
Identifying New Resistance to Cassava Mosaic Disease and Validating Markers for the CMD2 Locus

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
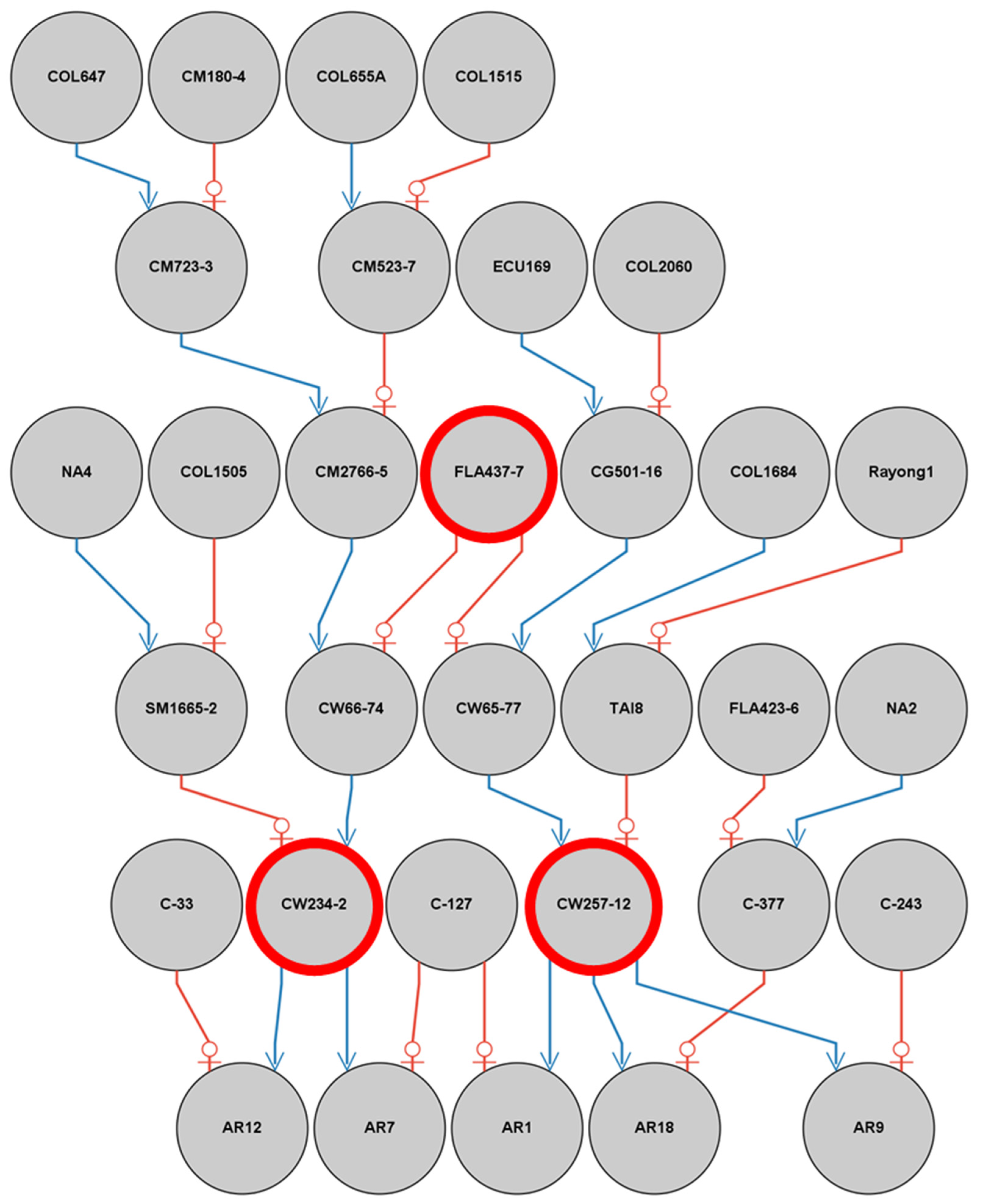
2.1. Plant Materials
2.2. Field Trials for CMD Screening
2.3. Genotyping
2.4. Data Analysis
3. Results
3.1. CMD Resistance Identified from the Populations VNM142 and CIAT102
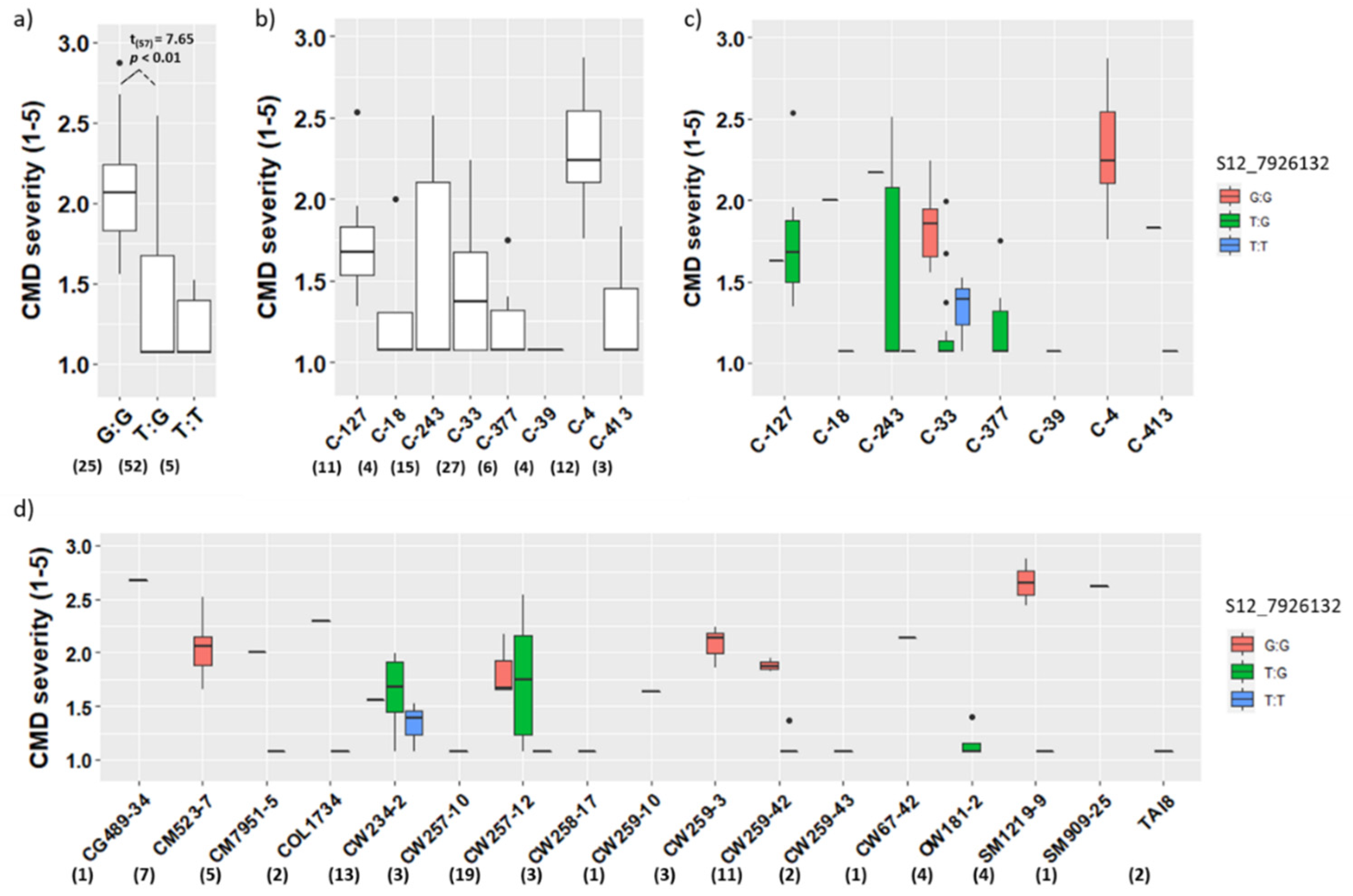
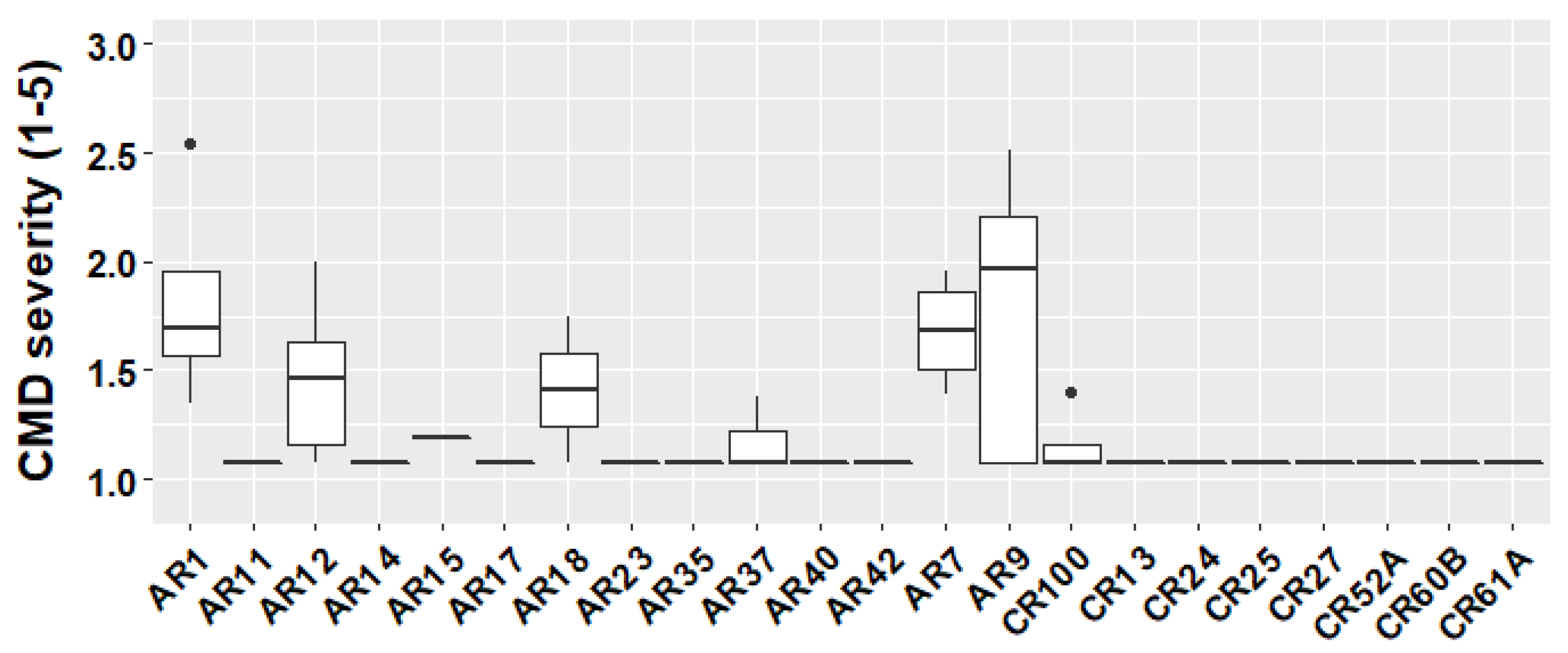
3.2. Validation of CMD Markers in the Population CIAT102
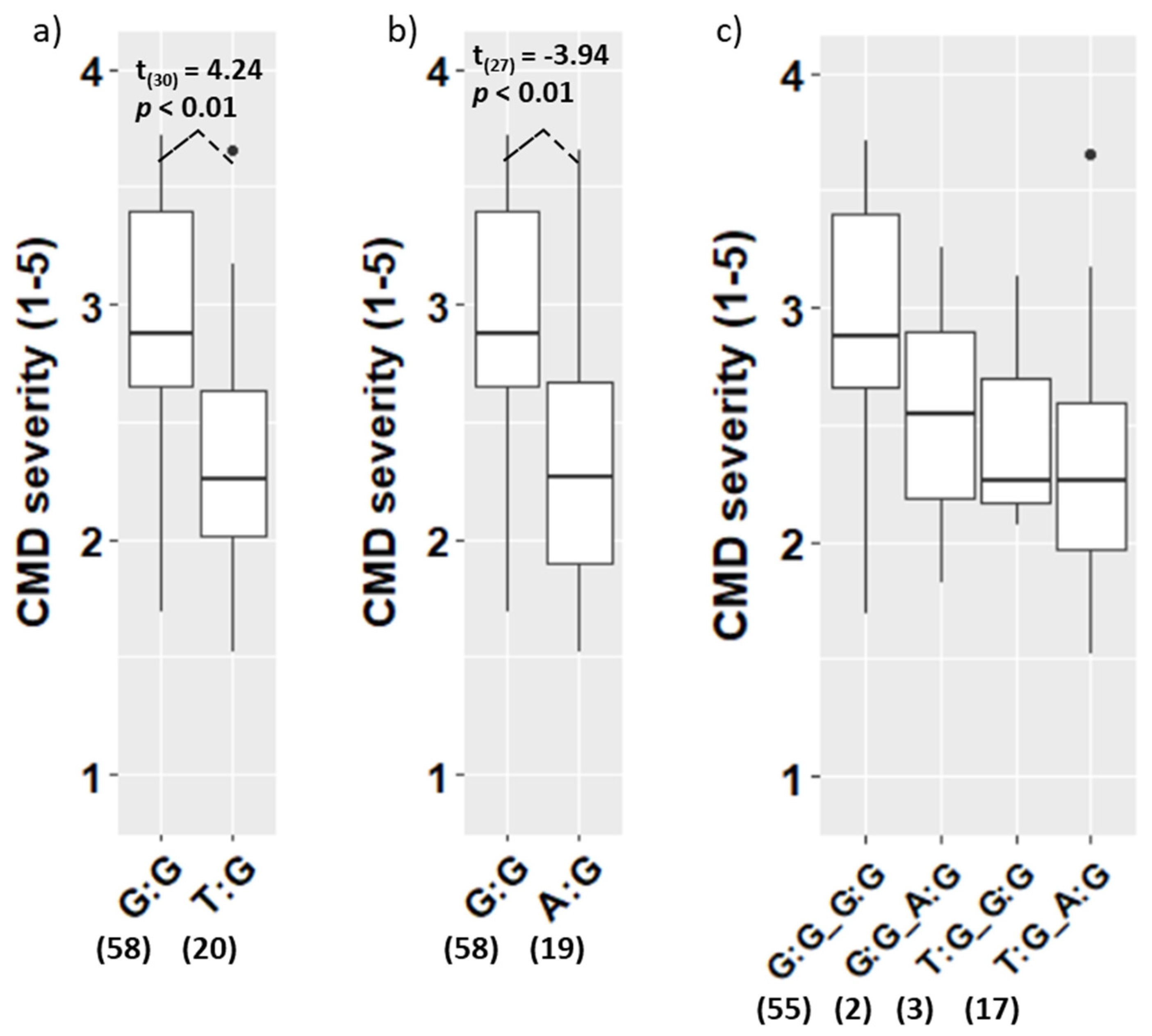
3.3. Validation of the CMD Markers in the Population VNM142
4. Discussion
4.1. CMD Resistance in the Introduced Germplasm from the CIAT and Asian Landraces
4.2. Effectiveness of the CMD Markers S12_7926132 and S14_4626854
4.3. Co-Segregation of the Markers S12_7926132 and S14_4626854
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clifton, P.; Keogh, J. Starch. In Encyclopedia of Food and Health; Elsevier: Amsterdam, The Netherlands, 2015; ISBN 9780123849533. [Google Scholar]
- FAOSTAT (Food and Agriculture Organization of the United Nations). FAOSTAT Stat Database; FAO: Rome, Italy, 2019. [Google Scholar]
- Dufour, D.; Hershey, C.; Hamaker, B.R.; Lorenzen, J. Integrating end-user preferences into breeding programmes for roots, tubers and bananas. Int. J. Food Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Malik, A.I.; Kongsil, P.; Nguyễn, V.A.; Ou, W.; Sholihin; Srean, P.; Sheela, M.N.; López-Lavalle, L.A.B.; Utsumi, Y.; Cheng, C.; et al. Cassava breeding and agronomy in Asia: 50 years of history and future directions. Breed. Sci. 2020. [Google Scholar] [CrossRef] [Green Version]
