.; Tonon, G.; Tomasi, N.; Pinton, R.; Terzano, R.; Neumann, G.; Weisskopf, L.; Renella, G.; Landi, L.; et al. Plant-borne flavonoids released into the rhizosphere: Impact on soil bio-activities related to plant nutrition. A review. Biol. Fertil. Soils 2012, 48, 123–149. [Google Scholar] [CrossRef]
- Palacios, O.A.; Bashan, Y.; de-Bashan, L.E. Proven and potential involvement of vitamins in interactions of plants with plant growth-promoting bacteria—An overview. Biol. Fertil. Soils 2014, 50, 415–432. [Google Scholar] [CrossRef]
- Czarnes, S.; Hallett, P.D.; Bengough, A.G.; Young, I.M. Root- and microbial-derived mucilages affect soil structure and water transport. Eur. J. Soil Sci. 2000, 51, 435–443. [Google Scholar] [CrossRef]
- Canarini, A.; Dijkstra, F.A. Dry-rewetting cycles regulate wheat carbon rhizodeposition stabilization and nitrogen cycling. Soil Biol. Biochem. 2015, 81, 195–203. [Google Scholar] [CrossRef]
- Qian, J.H.; Doran, J.W.; Walters, D.T. Maize plant contributions to root zone available carbon and microbial transformations of nitrogen. Soil Biol. Biochem. 1997, 29, 1451–1462. [Google Scholar] [CrossRef]
- Buckwell, A.; Nadeu, E. Nutrient Recovery and Reuse (NRR) in European Agriculture. A Review of the Issues, Opportunities, and Actions; RISE Foundation: Brussels, Belgium, 2016. [Google Scholar]
- Möller, K.; Stinner, W.; Deuker, A.; Leithold, G. Effects of different manuring systems with and without biogas digestion on nitrogen cycle and crop yield in mixed organic dairy farming systems. Nutr. Cycl. Agroecosyst. 2008, 82, 209–232. [Google Scholar] [CrossRef]
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Secur. 2011, 3, 307. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility. A Handbook of Methods, 2nd ed.; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Van Ranst, E.; Verloo, M.; Demeyer, A.; Pauwels, J.M. Manual for the Soil Chemistry and Fertility Laboratory: Analytical Methods of Soils and Plants, Equipment, and Management of Consumables; NUGI 835: Ghent, Belgium, 1999; pp. 1–243. ISBN 90-76603-01-4. [Google Scholar]
- VITO. Bodem-Bepaling van Snel Vrijkomende Organische Stikstof. Available online: https://esites.vito.be/sites/reflabos/2019/Online%20documenten/BAM-deel1-12.pdf (accessed on 7 September 2021).
- Kirkham, D.O.N.; Bartholomew, W.V. Equations for following nutrient transformations in soil, utilizing tracer data. Soil Sci. Soc. Am. J. 1954, 18, 33–34. [Google Scholar] [CrossRef]
- De Neve, S.; Hofman, G. Modelling N mineralization of vegetable crop residues during laboratory incubations. Soil Biol. Biochem. 1996, 8, 1451–1457. [Google Scholar] [CrossRef]
- Kirchmann, H.; Lundvall, A. Relationship between N immobilization and volatile fatty acids in soil after application of pig and cattle slurry. Biol. Fertil. Soils 1993, 15, 161–164. [Google Scholar] [CrossRef]
- Sørensen, P.; Weisbjerg, M.R.; Lund, P. Dietary effects on the composition and plant utilization of nitrogen in dairy cattle manure. J. Agric. Sci. 2003, 141, 79–91. [Google Scholar] [CrossRef]
- Morvan, T.; Nicolardot, B. Role of organic fractions on C decomposition and N mineralization of animal wastes in soil. Biol. Fertil. Soils 2009, 45, 477–486. [Google Scholar] [CrossRef]
- Sigurnjak, I.; De Waele, J.; Michels, E.; Tack, F.; Meers, E.; De Neve, S. Nitrogen release and mineralization potential of derivatives from nutrient recovery processes as substitutes for fossil fuel-based nitrogen fertilizers. Soil Use Manag. 2017, 33, 437–446. [Google Scholar] [CrossRef]
