
Microbial Interactions and Roles in Soil Fertility in Seasonal Freeze-Thaw Periods under Different Straw Returning Strategies

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
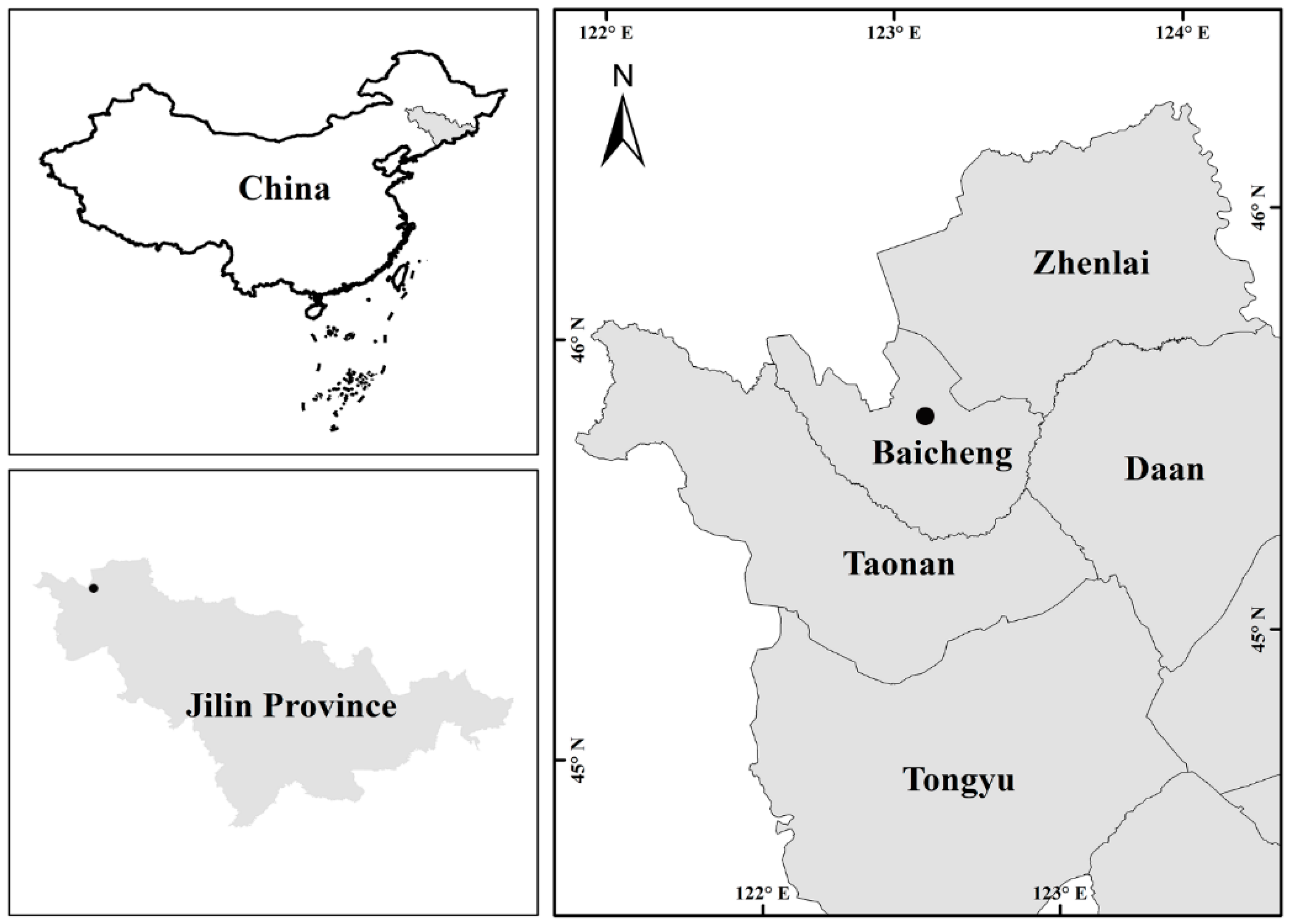
2.1. Site Description
2.2. Experimental Design and Sample Collection
2.3. Environmental Parameters
2.4. DNA Extraction and Sequencing
2.5. Bacterial Community Analysis
2.6. Correlation-Based Co-Occurrence Network and Topological Features
2.7. Variation Patterns of Treatment Modules
2.8. Statistical Analysis
3. Results
3.1. Chemical Properties of Soil
3.2. Bacterial Community Composition
3.3. Multidimensional Analysis and the Potential Drivers of Bacterial Community Composition
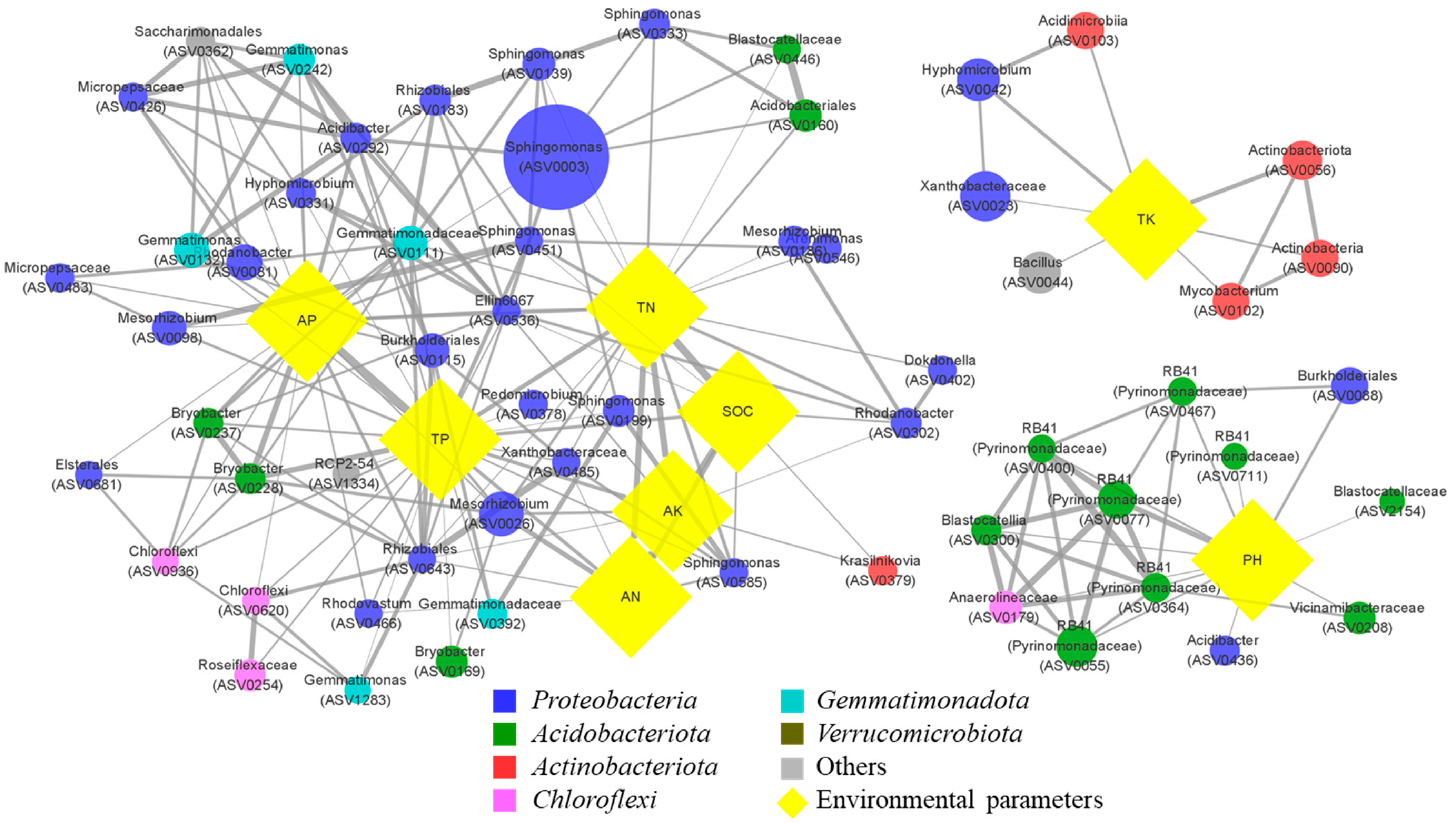
3.4. Co-Occurrence Networks, the Modular Structures, and the Taxonomic Features
3.5. Distinctive Bacterial Associations with Environmental Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Paul, E.A.; Clark, F.E. Soil Microbiology and Biochemistry; Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Van Gestel, M.; Ladd, J.; Amato, M. Microbial biomass responses to seasonal change and imposed drying regimes at increasing depths of undisturbed topsoil profiles. Soil Biol. Biochem. 1992, 24, 103–111. [Google Scholar] [CrossRef]
- Corre, M.D.; Schnabel, R.R.; Stout, W.L. Spatial and seasonal variation of gross nitrogen transformations and microbial biomass in a Northeastern US grassland. Soil Biol. Biochem. 2002, 34, 445–457. [Google Scholar] [CrossRef]
- Bell, C.; McIntyre, N.; Cox, S.; Tissue, D.; Zak, J. Soil microbial responses to temporal variations of moisture and temperature in a Chihuahuan Desert grassland. Microb. Ecol. 2008, 56, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, T.; Gilliam, F.S.; Gundersen, P.; Zhang, W.; Chen, H.; Mo, J. Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest. PLoS ONE 2013, 8, e61188. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Zhang, X.; Liu, X.; Fu, X.; Chen, F.; Wang, H.; Sun, X.; Wen, X. Responses of soil microbial communities and enzyme activities to nitrogen and phosphorus additions in Chinese fir plantations of subtropical China. Biogeosciences 2015, 12, 5537–5546. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.A.; Chen, H.Y.; Ruan, H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P.; Umar, S.; Baumann, K. The microbial community composition changes rapidly in the early stages of decomposition of wheat residue. Soil Biol. Biochem. 2011, 43, 445–451. [Google Scholar] [CrossRef]