- Owor, B.; Legg, J.P.; Okao-Okuja, G.; Obonyo, R.; Ogenga-Latigo, M.W. The effect of cassava mosaic geminiviruses on symptom severity, growth and root yield of a cassava mosaic virus disease-susceptible cultivar in Uganda. Ann. Appl. Biol. 2004. [Google Scholar] [CrossRef]
- Legg, J.P.; Lava Kumar, P.; Makeshkumar, T.; Tripathi, L.; Ferguson, M.; Kanju, E.; Ntawuruhunga, P.; Cuellar, W. Cassava virus diseases: Biology, epidemiology, and management. Adv. Virus Res. 2015, 91, 85–142. [Google Scholar] [PubMed]
- Legg, J.P. Emergence, spread and strategies for controlling the pandemic of cassava mosaic virus disease in east and central Africa. Crop Prot. 1999, 18, 627–637. [Google Scholar] [CrossRef]
- Legg, J.P.; Owor, B.; Sseruwagi, P.; Ndunguru, J. Cassava Mosaic Virus Disease in East and Central Africa: Epidemiology and Management of A Regional Pandemic. Adv. Virus Res. 2006, 67, 355–418. [Google Scholar] [PubMed]
- Wang, H.L.; Cui, X.Y.; Wang, X.W.; Liu, S.S.; Zhang, Z.H.; Zhou, X.P. First report of Sri Lankan cassava mosaic virus infecting cassava in Cambodia. Plant Dis. 2016. [Google Scholar] [CrossRef]
- Chittarath, K.; Jimenez, J.; Vongphachanh, P.; Leiva, A.M.; Sengsay, S.; Lopez-Alvarez, D.; Bounvilayvong, T.; Lourido, D.; Vorlachith, V.; Cuellar, W.J. First report of Sri Lankan cassava mosaic virus and Cassava Mosaic Disease in Laos. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Wang, D.; Yao, X.M.; Huang, G.X.; Shi, T.; Wang, G.F.; Ye, J. First Report of Sri Lankan Cassava Mosaic Virus Infected Cassava in China. Plant Dis. 2019. [Google Scholar] [CrossRef]
- Uke, A.; Hoat, T.X.; Quan, M.V.; Liem, N.V.; Ugaki, M.; Natsuaki, K.T. First report of Sri Lankan Cassava Mosaic Virus infecting cassava in Vietnam. Plant Dis. 2018, 102, 2669. [Google Scholar] [CrossRef]