- Cavalli, D.; Bechini, L.; Di Matteo, A.; Corti, M.; Ceccon, P.; Marino Gallina, P. Nitrogen availability after repeated additions of raw and anaerobically digested 15N-labelled pig slurry. Eur. J. Soil Sci. 2018, 69, 1044–1055. [Google Scholar] [CrossRef]
- Maurer, C.; Seiler-Petzold, J.; Schulz, R.; Müller, J. Short-Term Nitrogen Uptake of Barley from Differently Processed Biogas Digestate in Pot Experiments. Energies 2019, 12, 696. [Google Scholar] [CrossRef] [Green Version]
- Díez, J.A.; Hernaiz, P.; Muñoz, M.J.; De la Torre, A.; Vallejo, A. Impact of pig slurry on soil properties water salinization nitrate leaching and crop yield in a four-year experiment in Central Spain. Soil Use Manag. 2004, 20, 444–450. [Google Scholar] [CrossRef] [Green Version]
- Chantigny, M.H.; Angers, D.A.; Rochette, P.; Bélanger, G.; Massé, D.; Côté, D. Gaseous nitrogen emissions and forage nitrogen uptake on soils fertilized with raw and treated swine manure. J. Environ. Qual. 2007, 36, 1864–1872. [Google Scholar] [CrossRef] [Green Version]
- Chantigny, M.H.; Angers, D.A.; Bélanger, G.; Rochette, P.; Eriksen-Hamel, N.; Bittman, S.; Gasser, M.O. Yield and nutrient export of grain corn fertilized with raw and treated liquid swine manure. Agron. J. 2008, 100, 1303–1309. [Google Scholar] [CrossRef]
- Saunders, O.E.; Fortuna, A.M.; Harrison, J.H.; Whitefield, E.; Cogger, C.G.; Kennedy, A.C.; Bary, A.I. Comparison of raw dairy manure slurry and anaerobically digested slurry as N sources for grass forage production. Int. J. Agron. 2012, 2012, 101074. [Google Scholar] [CrossRef]
- Gagnon, B.; Ziadi, N.; Chantigny, M.H.; Bélanger, G.; Massé, D.I. Biosolids from treated swine manure and papermill residues affect corn fertilizer value. Agron. J. 2012, 104, 483–492. [Google Scholar] [CrossRef]
- Cavalli, D.; Cabassi, G.; Borrelli, L.; Geromel, G.; Bechini, L.; Degano, L.; Gallina, P.M. Nitrogen fertilizer replacement value of undigested liquid cattle manure and digestates. Eur. J. Agron. 2016, 73, 34–41. [Google Scholar] [CrossRef]
- Tampio, E.; Salo, T.; Rintala, J. Agronomic characteristics of five different urban waste digestates. J. Environ. Manag. 2016, 169, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Federolf, C.P.; Westerschulte, M.; Olfs, H.W.; Broll, G.; Trautz, D. Nitrogen dynamics following slurry injection in maize: Crop development. Nutr. Cycl. Agroecosyst. 2017, 107, 19–31. [Google Scholar] [CrossRef]
- Zhang, X.; Davidson, E.A.; Mauzerall, D.L.; Searchinger, T.D.; Dumas, P.; Shen, Y. Managing nitrogen for sustainable development. Nature 2015, 528, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Fuente, C.; Clemente, R.; Martinez, J.; Bernal, M.P. Optimization of pig slurry application to heavy polluted soils monitoring nitrification processes. Chemosphere 2010, 81, 603–610. [Google Scholar] [CrossRef]
- Liu, G.D.; Li, Y.C.; Alva, A.K. High Water Regime Can Reduce Ammonia Volatilization from Soils under Potato Production. Commun. Soil Sci. Plant Anal. 2007, 38, 1203–1220. [Google Scholar] [CrossRef]
- Sørensen, P.; Amato, M. Remineralisation and residual effects of N after application of pig slurry to soil. Eur. J. Agron. 2002, 16, 81–95. [Google Scholar] [CrossRef]
- Malique, F.; Ke, P.; Boettcher, J.; Dannenmann, M.; Butterbach-Bahl, K. Plant and soil effects on denitrification potential in agricultural soils. Plant Soil 2019, 439, 459–474. [Google Scholar] [CrossRef]