- Cong, P.; Wang, J.; Li, Y.; Liu, N.; Dong, J.; Pang, H.; Zhang, L.; Gao, Z. Changes in soil organic carbon and microbial community under varying straw incorporation strategies. Soil Tillage Res. 2020, 204, 104735. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Li, X.; Su, Z.; Li, X.; Xu, M. Effects of tillage and residue incorporation on composition and abundance of microbial communities of a fluvo-aquic soil. Eur. J. Soil Biol. 2014, 65, 70–78. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Röttjers, L.; Faust, K. From hairballs to hypotheses–biological insights from microbial networks. FEMS Microbiol. Rev. 2018, 42, 761–780. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.-K.; Wei, L.; Turner, N.C.; Ma, S.-C.; Yang, M.-D.; Wang, T.-C. Improved straw management practices promote in situ straw decomposition and nutrient release, and increase crop production. J. Clean. Prod. 2020, 250, 119514. [Google Scholar] [CrossRef]
- Zhao, S.; Qiu, S.; Xu, X.; Ciampitti, I.A.; Zhang, S.; He, P. Change in straw decomposition rate and soil microbial community composition after straw addition in different long-term fertilization soils. Appl. Soil Ecol. 2019, 138, 123–133. [Google Scholar] [CrossRef]
- Zhang, H.; Hobbie, E.A.; Feng, P.; Zhou, Z.; Niu, L.A.; Duan, W.; Hao, J.; Hu, K. Responses of soil organic carbon and crop yields to 33-year mineral fertilizer and straw additions under different tillage systems. Soil Tillage Res. 2021, 209, 104943. [Google Scholar] [CrossRef]
- Marcó, A.; Rubio, R.; Compañó, R.; Casals, I. Comparison of the Kjeldahl method and a combustion method for total nitrogen determination in animal feed. Talanta 2002, 57, 1019–1026. [Google Scholar] [CrossRef]
- Joshi, S.R.; Morris, J.W.; Tfaily, M.M.; Young, R.P.; McNear, D.H. Low soil phosphorus availability triggers maize growth stage specific rhizosphere processes leading to mineralization of organic P. Plant Soil 2021, 459, 423–440. [Google Scholar] [CrossRef]