- Fondong, V.N. The search for resistance to cassava mosaic geminiviruses: How much we have accomplished, and what lies ahead. Front. Plant Sci. 2017, 8, 408. [Google Scholar] [CrossRef] [Green Version]
- Jennings, D.L. Breeding for resistance to African cassava mosaic geminivirus in East Africa. Trop. Sci. 1994, 34, 110–122. [Google Scholar]
- Thresh, J.M.; Cooter, R.J. Strategies for controlling cassava mosaic virus disease in Africa. Plant Pathol. 2005, 54, 587–614. [Google Scholar] [CrossRef]
- Akano, A.O.; Dixon, A.G.O.; Mba, C.; Barrera, E.; Fregene, M. Genetic mapping of a dominant gene conferring resistance to cassava mosaic disease. Theor. Appl. Genet. 2002. [Google Scholar] [CrossRef] [PubMed]
- Rabbi, I.Y.; Hamblin, M.T.; Kumar, P.L.; Gedil, M.A.; Ikpan, A.S.; Jannink, J.L.; Kulakow, P.A. High-resolution mapping of resistance to cassava mosaic geminiviruses in cassava using genotyping-by-sequencing and its implications for breeding. Virus Res. 2014. [Google Scholar] [CrossRef] [Green Version]
- Rabbi, I.Y.; Kayondo, S.I.; Bauchet, G.; Yusuf, M.; Aghogho, C.I.; Ogunpaimo, K.; Uwugiaren, R.; Smith, I.A.; Peteti, P.; Agbona, A.; et al. Genome-wide association analysis reveals new insights into the genetic architecture of defensive, agro-morphological and quality-related traits in cassava. Plant Mol. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, M.D.; Rabbi, I.Y.; Egesi, C.; Hamblin, M.; Kawuki, R.; Kulakow, P.; Lozano, R.; Del Carpio, D.P.; Ramu, P.; Jannink, J.