- Sierra, J. Temperature and soil moisture dependence of N mineralization in intact soil cores. Soil Biol. Biochem. 1997, 29, 1557–1563. [Google Scholar] [CrossRef]
- Philippot, L.; Hallin, S.; Borjesson, G.; Baggs, E.M. Biochemical cycling in the rhizosphere having an impact on global change. Plant Soil 2009, 321, 61–81. [Google Scholar] [CrossRef]
- Rummel, P.S.; Well, R.; Pfeiffer, B.; Dittert, K.; Floßmann, S.; Pausch, J. Nitrate uptake and carbon exudation–do plant roots stimulate or inhibit denitrification? Plant Soil 2021, 459, 217–233. [Google Scholar] [CrossRef]
- D’haene, K.; Moreels, E.; De Neve, S.; Daguilar, B.C.; Boeckx, P.; Hofman, G.; Van Cleemput, O. Soil properties influencing the denitrification potential of Flemish agricultural soils. Biol. Fertil. Soils 2003, 38, 358–366. [Google Scholar] [CrossRef]
- Mikha, M.M.; Rice, C.W.; Milliken, G.A. Carbon and nitrogen mineralization as affected by drying and wetting cycles. Soil Biol. Biochem. 2005, 37, 339–347. [Google Scholar] [CrossRef]
- Sigurnjak, I.; Michels, E.; Crappé, S.; Buysens, S.; Biswas, J.K.; Tack, F.M.; De Neve, S.; Meers, E. Does acidification increase the nitrogen fertilizer replacement value of bio-based fertilizers? J. Plant Nutr. Soil Sci. 2017, 180, 800–810. [Google Scholar] [CrossRef]
- Möller, K. Effects of anaerobic digestion on soil carbon and nitrogen turnover, N emissions, and soil biological activity. A review. Agron. Sustain. Dev. 2015, 35, 1021–1041. [Google Scholar] [CrossRef]
- Dilly, O. Ratios of microbial biomass estimates to evaluate microbial physiology in soil. Biol. Fertil. Soils 2006, 42, 241–246. [Google Scholar] [CrossRef]
- Cheng, W.; Johnson, D.W.; Fu, S. Rhizosphere Effects on Decomposition: Controls of Plant Species, Phenology, and Fertilization. Soil Sci. Soc. Am. J. 2003, 67, 1418–1427. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Carrillo, Y.; Pendall, E.; Morgan, J.A. Rhizosphere priming: A nutrient perspective. Front. Microbiol. 2013, 4, 216. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; Van Der Heijden, M.G. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abubaker, J.; Risberg, K.; Jönsson, E.; Dahlin, A.S.; Cederlund, H.; Pell, M. Short-term effects of biogas digestates and pig slurry application on soil microbial activity. Appl. Environ. Soil Sci. 2015. [Google Scholar] [CrossRef] [Green Version]
- Maciel de Oliveira, S.; Almeida, R.E.M.D.; Ciampitti, I.A.; Pierozan Junior, C.; Lago, B.C.; Trivelin, P.C.O.; Favarin, J.L. Understanding N timing in corn yield and fertilizer N recovery: An insight from an isotopic labeled-N determination. PLoS ONE 2018, 13, e0192776. [Google Scholar] [CrossRef] [Green Version]
- Schröder, J.J.; Uenk, D.; Hilhorst, G.J. Long-term nitrogen fertilizer replacement value of cattle manures applied to cut grassland. Plant Soil 2007, 299, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Sharma, M.K. Nutrient deficiencies in cereal crops: Maize (Zea mays Linn.). In Nutrient Deficiencies of Field Crops: Guide to Diagnosis and Management; CABI: Rajasthan, India, 2013; pp. 25–45. ISBN 9781780642789. [Google Scholar]
- Plénet, D.; Lemaire, G. Relationships between dynamics of nitrogen uptake and dry matter accumulation in maize crops. Determination of critical N concentration. Plant Soil 1999, 216, 65–82. [Google Scholar] [CrossRef]