- Guo, A.; Zhao, Z.; Zhang, P.; Yang, Q.; Li, Y.; Wang, G. Linkage between soil nutrient and microbial characteristic in an opencast mine, China. Sci. Total Environ. 2019, 671, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbiomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Inter. J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Assenov, Y.; Ramirez, F.; Schelhorn, S.E.; Lengauer, T.; Albrecht, M. Computing topological parameters of biological networks. Bioinformatics 2008, 24, 282–284. [Google Scholar] [CrossRef] [Green Version]
- Telesford, Q.K.; Joyce, K.E.; Hayasaka, S.; Burdette, J.H.; Laurienti, P.J. The ubiquity of small-world networks. Brain Connect. 2011, 1, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Chun, S.-J.; Cui, Y.; Lee, C.S.; Cho, A.R.; Baek, K.; Choi, A.; Ko, S.-R.; Lee, H.-G.; Hwang, S.; Oh, H.-M. Characterization of distinct cyanoHABs-related modules in microbial recurrent association network. Front. Microbiol. 2019, 10, 1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H.H. Vegan: Community Ecology Package R Package Version 2.0-8. 2013. Available online: http://cran.r-project.org/package=vegan (accessed on 14 April 2021).
- Whitman, W.B.; Oren, A.; Chuvochina, M.; da Costa, M.S. Proposal of the suffix-ota to denote phyla. Addendum to ‘Proposal to include the rank of phylum in the International Code of Nomenclature of Prokaryotes’. Int. J. Syst. Evol. Microbiol. 2018, 68, 967–969. [Google Scholar] [CrossRef]
- Schmidt, S.; Costello, E.; Nemergut, D.; Cleveland, C.C.; Reed, S.; Weintraub, M.; Meyer, A.; Martin, A. Biogeochemical consequences of rapid microbial turnover and seasonal succession in soil. Ecology 2007, 88, 1379–1385. [Google Scholar] [CrossRef]
- Koranda, M.; Kaiser, C.; Fuchslueger, L.; Kitzler, B.; Sessitsch, A.; Zechmeister-Boltenstern, S.; Richter, A. Seasonal variation in functional properties of microbial communities in beech forest soil. Soil Biol. Biochem. 2013, 60, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Will, C.; Thürmer, A.; Wollherr, A.; Nacke, H.; Herold, N.; Schrumpf, M.; Gutknecht, J.; Wubet, T.; Buscot, F.; Daniel, R. Horizon-specific bacterial community composition of German grassland soils, as revealed by pyrosequencing-based analysis of 16S rRNA genes. Appl. Environ. Microbiol. 2010, 76, 6751–6759. [Google Scholar] [CrossRef] [Green Version]