-L. Genome-Wide Association and Prediction Reveals Genetic Architecture of Cassava Mosaic Disease Resistance and Prospects for Rapid Genetic Improvement. Plant Genome 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceballos, H.; Rojanaridpiched, C.; Chalermpol, P.; Becerra, L.A.; Kittipadakul, P.; Iglesias, C.; Gracen, V. Excellence in Cassava Breeding: Perspectives for the Future. Crop Breeding, Genet. Genomics 2020. [Google Scholar] [CrossRef] [Green Version]
- Kuon, J.E.; Qi, W.; Schläpfer, P.; Hirsch-Hoffmann, M.; Von Bieberstein, P.R.; Patrignani, A.; Poveda, L.; Grob, S.; Keller, M.; Shimizu-Inatsugi, R.; et al. Haplotype-resolved genomes of geminivirus-resistant and geminivirus-susceptible African cassava cultivars. BMC Biol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Ige, A.D.; Olasanmi, B.; Nkouaya Mbanjo, E.G.; Kayondo, I.S.; Parkes, E.Y.; Kulakow, P.; Egesi, C.; Bauchet, G.J.; Ng, E.; Becerra Lopez-Lavalle, L.A.; et al. Conversion and Validation of Uniplex SNP Markers for Selection of Resistance to Cassava Mosaic Disease in Cassava Breeding Programs. Agronomy 2021, 11, 420. [Google Scholar] [CrossRef]
- Fernandez-Pozo, N.; Menda, N.; Edwards, J.D.; Saha, S.; Tecle, I.Y.; Strickler, S.R.; Bombarely, A.; Fisher-York, T.; Pujar, A.; Foerster, H.; et al. The Sol Genomics Network (SGN)-from genotype to phenotype to breeding. Nucleic Acids Res. 2015. [Google Scholar] [CrossRef]
- Le, D.P.; Labarta, R.A.; de Haan, S.; Maredia, M.; Becerra, L.A.; Nhu, L.T.; Ovalle, T.; Nguyen, V.A.; Pham, N.T.; Nguyen, H.H.; et al. Characterization of Cassava Production Systems in Vietnam; International Center for Tropical Agriculture (CIAT): Hanoi, Vietnam, 2019; 54p. [Google Scholar]