- Peng, Y.; Niu, J.; Peng, Z.; Zhang, F.; Li, C. Shoot growth potential drives N uptake in maize plants and correlates with root growth in the soil. Field Crops Res. 2010, 115, 85–93. [Google Scholar] [CrossRef]
- Rengel, Z.; Marschner, P. Nutrient availability and management in the rhizosphere: Exploiting genotypic differences. New Phytol. 2005, 168, 305–312. [Google Scholar] [CrossRef]
- Yu, P.; White, P.J.; Hochholdinger, F.; Li, C. Phenotypic plasticity of the maize root system in response to heterogeneous nitrogen availability. Planta 2014, 240, 667–678. [Google Scholar] [CrossRef]
- Olmo, M.; Villar, R.; Salazar, P.; Alburquerque, J.A. Changes in soil nutrient availability explain biochar’s impact on wheat root development. Plant Soil 2016, 399, 333–343. [Google Scholar] [CrossRef]
- Kou, L.; Guo, D.; Yang, H.; Gao, W.; Li, S. Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China. Plant Soil 2015, 391, 207–218. [Google Scholar] [CrossRef]
- Tian, Q.; Chen, F.; Zhang, F.; Mi, G. Genotypic difference in nitrogen acquisition ability in maize plants is related to the coordination of leaf and root growth. J. Plant Nutr. 2006, 29, 317–330. [Google Scholar] [CrossRef]
- Gao, K.; Chen, F.; Yuan, L.; Zhang, F.; Mi, G. A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress. Plant Cell Environ. 2015, 38, 740–750. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Bei, S.; Zhang, J.; Li, X. Growth and Distribution of Maize Roots in Response to Nitrogen Accumulation in Soil Profiles after Long-Term Fertilization Management on a Calcareous Soil. Sustainability 2018, 10, 4315. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Pig Slurry | Liquid Fraction of Digestate |
|---|---|---|
| DW (g kg−1) | 94.0 ± 0.4 | 43.0 ± 0.4 |
| OM (g kg−1) | 63.3 ± 0.6 | 25.9 ± 0.1 |
| Total C (g kg−1) | 23.6 ± 0.5 | 11.6 ± 0.2 |
| pH | 7.0 | 7.4 |
| EC (mS cm−1) | 38.9 ± 0.3 | 30.4 ± 0.5 |
| Total N (g kg−1) | 7.80 ± 0.07 | 4.77 ± 0.02 |
| NH4+-N (g kg−1) | 4.51 ± 0.07 | 2.77 ± 0.02 |
| NO3−-N (g kg−1) | <0.002 | <0.002 |
| Total P (g kg−1) | 1.75 ± 0.17 | 0.42 ± 0.03 |
| Total K (g kg−1) | 4.24 ± 0.18 | 3.90 ± 0.41 |
| Mineral N to total N ratio | 0.58 | 0.58 |
| Total C to total N ratio | 3.0 | 2.4 |
| Growing Days (Approximate Phenological Stages) | Shoot | Root | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment | FW (g pot−1) | DW (g pot−1) | N (mg g−1) | C (mg g−1) | FW (g pot−1) | DW (g pot−1) | N (mg g−1) | C (mg g−1) | |
| 20 (V2) | Control | 2.3 ± 0.8 | 0.18 ± 0.06 | 46 ± 2 b | 384 ± 2 | 2.1 ± 0.7 | 0.30 ± 0.06 | 19 ± 3 b | 276 ± 24 ab |
| CAN | 1.9 ± 0.6 | 0.15 ± 0.04 | 50 ± 11 ab | 396 ± 26 | 1.6 ± 0.3 | 0.22 ± 0.01 | 22 ± 3 ab | 287 ± 16 ab | |
| PS | 3.0 ± 0.7 | 0.23 ± 0.05 | 56 ± 4 a | 381 ± 8 | 1.8 ± 0.3 | 0.25 ± 0.04 | 25 ± 3 a | 266 ± 18 b | |
| LFD | 1.5 ± 0.6 | 0.12 ± 0.04 | 47 ± 12 b | 396 ± 16 | 1.1 ± 0.5 | 0.17 ± 0.03 | 23 ± 6 a | 300 ± 7 a | |
| 40 (V3–V4) | Control | 18 ± 6 b | 1.5 ± 0.6 | 16 ± 1 b | 409 ± 4 | 8.1 ± 2.1 | 0.90 ± 0.27 | 11 ± 1 b | 327 ± 21 |
| CAN | 26 ± 9 ab | 1.9 ± 0.8 | 36 ± 6 a | 405 ± 10 | 7.5 ± 1.9 | 0.83 ± 0.19 | 19 ± 3 a | 316 ± 17 | |
| PS | 38 ± 7 a | 2.9 ± 0.5 | 30 ± 5 a | 415 ± 5 | 9.5 ± 1.6 | 1.15 ± 0.22 | 17 ± 3 a | 317 ± 20 | |
| LFD | 29 ± 7 ab | 1.7 ± 0.3 | 33 ± 4 a | 409 ± 5 | 6.8 ± 1.0 | 0.75 ± 0.09 | 19 ± 2 a | 329 ± 14 | |
| 60 (V5–V6) | Control | 43 ± 6 c | 5 ± 1 c | 8.1 ± 0.4 | 415 ± 2 | 15 ± 1 | 1.4 ± 0.2 b | 7.6 ± 0.6 | 381 ± 8 a |