- Shu, W.; Pablo, G.P.; Jun, Y.; Danfeng, H. Abundance and diversity of nitrogen-fixing bacteria in rhizosphere and bulk paddy soil under different duration of organic management. World J. Microbiol. Biotechnol. 2012, 28, 493–503. [Google Scholar] [CrossRef]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Kang, S.-M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.-Y. Bacterial endophyte Sphingomonas sp. LK11 produces gibberellins and IAA and promotes tomato plant growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, X.; Cao, Z.; Zhao, K.; Wang, S.; Chen, M.; Hu, X. Growth-promoting Sphingomonas paucimobilis ZJSH 1 associated with Dendrobium officinale through phytohormone production and nitrogen fixation. Microb. Biotechnol. 2014, 7, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, F.; Huang, Y.; Zhou, M.; Gao, J.; Yan, T.; Sheng, H.; An, L. Sphingomonas sp. Cra20 increases plant growth rate and alters rhizosphere microbial community structure of Arabidopsis thaliana under drought stress. Front. Microbiol. 2019, 10, 1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Meng, D.; Li, J.; Yin, H.; Liu, H.; Liu, X.; Cheng, C.; Xiao, Y.; Liu, Z.; Yan, M. Response of soil microbial communities and microbial interactions to long-term heavy metal contamination. Environ. Pollut. 2017, 231, 908–917. [Google Scholar] [CrossRef]
- Sun, W.; Xiao, E.; Pu, Z.; Krumins, V.; Dong, Y.; Li, B.; Hu, M. Paddy soil microbial communities driven by environment-and microbe-microbe interactions: A case study of elevation-resolved microbial communities in a rice terrace. Sci. Total Environ. 2018, 612, 884–893. [Google Scholar] [CrossRef]
- Xue, M.; Guo, Z.; Gu, X.; Gao, H.; Weng, S.; Zhou, J.; Gu, D.; Lu, H.; Gu, D.; Lu, H.; et al. Rare rather than abundant microbial communities drive the effects of long-term greenhouse cultivation on ecosystem functions in subtropical agricultural soils. Sci. Total Environ. 2020, 706, 136004. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Singh, R.; Eltis, L.D.; Mohn, W.W. Bacterial contributions to delignification and lignocellulose degradation in forest soils with metagenomic and quantitative stable isotope probing. ISME J. 2019, 13, 413–429. [Google Scholar] [CrossRef] [Green Version]