- Ocampo, J.; Ovalle, T.; Labarta, R.; Le, D.P.; de Haan, S.; Vu, N.A.; Kha, L.Q.; Becerra Lopez-Lavalle, L.A. DNA fingerprinting reveals varietal composition of Vietnamese cassava germplasm (Manihot esculenta Crantz) from farmers’ field and genebank collections. Plant Mol. Biol. 2021. [Google Scholar] [CrossRef]
- Peña-Venegas, C.P.; Stomph, T.J.; Verschoor, G.; Lopez-Lavalle, L.A.B.; Struik, P.C. Differences in manioc diversity among five ethnic groups of the Colombian Amazon. Diversity 2014, 4, 792. [Google Scholar] [CrossRef] [Green Version]
- Floro, V.O.; Labarta, R.A.; Becerra López-Lavalle, L.A.; Martinez, J.M.; Ovalle, T.M. Household Determinants of the Adoption of Improved Cassava Varieties using DNA Fingerprinting to Identify Varieties in Farmer Fields: A Case Study in Colombia. J. Agric. Econ. 2018. [Google Scholar] [CrossRef] [Green Version]
- Hahn, S.K.; Terry, E.R.; Leuschner, K. Breeding cassava for resistance to cassava mosaic disease. Euphytica 1980. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Charoenkul, N.; Uttapap, D.; Pathipanawat, W.; Takeda, Y. Molecular structure of starches from cassava varieties having different cooked root textures. Starch/Staerke 2006. [Google Scholar] [CrossRef]
- Whankaew, S.; Poopear, S.; Kanjanawattanawong, S.; Tangphatsornruang, S.; Boonseng, O.; Lightfoot, D.A.; Triwitayakorn, K. A genome scan for quantitative trait loci affecting cyanogenic potential of cassava root in an outbred population. BMC Genomics 2011. [Google Scholar] [CrossRef] [Green Version]
- Chaengsee, P.; Kongsil, P.; Siriwong, N.; Kittipadakul, P.; Piyachomkwan, K.; Petchpoung, K. Potential yield and cyanogenic glucoside content of cassava root and pasting properties of starch and flour from cassava Hanatee var. And breeding lines grown under rain-fed condition. Agric. Nat. Resour. 2020. [Google Scholar] [CrossRef]