| CAN | 85 ± 20 ab | 10 ± 2 ab | 11.4 ± 0.1 | 426 ± 3 | 14 ± 2 | 2.2 ± 0.5 ab | 9.4 ± 0.6 | 335 ± 17 b | |
| PS | 100 ± 7 a | 13 ± 1 a | 8.4 ± 0.7 | 422 ± 5 | 15 ± 1 | 2.5 ± 0.3 a | 8.0 ± 0.2 | 348 ± 12 b | |
| LFD | 79 ± 10 b | 9 ± 2 b | 9.9 ± 1.3 | 420 ± 3 | 16 ± 2 | 1.9 ± 0.3 b | 8.8 ± 0.9 | 356 ± 7 ab | |
| 80 (VT) | Control | 49 ± 4 c | 8 ± 1 c | 4.9 ± 0.1 | 420 ± 3 | 9 ± 1 c | 1.3 ± 0.3 c | 6.9 ± 0.6 | 395 ± 8 |
| CAN | 147 ± 18 a | 26 ± 4 a | 4.6 ± 0.5 | 429 ± 1 | 20 ± 2 a | 3.1 ± 0.3 ab | 6.3 ± 0.3 | 372 ± 21 | |
| PS | 146 ± 4 a | 26 ± 1 a | 4.5 ± 0.4 | 430 ± 1 | 22 ± 3 a | 3.5 ± 0.7 a | 6.5 ± 0.3 | 374 ± 18 | |
| LFD | 122 ± 15 b | 19 ± 2 b | 5.1 ± 0.2 | 430 ± 1 | 16 ± 3 b | 2.5 ± 0.5 b | 6.4 ± 0.1 | 371 ± 21 | |
| 100 (R1–R3) | Control | 52 ± 4 c | 11 ± 1 c | 4.2 ± 0.1 | 403 ± 10 b | 10 ± 1 c | 1.6 ± 0.1 c | 8.6 ± 0.2 | 377 ± 11 |
| CAN | 148 ± 7 a | 32 ± 1 a | 4.0 ± 0.7 | 423 ± 3 a | 20 ± 3 a | 3.3 ± 0.3 a | 7.4 ± 0.7 | 365 ± 18 | |
| PS | 141 ± 8 a | 32 ± 2 a | 4.0 ± 0.3 | 424 ± 1 a | 21 ± 1 a | 3.4 ± 0.5 a | 7.6 ± 0.2 | 382 ± 18 | |
| LFD | 109 ± 15 b | 24 ± 3 b | 4.2 ± 0.1 | 418 ± 4 a | 14 ± 4 b | 2.2 ± 0.5 b | 8.1 ± 0.3 | 389 ± 16 | |
| Estimation Method | Biobased Fertilizers | Duration | References |
|---|---|---|---|
| Laboratory incubation | Pig slurry, digested pig slurry, and digested cattle slurry | 70 days | [34] |
| Dairy cattle slurry | 84 days | [35] | |
| Pig slurry, cattle slurry, farmyard cattle manure, and composted farmyard cattle manure | 107 days | [36] | |
| Pig manure, digestate, liquid fraction of digestate, and mineral concentrate | 120 days | [37] | |
| Pig slurry, and digested pig slurry | 56 days | [38] | |
| Pig slurry and liquid fraction of digestate | 100 days | This study | |
| Pot experiment | Unseparated digestate, and liquid fraction and solid fraction of digestate derived from animal manure or energy crops | 56 days | [39] |
| Pig slurry and liquid fraction of digestate | 100 days | This study | |
| Field experiment | Pig slurry | 3 years | [40] |
| Raw or digested liquid swine manure | 3 years | [41] | |
| Raw or digested liquid swine manure | 3 years | [42] | |
| Raw dairy manure slurry and anaerobically digested slurry | 3 years | [43] | |
| Raw liquid swine manure, solid fraction of swine manure, and digestate swine manure | 3 years | [44] | |
| Pig slurry and mineral concentrate | 2 years | [3] | |
| Raw cattle slurry, unseparated digestate, and liquid fraction and solid fraction of digestate | 3 years | [45] | |
| Digestate derived from food waste or municipal solid waste | 2 years | [46] | |
| Liquid fraction of pig manure | 2 years | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, H.; Robles-Aguilar, A.A.; Sigurnjak, I.; Michels, E.; Meers, E. Assessing Nitrogen Availability in Biobased Fertilizers: Effect of Vegetation on Mineralization Patterns. Agriculture 2021, 11, 870. https://doi.org/10.3390/agriculture11090870
Luo H, Robles-Aguilar AA, Sigurnjak I, Michels E, Meers E. Assessing Nitrogen Availability in Biobased Fertilizers: Effect of Vegetation on Mineralization Patterns. Agriculture. 2021; 11(9):870. https://doi.org/10.3390/agriculture11090870
Chicago/Turabian StyleLuo, Hongzhen, Ana A. Robles-Aguilar, Ivona Sigurnjak, Evi Michels, and Erik Meers. 2021. "Assessing Nitrogen Availability in Biobased Fertilizers: Effect of Vegetation on Mineralization Patterns" Agriculture 11, no. 9: 870. https://doi.org/10.3390/agriculture11090870