- Žifčáková, L.; Větrovský, T.; Lombard, V.; Hebrussat, B.; Howe, A.; Baldrian, P. Feed in summer, rest in winter: Microbial carbon utilization in forest topsoil. Microbiome 2017, 5, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.C.; Morgan, X.C.; Power, J.F.; Dunfield, P.F.; Huttenhower, C.; Stott, M.B. Complete genome sequence of the thermophilic Acidobacteria, Pyrinomonas methylaliphatogenes type strain K22 T. Stand. Genom. Sci. 2015, 10, 1–8. [Google Scholar]
- Wüst, P.K.; Foesel, B.U.; Geppert, A.; Huber, K.J.; Luckner, M.; Wanner, G.; Overmann, J. Brevitalea aridisoli, B. deliciosa and Arenimicrobium luteum, three novel species of Acidobacteria subdivision 4 (class Blastocatellia) isolated from savanna soil and description of the novel family Pyrinomonadaceae. Int. J. Syst. Evol. Microbiol. 2016, 66, 3355–3366. [Google Scholar] [CrossRef]
- Cui, Y.; Jin, L.; Ko, S.-R.; Chun, S.-J.; Oh, H.-S.; Lee, C.S.; Srivastava, A.; Oh, H.-M.; Ahn, C.-Y. Periphyton effects on bacterial assemblages and harmful cyanobacterial blooms in a eutrophic freshwater lake: A mesocosm study. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Lu, Q.; Wei, Y.; Cui, H.; Zhang, X.; Wang, X.; Shan, S.; Wei, Z. Effect of actinobacteria agent inoculation methods on cellulose degradation during composting based on redundancy analysis. Bioresour. Technol. 2016, 219, 196–203. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Month | Sample ID | AN | AP | AK | TN | TP | TK | pH | SOC | |
|---|---|---|---|---|---|---|---|---|---|---|
| (mg kg−1) | (mg kg−1) | (mg kg−1) | (g kg−1) | (g kg−1) | (g kg−1) | (g kg−1) | ||||
| CK | August | CK201808 | 136.72 ± 0.33 a | 35.99 ± 1.79 b | 134.00 ± 9.89 ab | 1.46 ± 0.08 a | 0.48 ± 0.09 ab | 22.75 ± 1.69 a | 6.59 ± 0.15 ab | 13.57 ± 0.16 a |
| December | CK201812 | 88.12 ± 0.14 c | 11.34 ± 0.67 d | 85.00 ± 8.48 c | 1.04 ± 0.09 b | 0.31 ± 0.08 b | 24.37 ± 0.98 a | 6.72 ± 0.04 a | 10.79 ± 0.98 b | |
| March | CK201903 | 88.12 ± 0.28 c | 10.70 ± 0.59 d | 91.00 ± 12.7 c | 1.04 ± 0.05 b | 0.31 ± 0.02 b | 24.79 ± 1.27 a | 6.40 ± 0.04 bc | 11.6 ± 0.82 b | |
| April | CK201904 | 121.07 ± 2.00 b | 23.00 ± 1.47 c | 115.00 ± 11.3 bc | 1.10 ± 0.07 b | 0.39 ± 0.07 ab | 22.57 ± 0.86 a | 6.48 ± 0.02 b | 11.31 ± 0.45 b | |