- Chiewchankaset, P.; Siriwat, W.; Suksangpanomrung, M.; Boonseng, O.; Meechai, A.; Tanticharoen, M.; Kalapanulak, S.; Saithong, T. Understanding carbon utilization routes between high and low starch-producing cultivars of cassava through Flux Balance Analysis. Sci. Rep. 2019, 9, 2964. [Google Scholar] [CrossRef]
- Ferguson, M.E.; Shah, T.; Kulakow, P.; Ceballos, H. A global overview of cassava genetic diversity. PLoS ONE 2019, 14, e0224763. [Google Scholar] [CrossRef]
- Sheat, S.; Fuerholzner, B.; Stein, B.; Winter, S. Resistance against cassava brown streak viruses from africa in cassava germplasm from South America. Front. Plant Sci. 2019. [Google Scholar] [CrossRef]
- Mansfeld, B.N.; Boyher, A.; Berry, J.C.; Wilson, M.; Ou, S.; Polydore, S.; Michael, T.P.; Fahlgren, N.; Bart, R.S. Large structural variations in the haplotype-resolved African cassava genome. bioRxiv 2021. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Trial | Trait | Mean | Median | Range | Vg | Ve | H2 |
|---|---|---|---|---|---|---|---|---|
| VNM142 | 201801MDEAR | CMD_1.5MAP | 1.56 | 1.43 | 1.00–4.00 | 0.10 | 0.21 | 0.50 |
| CMD_3MAP | 1.87 | 1.75 | 1.00–4.00 | 0.18 | 0.22 | 0.63 | ||
| CMD_6MAP | 2.23 | 2.20 | 1.00–4.00 | 0.27 | 0.19 | 0.75 | ||
| CMD_10MAP | 2.82 | 2.86 | 1.19–4.00 | 0.40 | 0.19 | 0.82 | ||
| 201901MDEAR * | CMD_10MAP | 2.65 | 2.33 | 1.00–4.12 | 1.30 | 0.04 | 0.99 | |
| 201902MDEAR # | CMD_10MAP | 2.83 | 2.63 | 1.70–4.10 | 0.62 | 0.11 | 0.94 | |
| CIAT102 | 201903MDEAR | CMD_3MAP | 1.73 | 1.64 | 1.00–3.77 | 0.42 | 0.12 | 0.91 |
| Genotype/Group | Clone | 2018–2019 | 2019–2020 | S12_7926132 | S14_4626854 |
|---|---|---|---|---|---|