| May | CK201905 | 118.96 ± 1.01 b | 54.06 ± 1.90 a | 156.00 ± 16.9 a | 1.37 ± 0.09 a | 0.55 ± 0.08 a | 23.56 ± 1.00 a | 6.04 ± 0.05 c | 13.63 ± 0.87 a | |
| CS | August | CS201808 | 136.72 ± 2.12 a | 28.34 ± 0.84 b | 150.00 ± 12.7 a | 1.53 ± 0.07 a | 0.46 ± 0.05 abc | 22.55 ± 0.86 a | 6.63 ± 0.08 ab | 15.89 ± 1.01 a |
| December | CS201812 | 117.49 ± 1.96 c | 55.12 ± 1.13 a | 117.00 ± 15.5 b | 1.19 ± 0.11 bc | 0.54 ± 0.05 ab | 23.77 ± 1.24 a | 6.51 ± 0.09 bc | 12.65 ± 0.93 b | |
| March | CS201903 | 110.15 ± 1.01 d | 56.18 ± 0.59 a | 118.00 ± 11.3 b | 1.23 ± 0.05 b | 0.57 ± 0.07 a | 25.98 ± 1.76 a | 6.12 ± 0.07d | 12.59 ± 0.80 b | |
| April | CS201904 | 104.92 ± 1.32 a | 6.50 ± 0.26 d | 109.00 ± 8.48 b | 1.02 ± 0.05 c | 0.32 ± 0.04 c | 24.07 ± 1.52 a | 6.89 ± 0.15 a | 10.85 ± 0.48 b | |
| May | CS201905 | 108.68 ± 0.83 b | 21.33 ± 0.72 c | 98.00 ± 9.89 b | 1.23 ± 0.04 b | 0.41 ± 0.05 bc | 22.58 ± 0.86 a | 6.33 ± 0.08 cd | 15.89 ± 0.98 a | |
| RS | August | RS201808 | 120.37 ± 2.34 a | 40.24 ± 0.80 a | 116.00 ± 8.48 a | 1.30 ± 0.11 a | 0.49 ± 0.09 a | 22.57 ± 0.98 b | 6.68 ± 0.08 a | 12.35 ± 0.55 a |
| December | RS201812 | 96.93 ± 0.84 c | 10.49 ± 0.29 d | 89.00 ± 15.5 a | 1.03 ± 0.05 b | 0.29 ± 0.02 b | 23.96 ± 1.51 ab | 6.29 ± 0.12 b | 10.79 ± 0.82 ab | |
| March | RS201903 | 89.95 ± 0.25 d | 9.85 ± 0.25 d | 85.00 ± 9.89 a | 1.03 ± 0.07 b | 0.32 ± 0.04 ab | 25.58 ± 1.04 a | 6.44 ± 0.07 b | 10.09 ± 0.42 b | |
| April | RS201904 | 118.24 ± 1.01 a | 13.3 ± 0.59 c | 116.00 ± 14.1 a | 1.07 ± 0.09 ab | 0.35 ± 0.08 ab | 23.56 ± 0.77 ab | 6.28 ± 0.09 b | 10.9 ± 0.90 ab | |
| May | RS201905 | 108.68 ± 0.91 b | 16.23 ± 0.45 b | 115.00 ± 12.7 a | 1.15 ± 0.08 ab | 0.38 ± 0.08 ab | 24.35 ± 0.65 ab | 6.31 ± 0.05 b | 12.01 ± 0.45 a | |
| CRS | August | CRS201808 | 103.67 ± 0.96 a | 13.89 ± 0.29 b | 100.00 ± 9.89 ab | 1.20 ± 0.04 a | 0.38 ± 0.05 a | 22.16 ± 0.25 b | 6.71 ± 0.08 a | 11.02 ± 0.76 a |
| December | CRS201812 | 88.12 ± 0.22 b | 16.44 ± 0.60 a | 77.00 ± 5.65 c | 0.98 ± 0.05 b | 0.37 ± 0.04 a | 24.38 ± 0.65 a | 6.81 ± 0.04 a | 9.45 ± 0.69 a | |
| March | CRS201903 | 88.12 ± 0.62 b | 14.74 ± 0.67 b | 82.00 ± 7.07 bc | 0.98 ± 0.08 b | 0.37 ± 0.05 a | 22.76 ± 0.82 ab | 6.45 ± 0.04 b | 10.03 ± 0.79 a | |
| April | CRS201904 | 104.92 ± 1.01 a | 11.10 ± 0.43 c | 108.00 ± 9.89 a | 1.05 ± 0.08 ab | 0.33 ± 0.08 a | 23.29 ± 0.79 ab | 6.725 ± 0.12 a | 10.67 ± 0.67 a | |
| May | CRS201905 | 88.12 ± 0.26 b | 15.17 ± 0.56 ab | 75.00 ± 9.89 c | 1.00 ± 0.07 b | 0.35 ± 0.01 a | 23.14 ± 0.63 ab | 6.38 ± 0.14 b | 9.74 ± 0.72 a |
| Factor | Index | Df | F Value | p Value |
|---|---|---|---|---|
| Treatments * | AN | 3 | 577.394 | 0.000 |
| AK | 3 | 14.570 | 0.000 | |
| AP | 3 | 971.650 | 0.000 | |
| TN | 3 | 12.763 | 0.000 | |
| TP | 3 | 4.813 | 0.011 | |
| TK | 3 | 1.144 | 0.356 | |
| pH | 3 | 9.968 | 0.000 | |
| SOC | 3 | 36.868 | 0.000 | |
| Months | AN | 4 | 840.829 | 0.000 |
| AK | 4 | 11.674 | 0.000 | |
| AP | 4 | 380.110 | 0.000 | |
| TN | 4 | 23.829 | 0.000 | |
| TP | 4 | 2.959 | 0.045 | |
| TK | 4 | 5.031 | 0.006 | |
| pH | 4 | 26.827 | 0.000 | |
| SOC | 4 | 17.966 | 0.000 | |
| Treatment × Months | AN | 12 | 142.801 | 0.000 |
| AK | 12 | 4.929 | 0.001 | |
| AP | 12 | 704.079 | 0.000 | |
| TN | 12 | 2.302 | 0.048 | |
| TP | 12 | 3.173 | 0.011 | |
| TK | 12 | 1.067 | 0.433 | |
| pH | 12 | 7.786 | 0.000 | |
| SOC | 12 | 3.526 | 0.006 | |
| AN | 19 | 358.375 | 0.000 | |
| Calibration model | AK | 19 | 7.871 | 0.000 |
| AP | 19 | 678.123 | 0.000 | |
| TN | 19 | 8.486 | 0.000 | |
| TP | 19 | 3.387 | 0.005 | |
| TK | 19 | 1.914 | 0.079 | |
| pH | 19 | 12.139 | 0.000 | |
| SOC | 19 | 11.831 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.; Chen, B.; Wang, H.; Wang, N.; Ma, T.; Cui, Y.; Luan, T.; Chun, S.; Liu, C.; Wang, L. Microbial Interactions and Roles in Soil Fertility in Seasonal Freeze-Thaw Periods under Different Straw Returning Strategies. Agriculture 2021, 11, 779. https://doi.org/10.3390/agriculture11080779
Sun M, Chen B, Wang H, Wang N, Ma T, Cui Y, Luan T, Chun S, Liu C, Wang L. Microbial Interactions and Roles in Soil Fertility in Seasonal Freeze-Thaw Periods under Different Straw Returning Strategies. Agriculture. 2021; 11(8):779. https://doi.org/10.3390/agriculture11080779
Chicago/Turabian StyleSun, Mengqi, Baoyu Chen, Hongjun Wang, Nan Wang, Taigang Ma, Yingshun Cui, Tianhao Luan, Seongjun Chun, Chunguang Liu, and Lichun Wang. 2021. "Microbial Interactions and Roles in Soil Fertility in Seasonal Freeze-Thaw Periods under Different Straw Returning Strategies" Agriculture 11, no. 8: 779. https://doi.org/10.3390/agriculture11080779