| UNK-CI-2 | VN19-442 | 1.5 | 2.3 | T:G | A:G |
| CR63_PER262_TAI9 | VN19-1432, VN19-1556 | 1.6 | 1.9 | T:G | A:G |
| KM57_VNM8_Xanh Vinh Phu | VN19-1039, VN19-1050 | 1.6 | 1.9 | T:G | A:G |
| UNQ-115 | VN19-773 | 1.7 | 2.1 | T:G | A:G |
| UNK-F | VN19-1184, VN19-1194 | 2.0 | 2.6 | T:G | A:G |
| UNQ-44 | VN19-320 | 1.7 | 1.8 | G:G | G:G |
| UNK-AF-2 | VN19-1805 | 1.8 | NA | G:G | A:G |
| UNK-CH | VN19-390 | 1.9 | 2.2 | G:G | G:G |
| KU50_KM94_TAI16 | 11 clone samples (e.g., VN19-1739) | 2.6 | 3.5 | G:G | G:G |
| KM140 | 4 clone samples (e.g., VN19-2659) | 3.6 | NA | G:G | G:G |
| KM419 | 2 clone samples (e.g., VN19-2202) | 3.0 | 4.0 | G:G | G:G |
| C33 | C33 | NA | 1.1 | T:G | A:G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thuy, C.T.L.; Lopez-Lavalle, L.A.B.; Vu, N.A.; Hy, N.H.; Nhan, P.T.; Ceballos, H.; Newby, J.; Tung, N.B.; Hien, N.T.; Tuan, L.N.; et al. Identifying New Resistance to Cassava Mosaic Disease and Validating Markers for the CMD2 Locus. Agriculture 2021, 11, 829. https://doi.org/10.3390/agriculture11090829
Thuy CTL, Lopez-Lavalle LAB, Vu NA, Hy NH, Nhan PT, Ceballos H, Newby J, Tung NB, Hien NT, Tuan LN, et al. Identifying New Resistance to Cassava Mosaic Disease and Validating Markers for the CMD2 Locus. Agriculture. 2021; 11(9):829. https://doi.org/10.3390/agriculture11090829
Chicago/Turabian StyleThuy, Cu Thi Le, Luis Augusto Becerra Lopez-Lavalle, Nguyen Anh Vu, Nguyen Huu Hy, Pham Thi Nhan, Hernan Ceballos, Jonathan Newby, Nguyen Ba Tung, Nguyen Trong Hien, Le Ngoc Tuan, and et al. 2021. "Identifying New Resistance to Cassava Mosaic Disease and Validating Markers for the CMD2 Locus" Agriculture 11, no. 9: 829. https://doi.org/10.3390/agriculture11090829